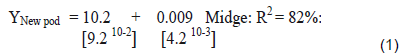ABSTRACT
Cocoa midges [Forcipomyia sp (Diptera: Cerato-pogonidae)] are major pollinators of cocoa and it is assumed that the number of fertilized pods and the increase in bean numbers may be the approach to enhancing cocoa yield. An insect survey using suction traps was employed to estimate the midge population dynamics in three Caribbean territories. Separate studies were conducted on the cocoa floral and reproductive phenology in addition to the evaluation of several naturally occurring substrates. The results indicated that the insect population as determined by the suction traps were low (27.1 ± 3.37 to 53.5 ± 8.47 transect site). The trees maintained the floral prolificacy even though the pollination [%] was low for Jamaica (0.91), Trinidad (0.88), and Tobago (0.11). However, it was improved when the midge pollinator population was increased with augmentation of substrates of cacao pods [5660] and banana pseudo-stem (1885). This resulted in significant increases in new pods which increased from < 10 pods/tree in the untreated areas to 49 to 76 pods/tree with substrate augmentation. It was evident that the discarded cocoa pod after harvest was a suitable feeding substrate and breeding site for the midge. This information is to be used to advance further studies in plant-pheromones which can serve as attractants to increase pollination/fertilization in cocoa.
Key words: Theobroma cacao, cocoa midges, substrate augmentation, pollinators, floral phenology.
The cacao industry is driven by the major international chocolate manufacturing in Europe and USA. However, all the raw materials are produced in the tropical south and Central America, Africa and the Caribbean (Motamayor et al., 2002). Commercial cacao (Theobroma cacao L.; formerly Sterculiaceae family; reclassified Malvaceae family] (Alverson et al., 1999) is a tropical tree [3 to 5 m] which is derived from varieties belonging to three major groups viz: Criollo, Forastero and Trinitario (Lachenaud et al., 1997).
The varieties and the hybrids exhibit considerable genetic variability in morphological and physiological traits (Cheesman, 1944; Bartley, 2005; Daymond and Hadley, 2004; Maharaj et al., 2011).
The crop growth is highly influenced by environmental conditions viz. temperature (Daymond and Hadley, 2004), flooding (Sena and Kozlowski, 1986), and water stress (Almeida and Valle, 2007). The bi-modal seasons influence the phenological stages of flowering, fruiting and pod growth (Cazorla et al., 1989). The plant produces caulescent flowers with the non-pollinated flowers abscising 24 to 36 h after anthesis (Garcia, 1973). The cacao flower is hermaphrodite and is pollinated by insects, mainly Forcipomyia sp. (Diptera: Ceratopogonidae (Dias et al., 1997)). The flowers setting to pods are very low [0.5 to 5%] (Aneja et al., 1999).
The quality of pollination can depend on two factors, the degree of pollen compatibility and the number of pollen grains deposited on the stigma (Lanaud et al., 1987). It is assumed that with increased pollen grains pod set is improved (Hasenstein and Zavada, 2001) and more pollinations result from the visit of a single pollinator (Yamada and Guries, 1998). The increase in Forcipomyia larvae and pupae associated with rotten banana stems had shown to produce more cocoa flowers (Young, 1986). The pod yield is influenced by photosynthesis and partition of photo-assimilate (Sounigo et al., 2003).
It is assumed that midge population can be a limiting factor in the pollination of cocoa in addition to the environmental conditions. However, populations of insect pollinators are often severely disturbed by hurricanes through flooding of essential habitat and the widespread loss of existing flowers. Small, poor-flight insects such as midges are likely to be swept away by high winds. Climate variation, particularly changes in rainfall leading to sporadic or less rain, may also affect midges which normally thrive in moist humid environments.
Understanding these ecological dynamics can lead to ways of conserving midge populations and mitigating the effects of global climate change and extreme climatic events. The objective of this study is to examine the relationship between the midge population, flower pollination in Trinidad Selected Hybrids (TSH) cacao, and selected weather variables in several different Caribbean cocoa producing islands.
Characteristics of the study area
A multi-location study during the project period of 2013 to 2016 was conducted on several farms in the islands of Trinidad and Tobago (10.667°N, 61.567°W), and Jamaica (18.1824°N, 77.3218°W) in the Anglo-Caribbean which were previously under natural forest (tropical Montane Crappo-guatecare, fine leaf cocorite, black heart) in altitude 120 to 330 m (Nelson, 2004). The areas experienced annual average temperatures of 26.5 ± 2.09°C, relative humidity of 86.1 ± 12. 6%, and mean monthly rainfall ranging between 19.1 and 235.1 mm (Anon, 2016).
The 4 farms/estates were in Trinidad: Jude Lee Sam Estate (July 2014 - July 2015), San Juan Estate (February 2015 - July 2015), San Antonio Estate (February 2015 - July 2015) in Gran Couva and ECIAF Estate (July 2014 - July 2015) in Centeno. Data was also collected at two (2) sites in Tobago; L’eau Estate (November 2014 - July 2015) and Providence Estate. The 2 estates selected in Jamaica were: Orange River (September 2014 - October 2015) and Richmond (October 2014 – October, 2015).
The cocoa vars. were mainly form the Trinidad Selected Hybrids [TSH] (Maharaj et al., 2011), and the trees were in full reproductive phase. The first flowerings were in early January over a 3 month period, and a second period, depending on the rains, in June. Harvesting usually occurred over a 2 month period around 6 months after the first flowering.
All the islands experienced a bimodal rainfall distribution, with peaks in June and November. The first and second growing seasons typically last from mid-March to mid-July and from mid-August to end of November, respectively. However, this is separated by a short dry spell of about two weeks in September and referred as petite careme. The major dry season starts in mid-December and lasts till end of May, and the climate is marked by high incidence of solar radiation and relatively little variation in day length. All data on temperature and relative humidity were measured using the Data Davis Wireless Vantage Weather Pro [Model E14062 Rainfall data, were taken from the meteorological records from the National Water Resources Agency.
Experimental
Four separate studies were conducted during the period 2013 to 2016 in which the European Union COCOAPOP was executed in the following areas:
1. Insect population dynamics.
2. Cocoa floral phenology.
3. Substrate augmentation trials for culture of cocoa midges (Diptera: Ceratopogonidae), and
4. Generalized linear modelling of weather, midge dynamics and floral phenology.
Study 1: Insect population dynamics
The cocoa insect population dynamics survey was conducted in the 3 islands on 2 well established and managed farms that cultivated the cacao TSH variety under similar agronomic practices. The selected farms were of similar altitude (120 m) and agronomic conditions. The study was conducted over a minimum of fifteen (15) months duration (2013-2015). However, the data analysis was confined to 2 complete flowering seasons over 1 year period.
Insect suction traps (Arnold and Chittka, 2012) were set up in 9 representatives transects within each cocoa estate of the different territories. These traps were secured onto branches of cocoa tress, powered by 9-volt batteries and insects were sucked into vials containing 90% ethanol. Insect samples were collected for 2 days/month for each sample site, labelled, stored properly for analysis in the insectary for other insects and midge count. Collection was timed to the midge life cycle (Figure 1).
Study 2: Cocoa floral phenology
The cocoa floral phenology was conducted on the same cocoa farms for each island. Over 20 mature cocoa trees95 to 12 m tall] with 5 cushions/ tree were randomly selected and labelled within an experimental area not exceeding 500 m2. The study ensured that data were collected from a minimum 100 plants over 3 consecutive flowering years (2013 to 2015). The observations were conducted monthly on each tree using the modified BCCH (Bleiholder et al., 1991) on counts of flowers, buds, number of mature flower buds, open flowers, new pods or cherelles, small pods (5 to 10 mm), medium pods (11 to 60 mm), and large pods (>60 mm); numbers of diseased pods, number of aborted pods (cherelle wilt), and fruit-set over both season.
The BBCH scale was amended to include days from the first day buds become visible [FBV] for each stage and was used to compute the length of each reproductive phase (Figure 2). The BBCH Scale (Bleiholder et al., 1991) and the extended BBCH scale (Hack et al., 1992) covered the 10 principal growth stages numbered 0 to 9 (Table 1). However, for the purpose of this study, only 4 of these stages were considered; namely ‘macro stages’ numbered from 5 to 7.
Study 3. Substrate augmentation trials for culture of cocoa midges (Diptera: Ceratopogonidae)
Two (2) separate studies were conducted on 3 commonly found substrates within the fields to determine if they can augment the midge population as suitable breeding sites (Figures 3 and 4). These studies were confined to Trinidad farms only, as the insectary was located there. The substrates assessed over the 2 crop seasons in 2015 were as follows:
1. Field substrate in-situ assessment, and
2. Field augmentation and insectary evaluation.
Field substrate in-situ assessment: During the cropping season of 2015, four (4) cocoa farms were designated for field manipulation to determine if the substrates had any effect on the midge population dynamics. Three substrates were assessed in heaps viz: Rotted cocoa pod (15 kg) (Figure 4), banana pseudo-stem slices (Figure 2) (15 kg) and cocoa leaf litter, all of which replicated three times per farm. All treatments were moistened (5 L water/heap/weekly). The experimental sites (25 m2/substrate) were laid out as a Latin square (3 × 3) design. During the first 2 months, insect populations were monitored for 2 days per month using a standard suction trap placed in the approximate centre of each area. Cocoa floral phenology was also monitored during the duration of the study which lasted over 6 months.
Field augmentation and insectary evaluation: The field experimentation was conducted at one farm (Gran Couva, Trinidad) and over a five week period [September to October, 2015]. The treatments were the same three substrate treatments with some variations viz., fresh cocoa split pods (35 kg), fresh banana pseudo-stem [35 kg, 10 cm thick slices) and cocoa leaf litter (35 kg) with three replicates of each treatment. The substrates were placed at the base and within the buttress of 15 randomly selected immortelle trees to aid moisture retention. All trees were located within 20 m of one another and from the edges of the field. The substrates samples (2 kg) were evaluated for midge oviposition and larval development from the centre of the piles at 7-day intervals in the insectary. The Ceratopogonid midge larvae after developing in the organic matter were collected using the Berlese Funnel Traps (Dietick et al., 1959). The substrates were inspected for larger midge larvae (Forcipomyia spp.) which are removed from the substrates and placed in a ball of well-decomposed cocoa pod husk with 100 larvae/vial and adequate air-flow and temperature (26°C).
Study 4. Generalized linear modelling of midge dynamics, floral phenology and weather variable
The approach was to determine the relative role of the midge population dynamics and cocoa floral and reproductive phenology, and its interaction under the prevailing weather variables (rainfall and temperature). This study was conducted over the period 2014 to 2015 in the three countries (Trinidad, Tobago, and Jamaica) on two estates per country. The data was collected from previous midge collection and the floral phenology trials and daily weather data (Table 11) for each location. Best fit generalized linear models were developed to determine the interactions and significance.
Data analysis
The count of flowers and other parameters taken were pooled together on each farm, but separate for each location. All count data were transformed when necessary using the square-root (√x + 0.1) before analysis. Regression analysis were used to determine the relationship between weather variables (temperature, relative humidity, rainfall and light intensity) and flower production, and insect population dynamics using the MINTAB statistical package.
Study 1. Insect population dynamics
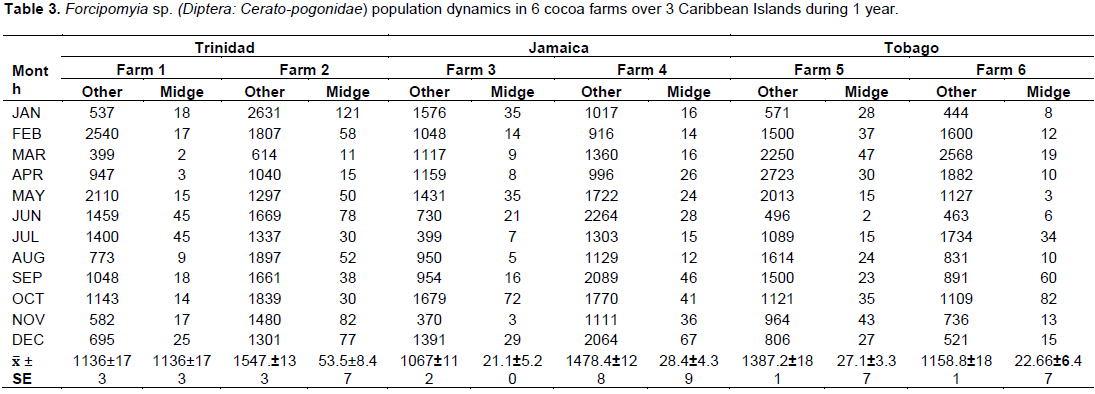
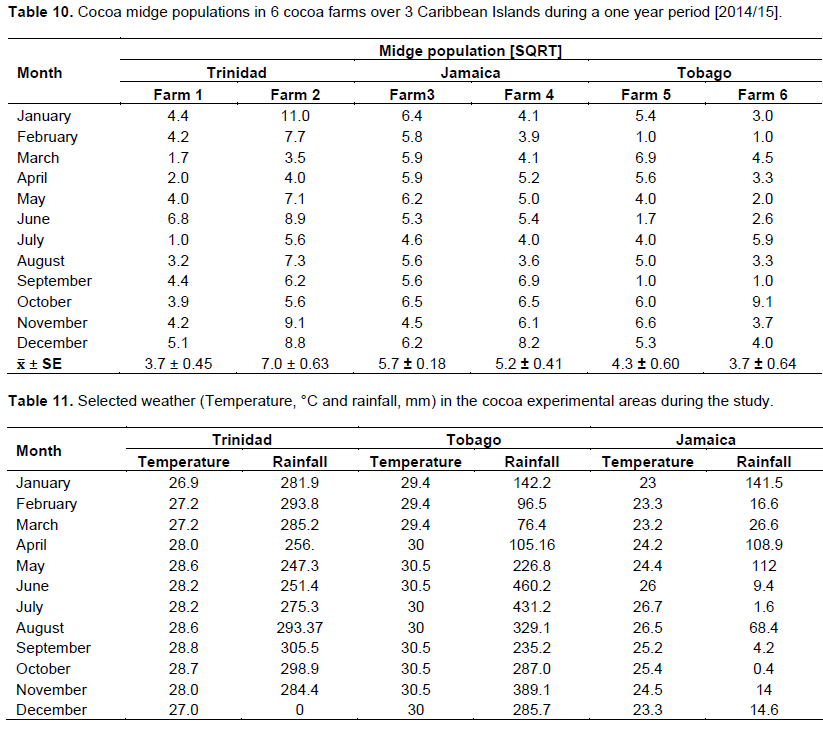
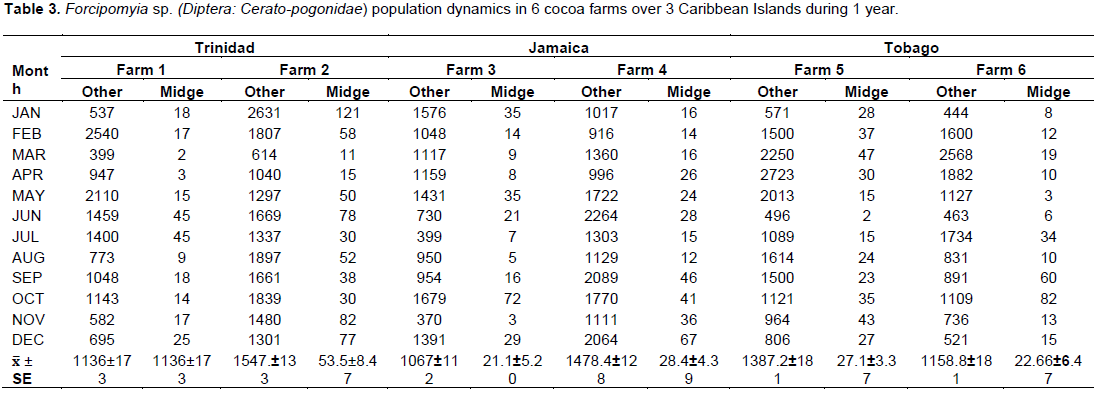
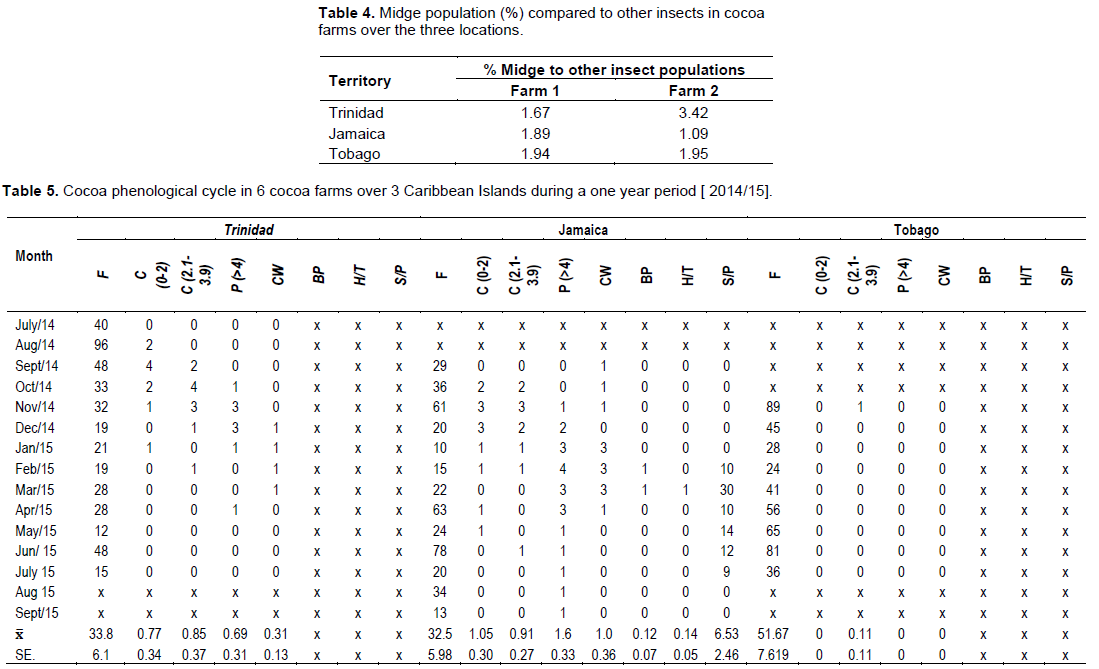
There were significant differences between the monthly midge and other insect’s population and farms over the 3 territories. There were two distinct and observable high populations May/June and November/January. These periods coincided with the new flushes of cocoa flowers (Figure 4) and the higher rainfall patterns. In Trinidad, the seasonal midge population was 19 ± 3.65 and 53.5 ± 8.47 compared to Tobago which varied between 27.1 ± 3.37 and 22.6 ± 6.47, and Jamaica 21 to 28 ± 4.39/ transect site (Table 3).
Study 2. Cocoa floral phenology
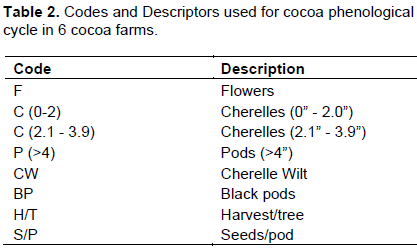
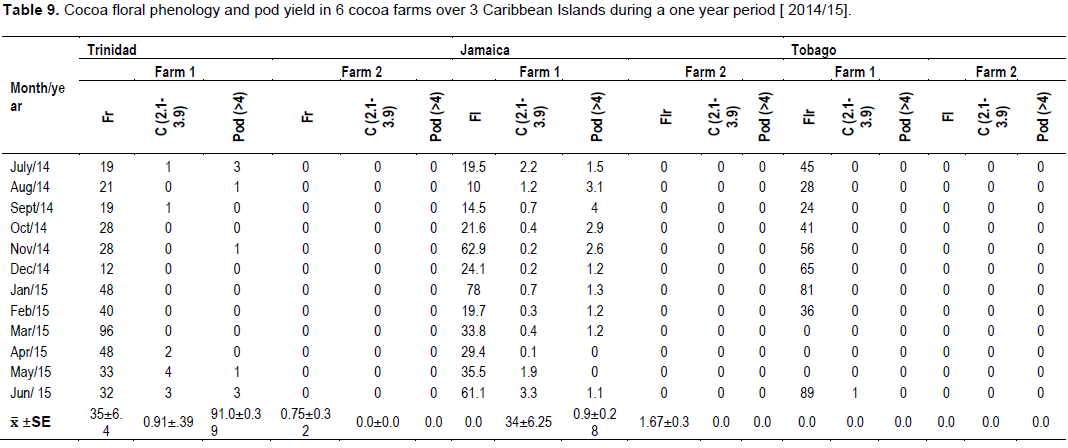
The cocoa floral and reproductive phenology followed a similar pattern (Figure 4) as outlined on the modified model developed by Bleiholder et al. (1991). In Trinidad, the mean number of flowers was 33.6 ± 6.1/cushion, with the highest ranging between 40 to 96 flower/cushion [July/Sept, 2015] and lowest [21 to 28] during Jan/March, 2014 (Tables 2 and 5). This represented the 2 major flowering flushes, which corresponded with the early and late wet seasons, respectively.
Tobago experienced a similar weather pattern to Trinidad during that period (Table 11), and the trees in the study exhibited a slightly higher mean flower/cushion (51.1 ± 7.61). The mature cocoa trees displayed 2 distinct flushes, with the first in November/December 2014 (45 to 89), and a second flush (65 to 81) in the beginning of the wet season (May/June, 2015). The mean flower/ cushion in Jamaica did not vary compared to Trinidad (32 ± 5.98), as the trees were of same variety and age, and also displayed two distinct flusher in Sept/Nov, 2014 (29 to 61) and April/June, 2015 (63 to 78).
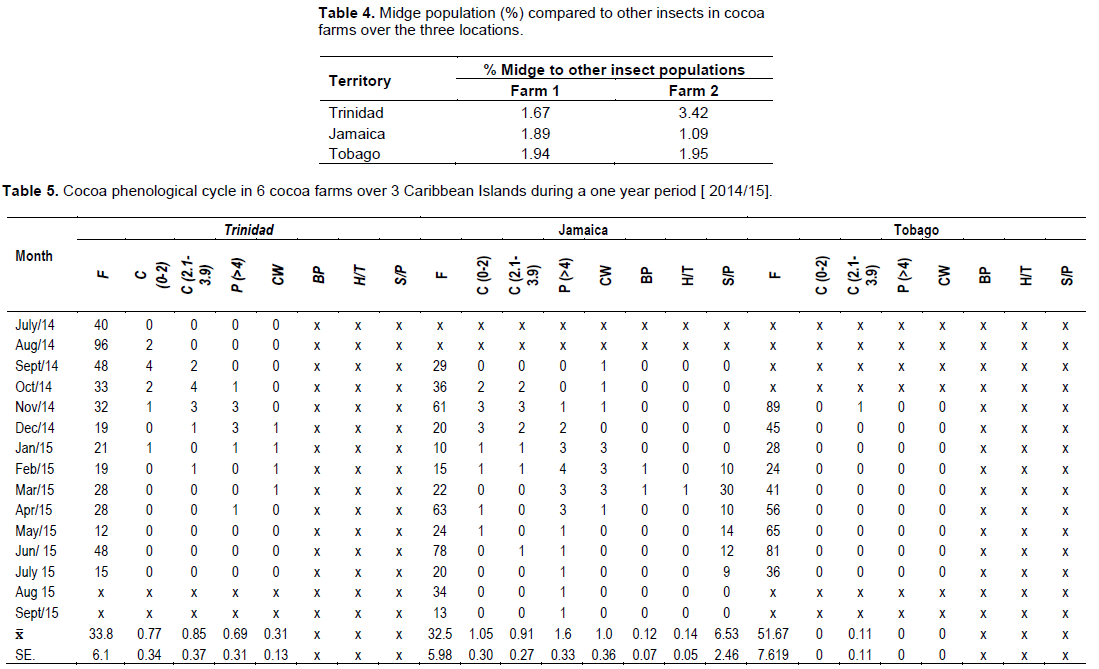
The percentage of flowers that were pollinated and successfully fertilized i.e. ( Flowers→ Cherelles (0”– 2.0”)) were higher in Jamaica (0.91) compared to Trinidad (0.88), and Tobago (0.11). This manifested with a similar pod/cushion yield between countries, with Jamaica (1.5) having a higher pollination/ fertilization, compared to Trinidad (1.0) and Tobago (<1), and was very low for that season (Table 9).
Study 3. Substrate Augmentation trials for culture of cocoa midges (Diptera: Ceratopogonidae)
Trial 1: Field substrate assessment
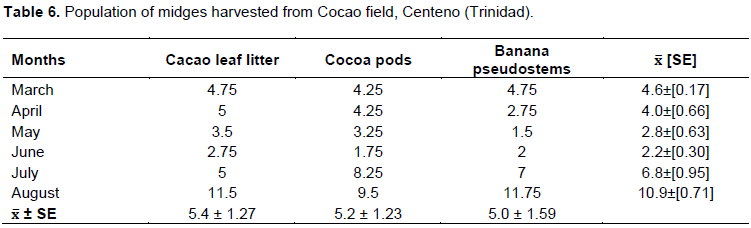
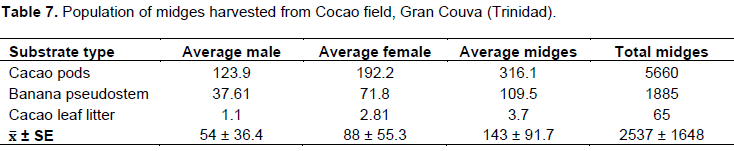
The field trials (Table 6) indicated that there were no variations between the 3 substrates (5.0 to 5.4 ± 1.27) during the experimental period. However, during the wet months of July/August, 2014, the number of midges caught in the suction traps located in the areas of the banana pseudo-stem, and cocoa pod increased, compared to the litter substrate. Similarly, the cocoa leaf litter was not significantly different from pods or pseudo-stems in August.
The number of midges per suction trap in this trial was consistent to the results obtained in the cocoa insect population dynamics studies (2013/14). The study demonstrated that regardless of the quality of the substrate to improve on the feeding and fecundity of midges, the suction trap appeared to have a determining factor, and may not actually reflect the substrate suitability.
Trial 2: Field manipulative and laboratory evaluations
In this study, no suction traps were used, but samples of the substrate were removed and incubated in the insectary, where the emerging larva were counted, and reared to adult. The results in this study are different from Trial 1, and reflected the potential midge population when interventions of substrates are manipulated in the field.
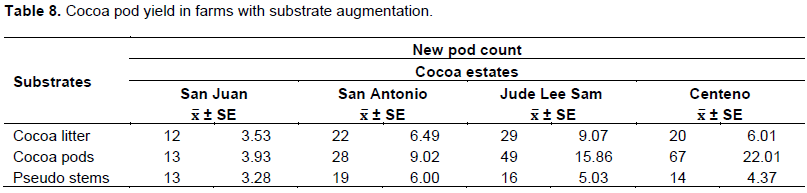
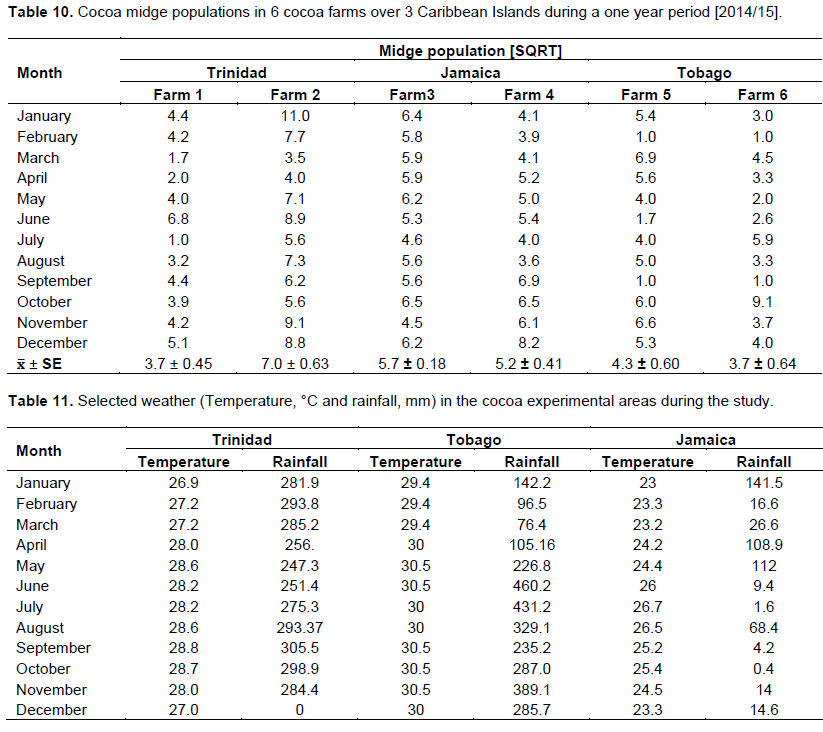
The fresh cocoa pod (Table 7) left to decay was the preferred substrate for the adult midge to feed and continue its reproductive cycle (Figure 1). The total midge population in the cocoa pod was 3 to 4 times higher than the banana pseudo-stem. The data suggested that increasing the breeding sites with augmentation of cocoa pod substrates can increase the midge population (Table 7) dynamics in the field and new pods development (Table 8). Further, the use of suction traps are not effective or a reliable indicator of the true insect population dynamic in the cocoa estates.
Study 4. Generalized linear modelling of midge dynamics, floral phenology and weather variable
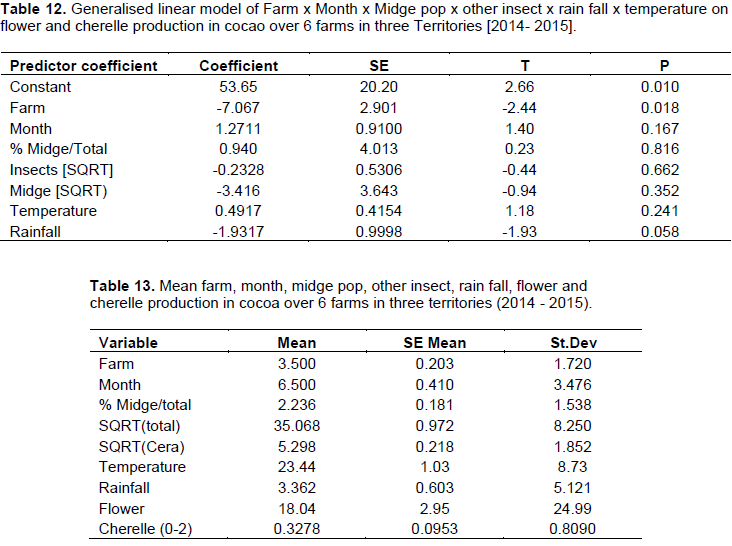
This study involved data transformation and statistical manipulation of observations on the cocoa crop reproductive phenology (Table 9), and midge population dynamics (Table 10) during a one year period, and taking into consideration the prevailing weather variables (Rainfall and Temperature at the different Farm locations) (Table 11).
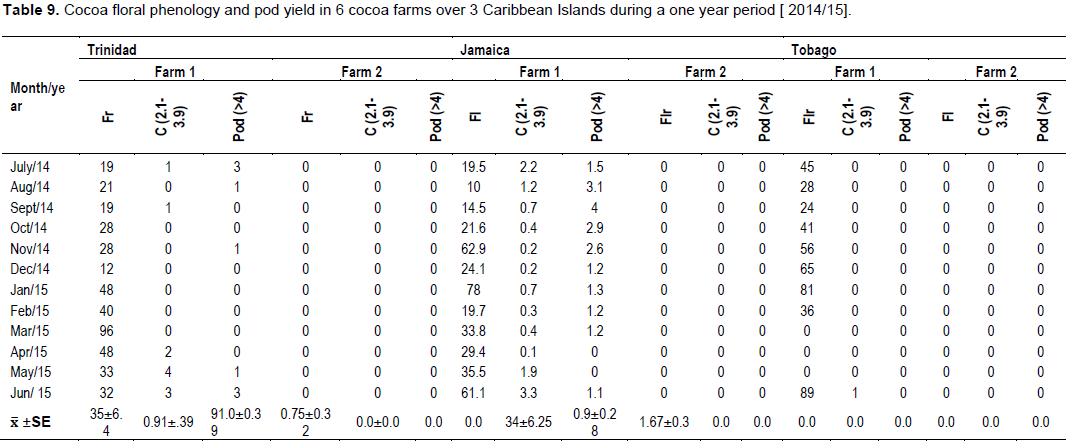
The generalized linear model revealed that there were variations between farms which influenced the yield of flowers and cherelles (Table 12). Also, the variation in rainfall between months, confirmed the bimodal (wet/dry) season which affected flower emergence and pollination into cherelles. The other main variables in the model; midge, other insects, and temperature, were not significant and had no impact on flower and pollination. Additionally, the analysis did not reveal any interactions between any of the independent variables on flower and cherelles (Table 13). The analysis showed that the ratio of flowers to cherelle per cushion varied between territories: Jamaica (33:10), Trinidad (33:0.7), and Tobago (18:0.3). However, this data has to be interpreted in the light of the limitations of the suction trap and the true midge population as reported in Study 3. Further, the numbers of flowers were similar between territories and treatment substrate, and pollination: fertilization ratio was not affected, regardless of the indicator midge population dynamics.
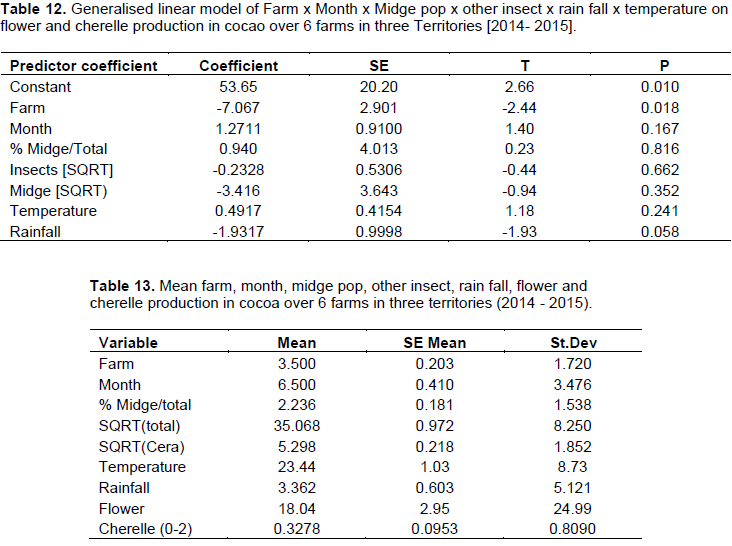
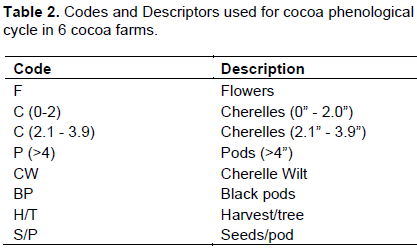
In the augmentation of substrates, freshly harvested cocoa pod waste was the best medium for midge incubation, and was 3 times more desirable than banana pseudo-stem (Table 7). Similarly in the plots with this substrate, there was significantly improved new pod development in all the locations. There is evidence that pod yield increased with increasing midge population as the substrate improved from decaying cocoa leaf litter, to banana pseudo-stem, to cocoa pod (Equation 1).
Generally, the flower emergence per cushion was well within the acceptable expectation for the variety. The midge population was the main pollinator as demon-strated by the substrate study and suggest that it was adequate for the fertilization process, although low. However, the new pod yield was acceptable 12 to 67 (pods/ tree) particularly when the midge was present.
Pound (1933) recommended the minimum yield of pods (50) for a fully grown TSH cocoa tree requiring 7 1/2 pods to 0.5 kg, and yielding 3.5 kg of high class cacao, and 25 for trees 10 to 15 years old. After standardized manual cross-pollination, Bos et al. (2007) obtained 12 fruits/trees and harvested under shade management an average: 27 ± 4% fruits/tree. In Upper Amazon cacao hybrids, 38 to 66% of the trees produced 1 to 10 pods/tree and 7 to 39% had more than 10 pods/tree/year, and high yielding tress and produced up to 180 pods/tree/year (Adomako and Adu-Ampomah, 2003).
According to Mohamed (pers. comm. 2016) the morphology of the cocoa flower does not lend itself easily to insect pollination due to the presence of the staminodes surrounding the style which has a needle-like stigma. The position of the hooded anther opening obtusely from the ovary base makes it difficult to transport the sticky pollen grains downwards. The flower orientation is like a dangling pendulum. The insect will descend directly on the area surrounding the ovary where the nectar glands are located. Per chance if it was crawling out of the flower it passes on the surface of the style, probably depositing pollens on the way out. These pollen will germinate on the surface of the style and affect fertilization. The stigma is no way involved in the fertilization process. The germination of the pollen grains could only occur along the style while the tiny midge is crawling out hence the reason for low pod set.
Krauss and Soberanis (2002) reported that fertilizer improved yields by 11% independent of the disease control measure, but Groeneveld et al. (2010) found that both pollination and resource (shade, fertilizer and water) limitations may cause low fruit : Flower ratios in T. cacao. However, none of the resource availability treatments had a significant effect, while number of mature pods and yield increased non-linearly with pollination intensity up to 200% of current yield levels. Despite an increase of fruit abortion with pollination intensity, T. cacao yield is determined by the number of flowers pollinated. This suggests pollination deficit in crops can be very large and that a better knowledge of pollen and resource limitation is needed to devise adequate pollinator management strategies.
The authors have not declared any conflict of interests.
REFERENCES
|
Adomako B, Adu-Ampomah Y (2003). Assessment of the yield of individual cacao trees in four field trials. In Proceedings of the International Workshop on Cocoa Breeding for Improved Production Systems. 19:41-49.
|
|
|
|
Almeida AAFD, Valle RR (2007). Ecophysiology of the cacao tree. Braz. J. Plant Physiol. 19(4):425-448.
Crossref
|
|
|
|
|
Alverson WS, Whitlock BA, Nyffeler R, Bayer C, Baum DA (1999). Phylogeny of the core Malvales: evidence from sequence data. Am. J. Bot. 86:1474-1486.
Crossref
|
|
|
|
|
Aneja M, Gianfagna T, Ng E (1999). The roles of abscisic acid and ethylene in the abscission and senescence of cocoa flowers. Plant Growth Regul. 27:149-155.
Crossref
|
|
|
|
|
Anon (2016). Weather and climate of Trinidad and Tobago. Available at: https://www.ttconnect.gov.tt/gortt/ portal/ttconnect/!ut/p/a1/jdDBCoJAFAXQb-kHfE9lIpcTmKmLUCl1NqEwTZbNxDhkn5-
|
|
|
|
|
Arnold SE, Chittka L (2012). Illumination preference, illumination constancy and colour discrimination by bumblebees in an environment with patchy light. J. Exp. Biol. 215(13):2173-2180.
Crossref
|
|
|
|
|
Bartley BGD (2005). The genetic diversity of cacao and its utilization. CABI Publishing, Wallingford, UK.
Crossref
|
|
|
|
|
Bleiholder H, Kirfel H, Langel¨uddeke P, Stauss R (1991). Codificac¸ao˜unificada dos est’adios fenol’ogicos de culturas e ervas daninhas. Pesquisa Agropecuaria Brasileira, 26:1423-1429.
|
|
|
|
|
Bos MM, Steffan-Dewenter I, Tscharntke T (2007). Shade tree management affects fruit abortion, insect pests and pathogens of cacao. Agric. Ecosyst. Environ. 120(2):201-205.
Crossref
|
|
|
|
|
Cazorla IM, Aidar T, Milde LCE (1989). Perfis do lançamento foliar, da floração, da bilração e de estágios do fruto do cacaueiro no Estado da Bahia, no período de 1987/1988. In: Boletim Técnico, Ceplac/Cepec, Ilhéus, Brasil, 58p.
|
|
|
|
|
Cheesman EE (1944). Notes on the nomenclature, classification and possible relationships of cacao populations. Trop. Agric. 22:144-159.
|
|
|
|
|
Daymond AJ, Hadley P (2004). The effects of temperature and light integral on early vegetative growth and chloroplyll fluorescence of four contrasting genotypes of cacao (Theobroma cacao). Ann. Appl. Biol. 145:257-262.
Crossref
|
|
|
|
|
Dias LAS, Kageyama PY, Castro GCT (1997). Divergência genética multivariada na preservação de germoplasma de cacau (Theobroma cacao L.). Agrotrópica 9:29-40.
|
|
|
|
|
Dietick EJ, Schlinger EI, Van den Bosch R (1959). A new method for sampling arthropods using a suction collecting machine and modified Berlese funnel separator. J. Econ. Entomol. 52(6):1085-1091.
Crossref
|
|
|
|
|
Garcia JR (1973). Estudo de alguns índices de crescimento e produtividade para seleção juvenil em híbridos de cacau. Turrialba, IICA, M.Sc. dissertation.
|
|
|
|
|
Groeneveld JH, Tscharntke T, Moser G, Clough Y (2010). Experimental evidence for stronger cacao yield limitation by pollination than by plant resources. Perspect. Plant Ecol. Evol. Syst. 12(3):183-191.
Crossref
|
|
|
|
|
Hasenstein KH, Zavada MS (2001). Auxin modification of the incompatibility response in Theobroma cacao. Physiol. Plant. 112:113-118.
Crossref
|
|
|
|
|
Krauss U, Soberanis W (2002). Effect of fertilization and biocontrol application frequency on cocoa pod diseases. Biol. Control 24(1):82-89.
Crossref
|
|
|
|
|
Lachenaud P, Mooleedhar V, Couturier C (1997). Les cacaoyers spontanés de Guyane. Nouvelles prospections. Plant Rech. Dév. 4:25-30.
|
|
|
|
|
Lanaud C, Sounigo O, Amefia YK, Paulin D, Lachenaud P, Clement D (1987) Nouvelles données sur le fonctionnement du système d'incompatibilité du cacoyer et ses conséquences pour la sélection. Café Cacao Thé 31:267-277.
|
|
|
|
|
Maharaj K, Maharaj P, Bekele FI, Ramnath D, Bidaisee GG, Bekele I, Sankar R (2011). Trinidad selected hybrids: An investigation of the phenotypic and agro-economic traits of 20 selected cacao cultivars. Trop. Agric. 41(3216): 040175-11.
|
|
|
|
|
Motamayor JC, Risterucci AM, Lopez PA, Ortiz CF, Moreno A, Lanaud C (2002). Cacao domestication. I. The origin of the cacao cultivated by the Mayas. Heredity 89:380-386.
Crossref
|
|
|
|
|
Nelson HP (2004). Tropical forest ecosystems of Trinidad: Ecological patterns and public perceptions. Ph.D. Dissertation, University of Wisconsin, Madison.
|
|
|
|
|
Pound FJ (1933). Criteria and methods of selection in cacao. Second Ann. Rep. Cacao Res. 1932:27-29.
|
|
|
|
|
Sena GAR, Kozlowski TT (1986). The effects of flooding on water relations and growth of Theobroma cacao var. catongo seedlings. J. Hortic. Sci. 61:265-276.
Crossref
|
|
|
|
|
Sounigo O, Lachenaud P, Bastide P, Cilas C, N'Goran J, Lanaud C (2003). Assessment of the value of doubled haploids as progenitors in cocoa (Theobroma cacao L.) breeding. J. Appl. Genet. 44:339-353.
|
|
|
|
|
Yamada MM, Guries RP (1998). Mating system analysis in cacao (Theobroma cacao L.). Agrotrópica 10:165-176.
|
|
|
|
|
Young AM (1986). Habitat differences in cocoa tree flowering, fruit-set, and pollinator availability in Costa Rica. J. Trop. Ecol. 2(02):163-186.
Crossref
|
|