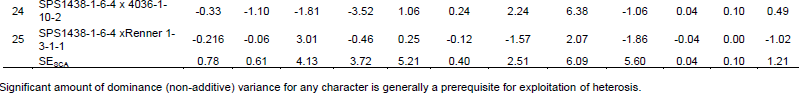ABSTRACT
An 8 x 8 diallel cross mating design with the parents and F1s was used to estimate gene action and combining ability for yield and yield characters in sesame (Sesamum indicum L.). The experiment was conducted at the National Semi-Arid Resources Research Institute (NaSARRI), Serere, Uganda during the second season of 2013. The traits involved: days to 50% flowering, days to maturity, height to first branch, height to first capsules, plant height, length of the capsule fruiting zone and yield per plant. The genetic variance of combining ability was separated into general (GCA) and specific (SCA) combining ability variance components. Baker’s ratio, coefficients of genetic determination (CGD) were determined for narrow and broad sense heritabilities. High GCA effects were recorded for days to flowering, height of first capsule and number of branches, branches per plant and capsule length. Desirable SCA effects were recorded for Sesim2//5181 x Renner 1-3-1-1 for reduced height of first branch (-9.48*), Sesim1 x Sesim2 for number of branches per plant (0.901*), capsules on branches (20.75**) and yield per plant (2.42*). High Baker’s ratio was recorded for most of the traits except for yield per plant (0.233). CGDbs was high for most of the traits except for days to maturity (0.064), plant height (0.346), capsules on main stem (0.358) and capsule width (0.286). These results suggested that both additive and non-additive gene actions played a greater role in these traits. Sesim 1 had the highest number of desirable traits scored for GCA effects and could be considered as a parent for crossing to produce desirable progeny. Sesim 1 x Sesim 2, Sesim 1 x Ajimo A1-5 and Sesim2//5181 x SPS1438-1-6-4 recorded positive significant SCA effects for yield per plant and therefore they could be recommended for hybrid seed production.
Key words: Baker’s ratio, coefficient of genetic determination, gene effects, yield, yield components, sesame.
Sesame (Sesamum indicum L.), commonly known as gingelly, til, benniseed and simsim is a member of the order Tubiflorae and family Pedaliaceae (Ahmed and Adam, 2014). It is normally called ‘Simsim’ in Eastern Africa and it is a traditional and important oilseed crop in Uganda (Auckland, 1970). In 2011, Uganda was the 7th in world sesame production with 173,000 tons of seed. The highest in the world was Myanmar, India and China with 861,573; 769,000 and 605, tons, respectively (FAOSTAT, 2015). In Uganda, sesame ranks second in importance to
groundnuts and it is predominantly grown in the North and North Eastern parts of the country, although there is a considerable amount grown in Eastern Uganda (Anyanga and Obong, 2001). Sesame is gaining considerable importance in Uganda on account of its economic value and especially its export potential, as a non-traditional export crop from Uganda. The national yield is reported to be about 600 kg/ha (FAOSTAT, 2009). Brigham (1985) reported average yields of 1,564 kg/ha in USA. This disparity in yields is caused by various constraints including lack of improved varieties. It is called the “Queen of oil seeds” because of its excellent quantities of the seed, oil and meal. Sesame is highly nutritive (oil 50% and protein 25%). It is an important annual oilseed crop in the tropics and warm sub-tropics where it is mainly grown in small plots as source of edible oil and one of the ingredients in food products. The seed is also rich in protein, vitamins including minerals and lignans such as sesamolin and sesamin (Moazzami and Kamal-Eldin, 2006). Sesame oil has medicinal and pharmaceutical value and is being used in many health care products (Coulman et al., 2005). The seed contains 50 to 60% oil and 25% protein with antioxidants lignans such as sesamolin, sesamol, sesamin which impart to it a high degree of resistance against oxidative rancidity and gives it a long shelf life (Ashri, 1989). It has been used as an active ingredient in antiseptics, bactericides, viricides, disinfectants, moth repellants and anti-tubercular agents (Bedigian et al., 1985). It is a source of calcium, tryptophan, methionine and many minerals (Johnson et al., 1979).
Although, it is a self-pollinated crop, there is some extent of cross-pollination which has a good scope for exploitation of heterosis. Further, an understanding of the combining ability and gene action is a prerequisite for any successful breeding programme. For breaking the yield barrier and evolving varieties with high yield potential, it is desirable to combine the genes from genetically diverse parents. There are several techniques for evaluating the varieties or cultivars or lines in terms of their combining ability and genetic makeup. Of these, Diallel, partial Diallel and line x tester techniques are in common use.
The concept of combining ability analysis gives precise estimates of the nature and magnitude of gene actions involved in the inheritance of quantitative characters, which facilitate the identification of parents with good general combining ability effects and crosses with good specific combining ability effects.
Successful breeding programme depends on the variability available among the different genotypes and in-depth understanding of the underlying gene action and genetic architecture of traits related to yield. Selection of parents based on their performance per se alone may not always be a sound procedure, since phenotypically superior genotypes may yield inferior hybrids and/or poor recombinants in the subsequent segregating generations (Banerjee and Kole, 2009). It is very important to identify parents with high GCA value for the trait to be improved (Banerjee and Kole, 2009). Griffing (1956) provides an efficient estimation of combining ability and the nature of gene action involved. General combining ability is largely due to additive genetic effects and additive x additive epistasis, while specific combining ability is largely a function of non-additive dominance and other types of epistasis.
Knowledge on the nature of the combining ability effects and their resulting variances has a paramount significance in deciding on the selection procedure for exploiting either heterosis or obtaining new recombinants of desirable types in sesame (Solanki and Gupta, 2003). Combining ability is helpful to identify the desirable parents for producing better recombinants (Muhammad and Sedeck, 2015). No breeding research work has been done in Uganda regarding combining ability and gene action on sesame. This can also be observed by limited publications on sesame breeding work in Africa.
The objectives of the present study were to estimate general and specific combining ability, and assess the nature and magnitude of gene action controlling the inheritance of seed yield and some of its components in Uganda’s sesame germplasm.
Experimental site
The trial was planted at the National Semi-Arid Resources Research Institute (NaSARRI), Serere district, Uganda (Latitude 1°31’N; Longitude 33°27’E;and altitude 1,140 masl) during the second rains season of 2013. Uganda has rainy seasons with the first rainy season planted between March to May and the second season planted between July to September. The climate is sub-humid with a mean rainfall of 1350 mm which is bi-modally distributed with higher peak in April-May and lower peak in September-October. There is a very dry season from December to March, during which the temperatures rise to a maximum of around 95°F. The institute has mainly sandy loam soils and some clay loams with tropical savannah grassland vegetation.
Experimental materials
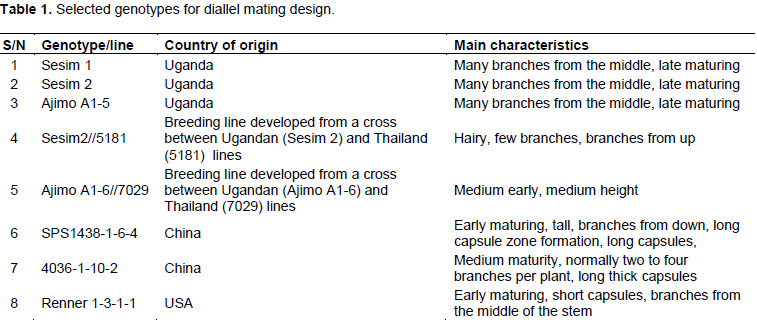
Eight diverse sesame genotypes including local and exotic lines (Table 1) that were previously characterized morphologically in the breeding programme at NaSARRI, Serere and showed contrasting traits were crossed in an 8 x 8 diallel mating design without the reciprocals resulting in 28 F1 combinations. These hybrids along with their parents were grown in a completely randomized block design with three replications. The plots were single rows in 2 m length with spacing of 30 x 10 cm. In the experimental design, Sesim 2 variety was used as border row planted at the beginning and the end of each replication to control the border effects.
Data collection
The data recorded were: number of days to 50% flowering, days to physiological maturity, number of primary branches per plant, height to first branch on the main stem, height to first capsule on the main stem, length of the capsule formation zone, number of capsules on the branches, number of capsules on the main stem, total number of capsules per plant, plant height and seed yield per plant. Five plants were selected from the centre of the row for data collection except for days to 50% flowering and days to maturity which was on the plot basis.
Statistical analysis
The characters under study were subjected to analysis of variance for a completely randomized block design on an entry mean basis. The combining ability analysis was carried out following Griffings (1956) method 4 which includes only the direct F1 crosses without parents and reciprocals. The estimates of GCA effect for the parents and the SCA effects for the crosses were calculated according to Singh and Chaudhary (1985).
The model followed was:
Yij= µ + gi + gj + sij + 

 ɛijkl
É›ijkl  i,j=1,…p,
i,j=1,…p,
Where, Yij is the observation of cross (xij), µ is the population mean, gi and gj are the general combining ability effect for the ith and jth parents, Sij is the specific combining ability effect of the cross between the ith and jth parents such that Sij = Sji and É›ijkl is the experimental error due to environmental effect associated with the ijklth. The variance component was calculated using the formula: σ2g=(MSgca-MSerror)/(p-2); σ2s=(MSsca-MSerrror)/1
Where, MSgca= variance due to GCA; MSsca= variance due to SCA; MSerror= error variance.
Gene action
Since the parents used in the crosses were considered fixed, coefficient of genetic determination were used to estimate total genetic variability, broad sense and narrow sense coefficient of genetic determination (CGD) by the formula below:
Baker’s ratio = (2 σ2gca)/( 2 σ2gca + σ2sca )
CGDBS= (2 σ2gca + σ2sca)/( 2 σ2gca + σ2sca + σ2e
CGDNS= (2 σ2gca)/( 2 σ2gca + σ2sca + σ2e)
Both Genstat 14th Edition (Payne et al., 2010) and Excel computer programmes were used to analyze the data.
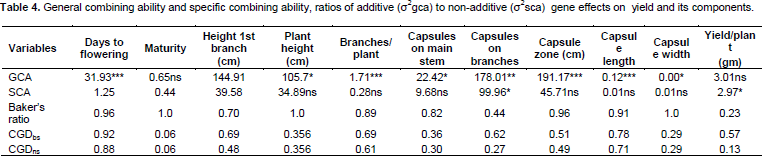
General combining ability
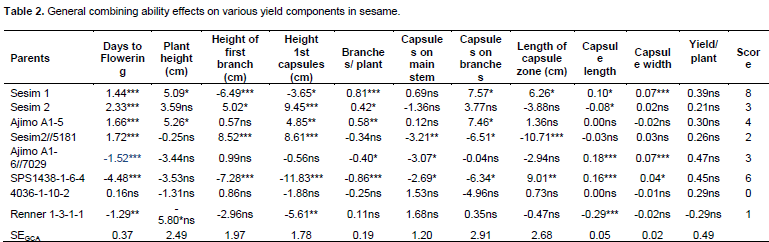
The general combining ability (GCA) effects of the parents are presented in Table 2. Sesim 1 had highly significant effect (P<0.001) for days to flowering, height to first branch, branches per plant and capsule width. It was also significant (P<0.05) for height to first capsule, plant height, capsules on branches, length of capsule zone and capsule length. This indicated that Sesim1 as a parent contributed much to the progenies and therefore is a desirable parent for crossing in order to improve those traits in a crossing programme. Sesim 2 was desirable for number of branches per plant such that it can only be used to increase the number of branches in the progeny. Ajimo A1-5 had desirable GCA effects on number of branches, plant height and capsules on the branches. It is second to Sesim 1 parent as it recorded significance to three desirable traits in its general combining ability. Sesim2//5181 and 4036-1-10-2 did not show any desirable trait as there was no significant level of difference in its contribution. They are therefore not desirable parents to be used in the crossing programme to improve yield and yield components but could be used for improving some other traits such as pest resistance. Ajimo A1-6//7029 had highly significant negative effect (P<0.001) on days to flowering and highly positive significant (P<0.001) effect on capsule length and width. Significant negative effect on days to flowering by Ajimo A1-6//7029 implies that it can be used as a parent to reduce days to flowering and maturity in progenies to adapt to effect of climate change.
Positive effect on capsule length and width would mean some increase in the number of seeds per capsule which indirectly contributes to yield. SPS1438-1-6-4 had highly negative significant (P<0.001) effect on days to flowering thus reducing flowering time and good to escape drought and other abiotic stresses. It had high significant (P<0.001) negative effect on height of first branch and first capsule and positive significant (P<0.5) effect on the length of the capsule fruiting zone. This is desirable since the number of capsules would be increased through low capsule setting and increased capsule length and thus increased yield. Renner 1-3-1-1 had high negative significant (P<0.01) effect on days to 50% flowering, height of first capsule, plant height and capsule length.
Sesim 1 was the best parent with the highest number of significant GCA effects followed by SPS1438-1-6-4. Thirugnana et al. (2006) reported significant results of general combining ability for all the characters in sesame for the experiment they evaluated under both normal and flood environments except 1000 seed weight under normal conditions. Significant results of GCA suggest the role of additive genetic effects in the inheritance of these characters. Ravindran and Raghinam (1996) reported predominance of additive gene effects on traits like branches per plant, days to 50% flowering, 1000 seed weight and height to first capsule. Early flowering could be a desirable selection criterion if the reproductive period was long enough to increase productivity or if the shorter time to flowering resulted in a concomitant decrease in time to maturity without decreasing the yield to a significant level or if it helps escape the terminal drought. Since none of the parents showed desirable combining ability for majority of the component characters, multiple crosses involving more than two parents would be appropriate techniques to be employed in the development of hybrids and or selection of superior recombinants in the segregating generation (Banerjee and Kole, 2009).
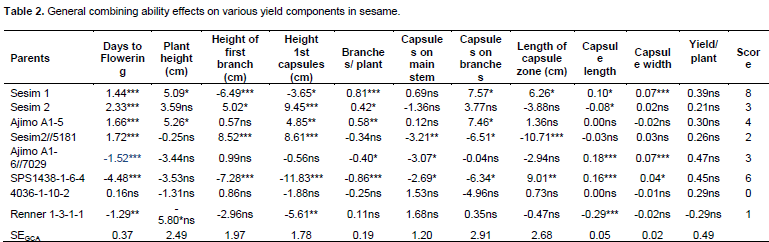
Specific combining ability
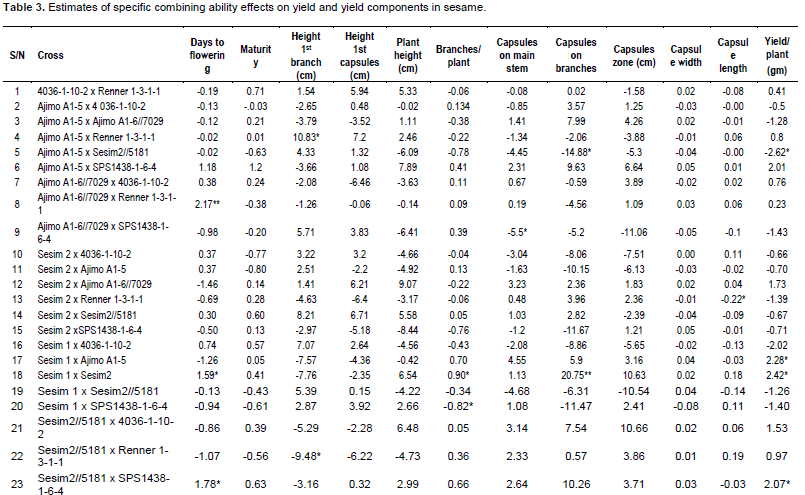
The results of specific combining ability (SCA) are presented in Table 3. Ajimo A1-5 x Renner 1-3-1-1 had positive significant (P<0.05) effect on the height of first branch (10.83) which is not desirable because few capsules would then be set on the main stem thus resulting in less yield per plant and more so if few or no branches are developed. Meanwhile, Sesim 2//5181 x Renner 1-3-1-1 had negative significant (P<0.05) SCA effect (-9.48) which is desirable as the capsules start developing from lower part of the main stem and therefore more capsules are formed on the plant. Sesim 1 x Sesim 2, Sesim1 x Ajimo A1-5 and Sesim 2//5181 x SPS 1438-1-6-4 showed positive significant (P<0.05) effect on yield per plant. Sesim 1 x Sesim 2 had positive significant (P<0.05) effect on days to flowering, number of branches per plant and number of capsules on branches. None of the hybrids exhibited superior SCA effects for all the characters indicating that there was no any cross that was superior for all the traits. Ravindran and Raghinam (1996) recorded seeds per capsule and capsule length to exhibit a preponderance of non-additive gene effects. Non-additive gene effects are non-fixable and have a consequence in slowing down selection progress but could be good for hybrid production. Yield per plant exhibited almost equal importance of both additive and non-additive gene effects. Thirugnana et al. (2006) recorded higher SCA variance than GCA variance for number of branches, number of capsules, number of seeds per capsule, 1000 seed weight and seed yield under normal conditions as compared to analysis under floods. They also reported that the magnitude of GCA variance was higher than that of SCA variance for days to 50% flowering and plant height under normal conditions, indicating the predominance of additive and additive x additive type of gene action. The SCA is considered to be the best criterion for selection of superior hybrids (Ahmed and Adam, 2014). From the results of this study, it could be concluded that both additive and non-additive gene action were important for improving seed yield in sesame.
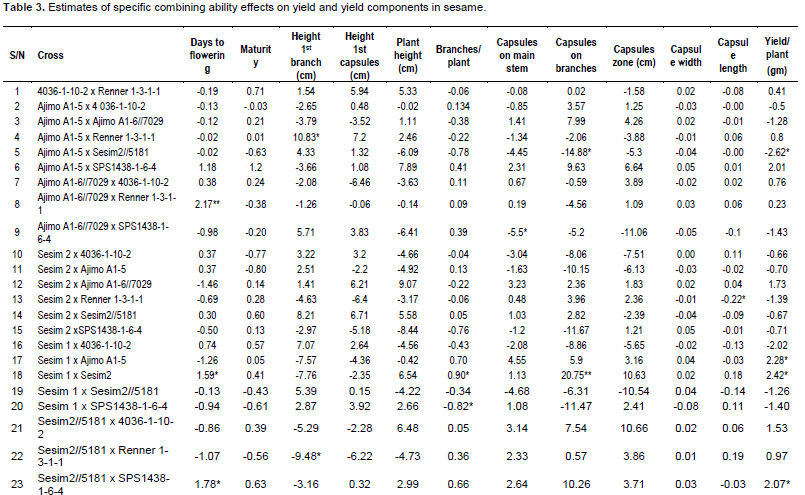
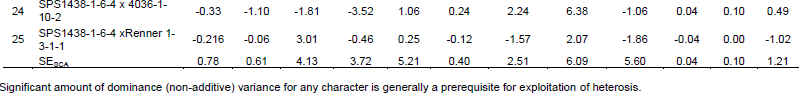
Effects of GCA and SCA were analyzed (Table 4). GCA effects were found to be highly significant (P<0.001) for days to flowering, branches per plant, length of capsule zone and significant (P<0.05) for plant height, capsules on main stem and capsule width, indicating preponderance of additive gene action. Praveenkumar et al. (2012) revealed that additive gene action was pre-dominant for plant height, number of secondary branches per plant, number of capsules per plant, 1000 seed weight and number of seeds per capsule. SCA showed significant effect on only capsules on branches and yield per plant, thus indicating major action of non-additive gene action for these traits. Murty (1975) reported that general combining ability variance was larger than specific combining ability variance for all the characters except of oil, indicating the predominance of additive gene action. Saravanan and Nadarajan (2003) reported that the GCA variance was greater than the SCA variance for eight traits including days to 50% flowering, plant height, number of primary branches per plant and phyllody incidence, indicating preponderance of additive gene action for those traits, while the SCA variance was greater than GCA variance for number of capsules per plant, number of seeds per capsule, single plant yield, indicating predominance of dominant gene action for these traits. Mothilal and Manoharan (2005) reported that non-additive gene action was involved in the expression of characters viz. number of capsules on branches, 1000 seed weight and seed yield per plant. They observed over dominance for number of branches, number of seeds per capsule and seed yield per plant. Thirugnana et al. (2006) reported the magnitude of GCA variance was higher than that of SCA variance for days to 50% flowering and plant height under normal conditions. Praveenkumar et al. (2012) also recorded that the proportion of GCA variances were higher as compared to SCA. Aladji et al. (2014) showed that the values of GCA/SCA ratios had SCA variance higher than GCA variance component except for number of seeds per capsule and days to maturity. They recorded that the SCA variance was more than GCA variance, indicating the role of non-additive gene action for the inheritance of date to flowering, duration of maturation, plant height, number of branches, number of capsules per plant and capsule length.
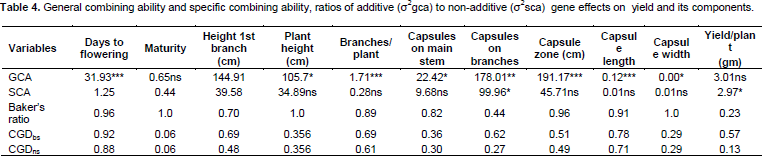
As recorded in Table 4, additive gene action controlled days to flowering, plant height, branches per plant, capsules on main stem, capsules on branches, length of capsule zone, capsule length and capsule width. Non-additive gene action controlled capsules on branches and yield per plant only. For selected traits controlled by non-additive gene action, it would therefore be desirable to maintain a certain degree of heterozygosity to exploit the additive gene effects and recurrent selection involving crossing desirable segregannts alternated with selection in order to increase the magnitude of additive genetic variance and at the same time to maintain heterozygosity.
Pedigree method is applied for selection of desirable traits that are controlled by additive gene action since they are fixable. Selection of progenies for these traits is done at early stages of segregation to produce pure lines. Meanwhile, traits controlled by non-additive (dominance and epistasis) gene action are good for hybrid production where dominance is important. The selection of these traits are normally delayed to later stages of segregation after some selfing in order to reduce the number of genes that heterozygous and genes would then be fixed to homozygosity.
Baker’s ratio was high for most of the traits (Table 4) except yield per plant (0.233) indicating that the parents contributed much to the progenies. Coefficient of genetic determination in broad sense (CGDbs) was higher for most traits except days to maturity, plant height, capsules on main stem and capsule width (0.29). Coefficient of genetic determination in narrow sense (CGDns) was high for most traits except days to maturity (0.06), capsules on main stem, capsules on branches, capsule width and yield per plant, implying that these traits have high heritability and the parents contributed highly to those traits. Where CGDns is low, it indicates non-additive (dominance or epistasis) behavior playing an important role. This also suggests that the progeny could not be predicted on the GCA values of the parents.
These results suggested that both additive and non-additive gene actions played a greater role in these traits. Sesim 1 had the highest number of desirable traits scored for GCA effects and could be considered as a parent for crossing to produce desirable progeny. Sesim 1 x Sesim 2, Sesim 1 x Ajimo A1-5 and Sesim2//5181 x SPS1438-1-6-4 recorded positive significant SCA effects for yield per plant and therefore they could be recommended for hybrid seed production.
The authors have not declared any conflict of interests.
REFERENCES
|
Aladji-Abatchoua MMI, Noubissie- Tchiangam JB, Njintang-Yanou N (2014). Genetic analysis of seed yield component in sesame (Sesamum indicum L.) at Mora (Cameroon). Sch. Acad. J. Biosci. 2(5):318-325.
|
|
|
|
Ahmed SBM, Adam SI (2014). Combining ability for yield and yield components in six parents and their 15 F1 hybrids of sesame (Sesamum indicum L.) in half diallel mating design. J. Plant Breed. Crop Sci. 6(12):179-184.
Crossref
|
|
|
|
|
Anyanga WO, Obong YY (2001). Sesame (Sesamum indicum L.) In: J.K. Mukiibi (ed.) Agriculture in Uganda. Crops 11:97-102.
|
|
|
|
|
Ashri A (1989). Sesame breeding objectives and approaches. In: Oil Crops: Sunflower, Linseed and Sesame, Ed. Omran, A., IDRC-MR205E, IDRC, Ottawa.
|
|
|
|
|
Auckland AK (1970). Sesame Improvement in East Africa. In: Crop Improvement in East Africa, Chapter Ed. C.L.A. Leakey. 6:157-177.
|
|
|
|
|
Baker RJ (1978). Issues in diallel analysis. Crop Sci. 18(4):533-536.
Crossref
|
|
|
|
|
Banerjee PP, Kole PC (2009). Combining ability analysis for seed yield and some of its component characters in sesame (Sesamum indicum L.). Int. J. Plant Breed. Genet. 3(1):11-21.
Crossref
|
|
|
|
|
Bedigian D, Seigher DS, Harlan JR (1985). Sesamin, sesamolin and the origin of sesame. Biochem. Syst. Ecol. 13(2):133-139.
Crossref
|
|
|
|
|
Brigham RD (1985). Status of sesame research and production in Texas and the USA. Sesame and safflower: status and potentials. FAO Plant Prod. Prot. 66:73-74.
|
|
|
|
|
Coulman KD, Liu Z, Hum WQ, Michaelides J, Thompson LU (2005). Whole sesame is as rich a source of mammalian lignin precursors as whole flaxseed. Nutr. Cancer 52:156-165.
Crossref
|
|
|
|
|
FAOSTAT (2009). Food and Agricultural Organization of the United Nations.
|
|
|
|
|
FAOSTAT (2015). Food and Agricultural Organization of the United Nations.
|
|
|
|
|
Griffings B (1956). Concept of general and specific combining ability in relation to diallel crossing system. Austr. J. Biol. Sci. 9:463-493.
Crossref
|
|
|
|
|
Johnson LA, Suleiman TM, Lusas EW (1979). Sesame protein, a review and prospectus. J. Am. Oil Chem. Soc. 56(3):463-468.
Crossref
|
|
|
|
|
Moazzami AA, Kamal-Eldin A (2006). Sesame seed is arich source of dietary lignans. J. Am. Oil Chem. Soc. 8:719-723.
Crossref
|
|
|
|
|
Mothilal C, Manoharan V (2005). Diallel analysis for the estimation of genetic parameters in Sesame (Sesamum indicum L.). Agric. Sci. Digest 25(2):133-135.
|
|
|
|
|
Muhammad SH, Sedeck FS (2015). Combining ability and heterosis estimates in sesame. World Appl. Sci. J. 33(5):690-698.
|
|
|
|
|
Murty DS (1975). Hetrosis, combining ability and reciprocal effects for agronomic and chemical characters in sesame. Theor. Appl. Genet. 45:294-299.
Crossref
|
|
|
|
|
Payne RW, Harding SH, Murray DA, Soutar DM, Baird DB, Glaser AR, Thompson R, Webster R (2010). The Guide to Genstat Release 14, Part 2: Statistics. Hemel Hempstead: VSN International, U.K.
|
|
|
|
|
Praveenkumar KM, Nadaj HL, Patil RK and Deshpande SK (2012). Combining ability and gene action studies in inter-mutant hybrids of sesame (Sesamum indicum L.). Karnataka J. Agric. Sci. 25(1):1-4.
|
|
|
|
|
Ravindran RG, Raghinam AD (1996). Combining ability analysis on sesame. Sesame and Safflower Newslett. 11:70-75.
|
|
|
|
|
Saravanan S, Nadarajan N (2003). Combining ability studies in sesame. Sesame and Safflower Newslett. 18:20-24.
|
|
|
|
|
Singh RK, Chaudhary BD (1985). Biometrical Methods in Quantitative Genetic Analysis. Kalyani Publishers, New Delhi, P 300.
|
|
|
|
|
Solanki ZS, Gupta D (2003). Inheritance studies for seed yield in sesame. Sesame and Safflower Newslett. 18:25-28.
|
|
|
|
|
Thirugnana-Kuma S, Srinivasan R, Kubenthiran K, Eswaran R, Kumar CPS (2006). Studies on combining ability and heterosis for yield and yield component characters under normal and temporarily flooded conditions in sesame (Sesamum indicum L.). Sesame and Safflower Newslett. 21.
|
|