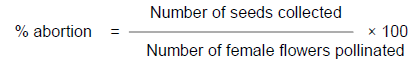ABSTRACT
Controlled crosses and selection of promising cassava genotypes were carried out at Cassava Breeding Programme of National Root Crops Research Institute (NRCRI), Umudike, Nigeria, in 2010. A 3 x 3 North Carolina mating design was used to generate nine F1 population. Out of 7,044 seeds expected from all the pollinated flowers, only 1,102 seeds (15.6%) were harvested. Majority of the pollinated flowers did not develop into fruits due to abortion that happened shortly after pollination. Family TMS 98-0002 x TMS 05-0473 (B3) had the highest seed abortion rate of 75.5% whereas family TMS 98-0505 x TMS 01-1368 (B7) had the lowest seed abortion rate of 25.3%. Seeds generated from viability test were sown in pots and maintained for 28 days in the screen house. Seed germination ranged from 15.5 to 80.9% with a mean of 43.19%. The emerged seedlings were hardened and transplanted to the field alongside their parents. Four hundred and sixty four progeny survived and were evaluated 12 months after planting (MAP). There were very high levels of variability in the segregating F1 progeny for all the traits studied that will be useful for cassava breeding in Nigeria. Subsequent evaluations of promising clones have reached advanced stage for release to farmers in Nigeria.
Key words: Cassava, botanical seed, germination, pollination, variation.
Cassava is one of the leading staples in the world with a global total production of 252 million metric tonnes (FAOSTAT, 2012). Though with the current national estimate of 55 million cassava production in 2013 (NRCRI, 2013), the cassava genotypes grown by farmers in Nigeria are mostly local cultivars and few improved released varieties by NRCRI Umudike in collaboration with International Institute of tropical Agriculture (IITA), Ibadan (NRCRI, 2013). Improving the local cultivars (landraces) that flower requires a hybridization programme to generate hybrid progeny for recombination and selection. Population improvement and recurrent selection in cross-pollinated crops such as cassava progressively increases the frequencies of genes for specific desirable traits. However, the success of a breeding programme depends largely on the choice of progenitors/parents used. Parental genotypes are usually selected on the basis of their performance or the performance of their F1 progeny (Ceballos et al., 2004). For instance, in maize, selection of parental genotypes to produce F1 hybrids is usually based on the performance of their progeny (Derera et al., 2000).
Cassava breeders have traditionally used performance per se of parental genotypes (Ceballos et al., 2004). Also, in the current study, parental genotypes or progenitors were selected based on their performance per se. The selected local cultivars, though with low in pro-vitamin A (PVAC) and susceptible to pests and diseases, have good root qualities and are popular with the farmers in the area of study (Njoku et al., 2014). The NRCRI improved varieties used to cross with the local cultivars are early bulking, high PVAC and resistant to pests and diseases but lacked high dry matter and yield attributes acceptable to farmers (Njoku et al., 2014). It was assumed that crossing the two groups (local cultivars and NRCRI varieties), would result to new genotypes, which combine high PVAC with other agronomic acceptable root qualities by Nigerian farmers.
In this study, the NC II mating design was used to generate the progeny from crosses between the two groups of parents (local cultivars versus improved varieties). Several researchers have used this breeding design, for example, in sugar cane (Hogarth et al., 1981), variety crosses in maize (Derera et al., 2000) and even feed conversion in broiler rabbits (Dedkova et al., 2002). In cassava, the design has been used to study resistance to cassava mosaic disease (Lokko et al., 2004).
However, experience has shown that cassava botanical seeds have low germination rate both in nursery and in the field (Ceballos et al., 2004; Kawano, 1978; Njoku et al., 2014) which eventually leads to loss of possible important genes, as well as wastage of land. Moreover, standard germination test by floatation do not always adequately predict seedling emergence under greenhouse and field conditions, and total percentage loss has not yet been established. The aims of the study were to investigate the effect of cassava genotypes on seed set, seed germination, seed abortion, and to ascertain the level of variability among progenitors in cross combination.
Evaluation site and germplasm source
Six cassava progenitors were selected and evaluated at National Root Crops Research Institute (NRCRI) Umudike research field, which lies within the humid rainforest agroecology of Nigeria (Njoku et al., 2014). They comprised three widely cultivated white root cassava cultivars and three yellow root cassava genotypes (one released in 2011 while others are yet to be released). The six cassava genotypes which were developed at NRCRI, Umudike have good agronomic, nutritional and disease resistance attributes. All the cultivars were selected based on their characteristics of good performance in terms of dry matter, pest and disease resistance, plant architecture, nutritional quality and flowering ability (Table 1).
The six selected progenitors were then planted in a crossing block at NRCRI, Umudike in May, 2010, in a 3 × 3 North Carolina mating design. Ridges were made with spacing of 1.0 m intra-row and 1.0 m inter-row. Hand weeding was performed as required. Controlled hand pollination was done according to the standard procedure described by Kawano (1980).
Plants were observed for signs of flowering each morning. Pollination bags were used to enclose flowers about to open to prevent fertilization by stray pollen upon opening. Pollen from the corresponding male parent was collected in the morning (7 to 8 am) and pollination was done later in the day between 11 am and 2 pm after female flowers had opened by dusting pollen on the stigma of a matching female. One male flower was used to pollinate up to three female flowers. After pollination the female flowers were covered with the bags for one to two weeks. Each flower branch was marked by a tag indicating cross combination with female parent listed first and followed by date of pollination and pollinator. Developing fruits were covered with netting bags three to four weeks after pollination to catch the dehisced seeds (Figure 1). Seeds were collected after two months, sorted, labeled and stored till the time of sowing. Percentage seed set was determined by dividing total number of flowers pollinated with total number of seeds collected per cross. For pollinated fruits, it was assumed that every fruit had three ovules.

Germination of hybrid seeds
Seeds collected from each of the nine cassava populations were subjected to viability test and those that submerged were collected and sown in perforated plastic trays filled with unsieved sandy loam soil collected from an undisturbed field at western farm of NRCRI, Umudike. The plastic trays used were lined with cotton wool to prevent loss of soil particles through the holes underneath the trays. The soil in each tray was watered to field capacity and left overnight (24 h) before seeds were sown about 2 cm deep in the soil. The soil surface was covered with black polythene nylon to raise the soil temperature and thereby aid in germination of the seeds. The number of seeds from each population determined the number of trays used. The number of seeds that passed viability test per cross ranged between 45 and 385 seeds. Percentage germination and seed set in each population were used to determine crossability among the 6 parents used in the hybridization process. The trays were watered on a daily basis for 28 days when the emerged seedlings had reached 25 cm with a minimum of 3 leaves. In other to acclimatize the seedlings to vagary of weather, hardening of seedlings was done for 5 days by reducing watering and exposing them to ambient temperature before transplanting to the main field (Figure 2).

Transplanting of hybrids to field
Vegetative stem cuttings of 25 cm long each, about 4 cm thick and a minimum of 5 nodes in each stem cutting of the six progenitors (TMS 01-1368, TMS 05--1636, TMS 05-0473, TMS 98-0002, TMS 98-0505 and TMS 97-2205) were dipped into a bucket solution of 10 ml basudin (an emulsifiable concentrate insecticide containing 60% Diazinon) to 10 L of water for 5 min and allowed to dry overnight before planting to prevent termite attack. The progenitors were also used as checks in evaluating their hybrid progeny.
The seedlings (473) raised from hybrid seeds in each population were transplanted to the field alongside their parents, in 2010 at a spacing of 0.75 × 1 m for individual plant evaluation at NRCRI, Umudike. Because of late establishment of rainfall, the transplanted plants and their parents were watered twice daily (500 ml/plant) for 38 days to supplement the soil moisture. Hand weeding was carried out as required.
Data collection and analysis
The number of seedlings that emerged was recorded on weekly intervals (7, 14, 21 and 28 days after planting DAP) for each family until 28 days after planting (DAP) when there was no further emergence of seedling. The percentage germination for each hybrid population was calculated as emerged seedlings divided by the number of seeds sown multiplied by 100. Variability in percentage germination of seeds developed in different crosses involving each male or female parent was estimated. All data were analyzed using Genstat Version 3 (Payne et al., 2008), SAS 9.2 Edition and SPSS 15.0. The REML linear mixed model was used to analyze family and progeny data at seedling stage. Mean, minimum, maximum, standard deviation and coefficient of variation (CV) were calculated for each hybrid population.
Seed set and seed abortion
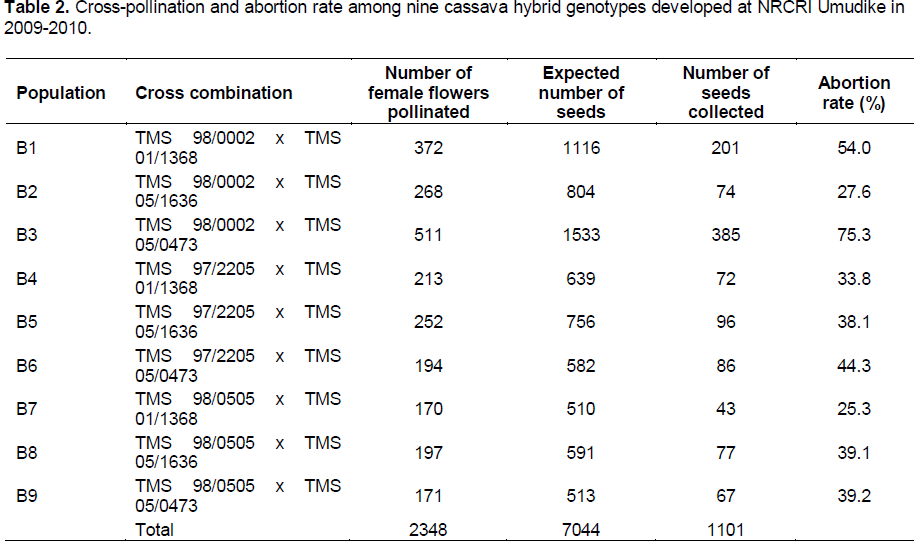
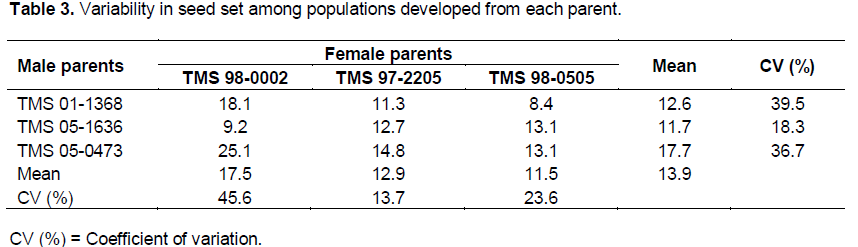
Table 2 shows the number of female flowers pollinated and number of seeds collected in each cross during the development of the hybrid populations at Umudike between November 2009 and February 2010. Out of 7, 044 seeds expected from all the pollinated flowers (2348) with three possible seeds in each trilocular fruit, only 1102 seeds (representing 15.6% of the maximum number of seeds expected) were collected. Majority of the pollinated flowers did not develop into fruits. This may be probably because of the late in pollination which was off-season (November to February, 2010) and a lot of seed abortions. However, less than one seed was obtained per pollination on average in this study. This is far below the expectation according to submission by Ceballos et al. (2004) who stated that one or two seeds are obtained per pollination on average. The highest percent seed set was recorded in the cross between TMS 98-0002 and TMS 05-0473 which were the best female and male parents in terms of seed set respectively. Population B3 had the highest abortion rate of 75.3%, followed by B1 (54.0%), while B7 had the lowest rate of 25.3%.
However, the dry season and harmattan aids drying of the seeds, hence, the drying period for the seeds was reduced. Opening of mature flowers and drying of matured fruits depend on environmental conditions (Ceballos et al., 2004; Olasanmi et al., 2014). Number of seeds per female flower is genotype dependent; therefore, selection of highly fertile genotypes as female parents is a critical factor.
The number of parents used in this study may not reflect the wide genetic variations often found among clones used as parents in cassava breeding programme. However, large differences in coefficient of variation for seed set among the parents and significantly better performance of TMS 98-0002 and TMS 05-1636 in terms of seed set than other female and male parents respectively, suggest genetic differences in the crossability of these clones (Table 2). This is also in agreement with Olasanmi et al. (2014) who reported that both maternal effect and compatibility between male and female parents have influence on percent seed set in cassava, and it may be necessary to investigate the potential seed crossibility of male and female parents used in the development of recombinant seeds in cassava breeding programmes. This will help cassava breeders to choose parental varieties with high seed set capability and also maximize the resources in terms of time, labour and funds needed.
Coefficient of variation (CV) for seed set was found to be low for TMS 97-2205, followed by TMS 98-0505 (Table 3). This suggests that seed set is more stable in these two parents. Although the number of parents analyzed may not reflect wide genetic variation often found among clones used as parents in breeding programmes, results based on CV indicate genetic differences in the crossability of these clones. There was no seed loss due to pest destruction because fruits were protected by the bags and the field was well maintained throughout the course of the hybridization programme. There was non-significant but very high positive correlation between number of female flowers pollinated and number of seeds harvested in each population (r = 0.97, p = 0.4022, n = 9). This shows that only 39.5% of the rate of seed set can be predicted using the number of female flowers pollinated. Also this non-significant correlation indicates that success of pollination programmes in cassava cannot be accurately predicted from the number of flowers pollinated (Ceballos et al., 2004; Olasanmi et al., 2014).

Hybrid seed germination
The number of seed sowed per cross ranged from 32 to 337 with a mean of 106.56, while the lowest number of seeds (32) was observed in family TMS 98-0505 x TMS 01-1368. The first sprouting was noticed and recorded four days after sowing (DAS) in population B3, followed by population B1 and B2 after 7 days. Also, sprouting ceased in populations B6 and B7 in 21 DAS but continued till 28 DAS in population B1, B2, B3, B4, B5, B8 and B9 respectively. There was a very high positive correlation between number of seeds sowed and number of emerged seedlings in each population (r = 0.93). Maximum temperature ranged between 31 and 34°C while the minimum temperature ranged between 22 and 24°C during hybridization, development and drying of the fruits and germination of seeds (November 2009-April 2010). The relative humidity ranged between 72 and 78% during the same period.
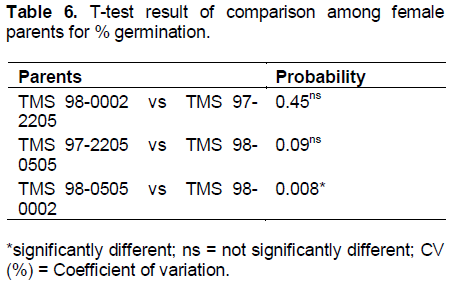
The highest germinability of 80.9 was recorded in B2 while B7 had the lowest value of 15.6% (Table 4). The lowest coefficient of variation (CV) of 14.6% for germinability found among the crosses involving TMS 98-0505 (Table 5) suggests that germinability is stable among the crosses. T-test results showed significant difference in the mean percent germinability among crosses developed from TMS 98-0505 and those from TMS 98-0002. However, there were no significant differences in percent germinability among crosses from TMS 98-0002 and those from TMS 97-2205 as well as among crosses from TMS 97-2205 and those from TMS 98-0505 (Table 5 ).
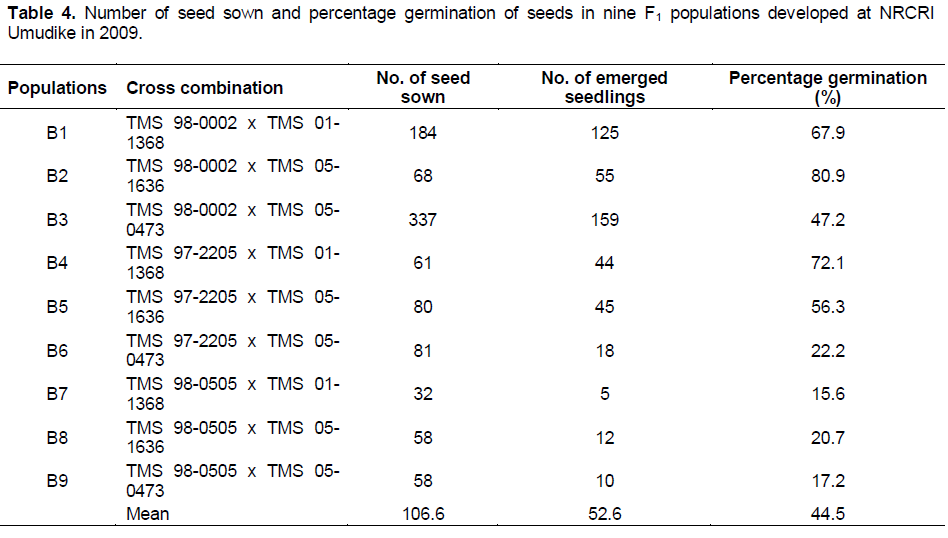
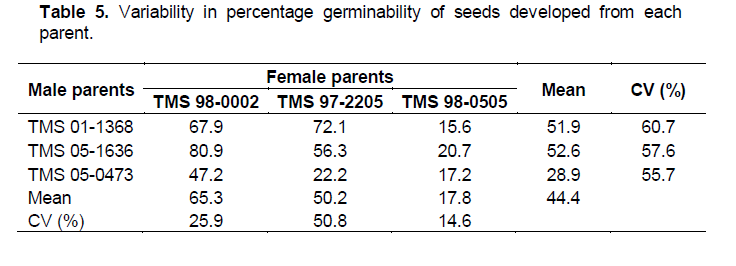
Generally, the differences observed in the rate of germination of hybrid seeds in the nine populations may also be another consequence of compatibility between the two parents used to generate each population since the germination process was carried out at the same time under the same weather conditions. This is also in agreement with Olasanmi et al. (2014) who suggested that one can predict to a large extent the number of seedlings expected in a breeding programme using the number of seed generated (Table 6).
Most breeding programs start with the evaluation of available genetic variability, and how easily this variability can be fixed in the genotypes is very important. After evaluation of all the yellow fleshed roots and white fleshed roots cultivars in NRCRI cassava breeding programme, 6 cultivars were selected (3 white and 3 yellow) based on their per se performance in terms of high yielding, pests and diseases resistance, high dry matter, high carotene amongst others. About 90% of the progeny have higher beta-carotene than their parents and an appreciable quantity of dry matter content. This diversity can be utilized by breeders to develop beta-carotene rich cassava varieties and improve existing varieties. Apart from combating vitamin A deficiency amongst heavy cassava consumers in Nigerians, beta-carotene rich cassava varieties can also be used to supplement yellow maize in poultry feed.
The authors have not declared any conflict of interest.
REFERENCES
|
Ceballos H, Iglesias AC, Pérez JC, Dixon AGO (2004). Cassava breeding: Opportunities and challenges. Plant Mole. Biol. 56: 506-516. |
|
|
|
Dedkova L, Mach K, Mohsen A (2002). Analysis of growth and feed conversion in broiler rabbits by factorial crossing. Czech J. Anim. Sci. 47:133-140. |
|
|
|
Derera J, Denash GP, Pixley KV (2000). Resistance of maize to the maize weevil: II. Non-preference. J. Crop Sci. 9:441-450. |
|
|
|
Food and Agriculture Organization of the United Nations (FAO). 2012. Standardized food balance sheet from the FAO Basic foodstuff services, Commodities and Trade Division. Rome, Italy: FAO. |
|
|
Hogarth DM, Wu KK, Heinz DJ (1981). Estimating genetic variance in sugarcane using a factorial cross design. Crop Sci. 21:21-25.
Crossref |
|
|
|
Kawano K (1980). Cassava. In: Fehr WR and Hadley HH (eds.) Hybridization of crop plants. ASA and CSSA, Madison, WI, pp. 225-233. |
|
|
Kawano K, Narintaraporn K, Narintaraporn P, Sarakarn S, Limsila A, Limsila J, Suparhan D, Watananonta W (1978). Yield improvement in a multistage breeding program for cassava. Crop Sci. 38:325-332.
Crossref |
|
|
|
Lokko Y, Dixon AGO, Offei SK (2004). Combining ability analysis of field resistance in cassava to the African cassava mosaic disease. Int. Crop Sci.
|
|
|
|
Lokko Y, Danquah EY, Offei SK, Dixon AGO, Gedil MA (2005). Molecular markers associated with a new source of resistance to the cassava mosaic disease. Afr. J. Biotechnol. 4(9):873-881. |
|
|
Njoku DN, Vernon GE, Offei SK, Asante IK, Egesi CN, Danquah Y (2014). Identification of Pro-vitamin A Cassava (Manihot esculenta Crantz) Varieties for Adaptation and Adoption through Participatory Research. J. Crops Improv. 28(3):212-120.
Crossref |
|
|
|
National Root Crops Research Institute (NRCRI) (2013). Annual report on genetic improvement of root and tuber crops in Nigeria. Umudike, Nigeria: National Root Crops Research Institute. |
|
|
|
Olasanmi B, Akoroda MO, Egesi C, Okogbenin E, Fregene M (2014). Cross-compatibility Among Six Improved Cassava (Manihot esculenta Crantz) Varieties. J. Root Crops 40:15-22. |