Full Length Research Paper

ABSTRACT
Stay green is the plant’s ability to sustain photosynthetically functional green leaf area for longer periods, even with excessive water stress. This study was done to understand the gene action that controlled inheritance of the stay green trait with the general objective of contributing to breeding more drought tolerant maize varieties. 10 F1 hybrids from crosses between highly performing and the least performing drought tolerant inbred lines as regards selected stay green physiological determinants were planted under a rain out shelter with a well-watered control and excessive water-stress treatments applied from six weeks after planting and eight weeks after planting. With progress in days after flowering, data were collected on relative chlorophyll per unit leaf area (RC: LAUG), maximum quantum yield of photosystem II photochemistry (ΦII), non-photochemical quenching (NPQ) and linear electron flow (LEF). RC: LAUG and NPQ were mainly inherited by additive gene action. ΦII was controlled more by the environment than interallelic interactions while LEF inheritance was controlled mainly by interallelic interactions.
Key words: Additive, inheritance, non-additive, photosynthesis, stay green.
INTRODUCTION
Stay green is the ability of a plant to maintain functional photosynthetic leaf area for longer periods after physiological maturity, even under excessive shortage of water (Luo et al., 2006). Due to their high photosynthetic activity beyond physiological maturity, such plants have a higher deposition of assimilates in their sinks, increased resistance to diseases and pests, along with the pronounced tolerance to water stress (Thomas and Smart, 1993). Breeding programs targeting water stress prone ecologies make use of secondary traits such as stay green when selecting and advancing populations that are suitable for these limiting environments (Edmeades et al., 1997). Such programs make use of knowledge on gene actions controlling the inheritance of the traits to effectively design breeding approaches that can maximize the breeding value of selected germplasm.
A number of annual crop species inclusive of maize exhibit genetic variation for the degree of retention of leaf pigmentation under water stress (Thomas and Smart, 1993). However, little has been established about the inheritance of stay green ability in maize, which is a prerequisite for the successful improvement of cultivars for the trait (Belicuas et al., 2014). In other crops such as sorghum, green leaf area retention, was reported to be regulated by both dominant and recessive epistatic interactions (Tenkouano et al., 1993), with the level of dominant gene action dependent on the environment (Walulu et al., 1994).
The few studies on stay green inheritance in maize such as that by Belicuas et al. (2014), reported that both the additive and dominance variances for the trait were significant, with the additive variance exceeding the dominance variance by a factor of 6.41 therefore classifying the average level of dominance as partial dominance. Indeed, other studies such as those by Guei and Wasson (1996), Banziger et al. (2000), Lee et al. (2005) and Costa et al. (2008) also reported that inheritance of the stay green trait was controlled mostly by additive effects than non-additive effects. Joshi et al. (2007) also reported that the trait had medium to high heritability with strong contribution of additive effects. Traits controlled largely by additive effects have the advantage of being easily improved for in populations using conventional approaches, which are cheaper, and have established legal systems in the developing world (La Rovere et al., 2010).
Majority of the previous studies on inheritance of the stay green trait in maize, however, made use of visual scoring while assessing genotypes for stay green (Belicuas et al., 2014). The need for less biased data requires less individual-centered methods for assessing for the trait as raised by Borrell et al. (2008). Chlorophyll fluorescence-based methods have since been developed to overcome this problem such as those provided by the Minolta chlorophyll meter SPAD-502 (Inoue et al., 1988) and more recently, photosynQ (Kuhlgert et al., 2016). These approaches, alongside providing un-biased information on visual leaf pigmentation, provide in-depth information about how physiologically functional the pigmentation is.
This is by assessing for rates of photosynthesis-related physiological processes such as non-photochemical quenching and linear electron flow under various environments, that way providing more useful information to the breeding programs during selection of germplasm. The parameters measured for by these chlorophyll fluorescence approaches have, however, not been studied for the gene actions controlling their inheritance, especially in tropical maize germplasm.
The objective of this study, therefore, was to explore the gene action that controlled the individual modes of inheritance of these parameters, in order to provide a better understanding of their heterosis, and of the contribution of individual parental lines to functional stay green expression in maize hybrids.
MATERIALS AND METHODS
Site of the study
The study was carried out at the National Crops Resources Research Institute (NaCRRI), Namulonge in Uganda, under a rain-out shelter. Namulonge is located in the Lake Victoria crescent area at 0.5288° N, 32.6123° E, at an altitude of 1204 m (Otim et al., 2018). The area had an average temperature of 21.1°C, with mean daily maximum and minimum temperatures of 27 and 17°C, respectively during the period of study (Uganda National Meteorological Authority, 2020).
Constitution of F1 hybrids
Five high performing inbred lines (CELQ15027, CELQ15034, CELQ15031, CELQ15018, CELQ15046), were planted and mated with two low performing inbred lines (CELQ15043, CELQ15045), in regard to stay green physiological determinants as assessed by Epaku et al. (2021). These inbred lines were sourced from the International Maize and Wheat Improvement Center (CIMMYT) breeding programs in Kenya and Ethiopia. A North-Carolina II mating design was used producing 10 F1 hybrids. This was done under field conditions with each entry having one row constituting eight hills spaced at 75 × 30 cm. Three seeds were planted per hill and later thinned to one. N.P.K fertilizer (17:17:17) was applied at a rate of 12 g per hill twice; first at planting and when the plants were 65 cm high by side dressing.
Evaluation of resultant F1 hybrids
The resultant F1 hybrids were evaluated using a completely randomized design, with two replications and four plants for each hybrid under each replicate. The experiment comprised a well-watered (WW) treatment and two water-stress treatments that were applied starting at six weeks after planting (6WAP) and at eight weeks after planting (8WAP). The potting mixture for the WW treatment was maintained at a water potential between 0.003-0.006 MPa; while that for the water-stress treatments was maintained at a water potential between 0.015 and 0.02 MPa using a tensiometer (Zaidi et al., 2015). The procedure involved measuring the soil moisture at 10, 20 and 30 cm three times every seven days to determine the amount of water to be added for maintenance of the required water potential. N.P.K fertilizer (17:17:17) was applied at a rate of 12 g per pot twice; first at planting and when the plants were 65 cm high, by side dressing within the pot. The photosynthetically active radiation under the rain-out shelter was 450 to 570 nm supplied by sunlight with a 12 h light period. Ambient temperatures in the shelter were 27 to 32°C and 24 to 56% relative atmospheric humidity as measured with a multi-spectrophotometer (Kuhlgert et al., 2016).
Data collection
Data were collected on maximum quantum yield of photosystem II photochemistry (ΦII), linear electron flow (LEF), non-photochemical quenching (NPQ) and relative chlorophyll (RC) using a multi-spectrophotometer operating on photosynq version v.10 (Kuhlgert et al., 2016). This was done at the base of the leaf lamina of the second and fourth leaves from the top of the plant, for three plants per entry, and their respective averages computed. For leaf area under green (LAUG), one sided green leaf area of each genotype was measured by multiplying the length with the widest width of the longest green leaf per genotype, followed by dividing this result with the ground area occupied by each genotype (Wang et al., 2012). The ratio of RC to LAUG was then computed to obtain RC: LAUG. Data for all parameters were collected first at six weeks after emergence of the crop before commencement of the water stress treatments. NPQ was assessed again at 60 days after flowering (DAF) under all treatments; RC: LAUG was assessed again at 20, 40 and 50DAF under WW, 8WAP and 6WAP treatments, respectively; ΦII was assessed again at 40DAF under all treatments; while LEF was assessed again at 20, 40 and 50DAF under WW, 8WAP and 6WAP treatments, respectively as suggested by Epaku et al. (2021).
Data processing and analysis
Data collected were analyzed using the linear model:
Yijk = μ + Mi + Fj + MFij + MEie + FEje + FMEije + Ee + R(E)kl + εijk (Comstock and Robinson, 1948).
where Yijk = value of F1 cross of the ith female and the jth male in the kth replication as regards stay green; μ = population mean; i = 1, 2, …, n (n = 5 males); j=1,2 (females); k = 1 to 2 replications; l = 1, 2,..b (b = 4 plants per entry); Mi = effect of the ith male; Fj = effect of the jth female; MFij = effect of interaction between ith male and jth female; Rk = effect of the kth replication; eijk = error effect of the ijkth observation.
General combining abilities (GCAi) were computed as the difference between the general performance of a line across all crosses (yi) and the population mean (y); (GCAi = yi- y). The significance of the GCA effects were tested using a t-test where t = GCA/SEGCA. Specific combining ability (SCAij) was computed as (SCAij = GCAij – gi – gj); where gi = GCA of the ith female; gj = GCA of jth male.
Heritability was estimated in form of broad-sense coefficient of genetic determination (BS.CGD) which is the total genetic variation, and narrow- sense coefficient of genetic determination (NS.CGD) which is the proportion which can be explained by additive effects (Dabholkar, 1992).
Heritability estimates across treatments were calculated as:
BS.CGD = ((2* σ2gca) + σ2sca) / ((2* σ2gca) + σ2sca+ ((σ2gca × site + σ2sca × site)/3) + (error/6))
NS.CGD = (2* σ2gca) / ((2* σ2gca) + σ2sca+ ((σ2gca × site + σ2sca × site)/3) + (error/6))
Heritability estimates within environments were calculated as:
BS.CGD= ((2* σ2gca) + σ2sca) / ((2* σ2gca) + σ2sca + error)
NS.CGD= (2* σ2gca)/ ((2* σ2gca) + σ2sca+ error)
Baker’s ratio (BR) was calculated using variance components due to GCA and SCA as:
BR= (2*σ2gca) / ((2*σ2gca) + σ2sca)
RESULTS
The results of the ANOVA for all the parameters are presented in Table 1. There was no significant variation for ΦII and RC: LAUG among genotypes under all treatments. LEF significantly varied among males, females and male*female interactions (P≤0.05) under the 6WAP treatment while under the 8WAP treatment it varied between females and male*female interactions (P≤0.05). NPQ showed significant variation (P<0.001) among males under the 6WAP and 8WAP treatments.
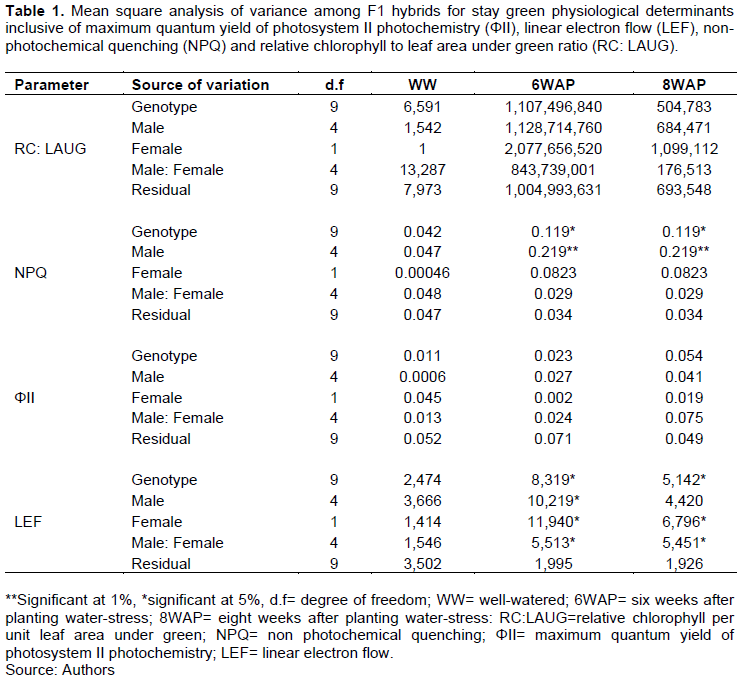
The results of the combining ability analysis and variance components for RC: LAUG are shown in Table 2. All the genotypes had insignificant GCA for RC: LAUG under all treatments. The cross CELQ15045× CELQ15046 had positive significant SCA (P≤0.05) for RC: LAUG under the 8WAP treatment. The trait generally had a low heritability across treatments but with a higher heritability under the well-watered treatment than in the water stress treatments.
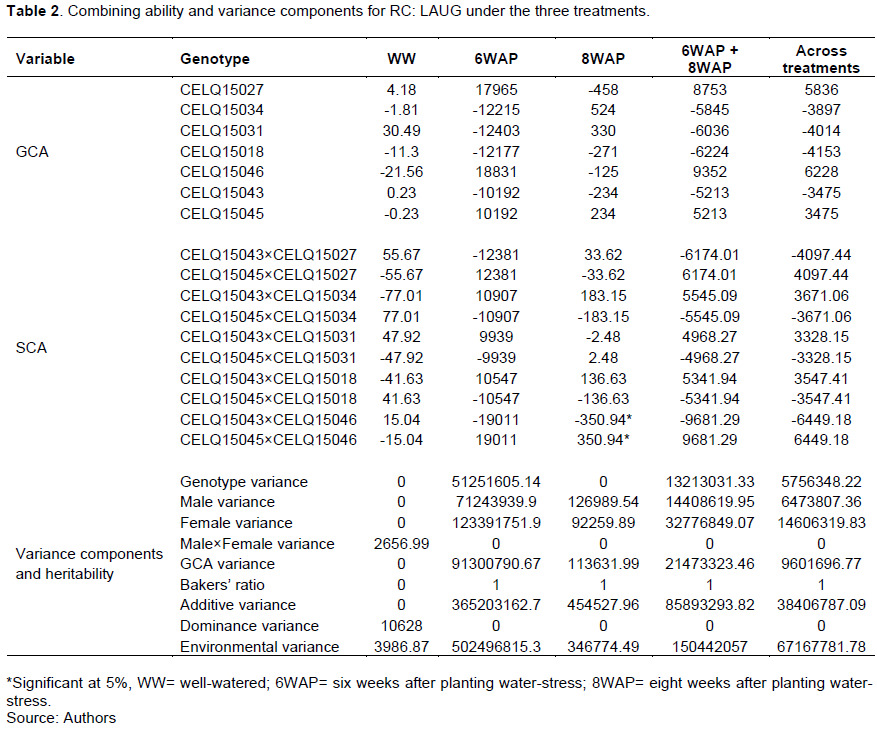
The results of the combining ability analysis and variance components for NPQ are shown in Table 3. All the genotypes had insignificant GCA and SCA for NPQ under all treatments. GCA for NPQ was higher under the water-stress treatments. CELQ15018 and CELQ15043 showed positive GCA in all the treatments. CELQ15018 and CELQ15043 had positive GCA for NPQ under all treatments. Heritability for NPQ was higher under the water stress treatments than in the WW treatment.
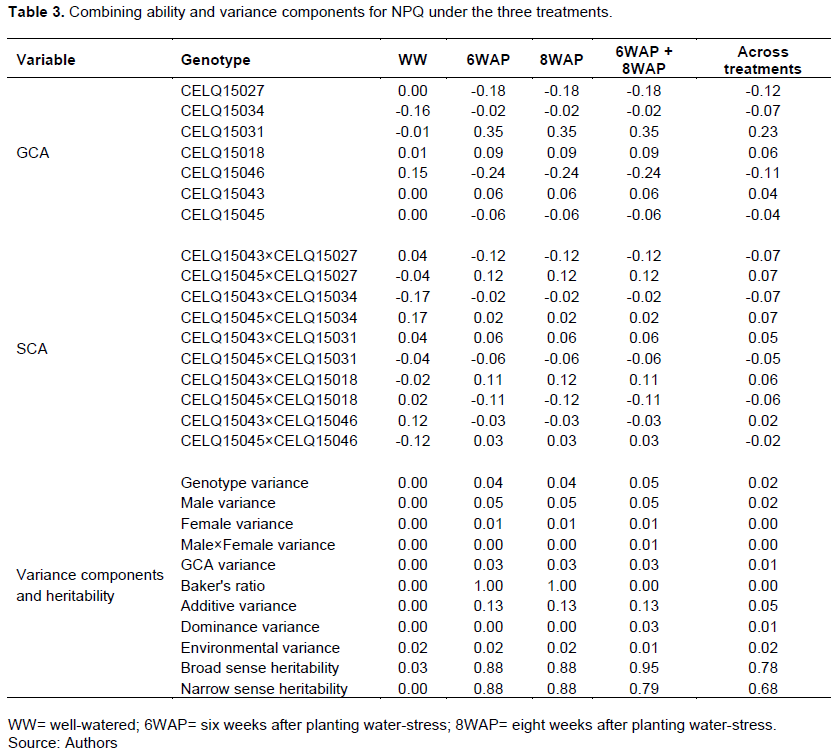
The results of combining ability analysis and variance components for ΦII are shown in Table 4. All the genotypes had insignificant GCA and SCA for ΦII under all treatments. GCA for ΦII was generally higher under the WW treatment. CELQ15031, CELQ15046 and CELQ15045 had positive GCA for ΦII under all treatments. Cross CELQ15045×CELQ15027 showed the highest positive SCA for ΦII under all treatments. The trait had no detectable narrow sense heritability under all treatments but a moderate broad sense heritability under the water-stress treatments.
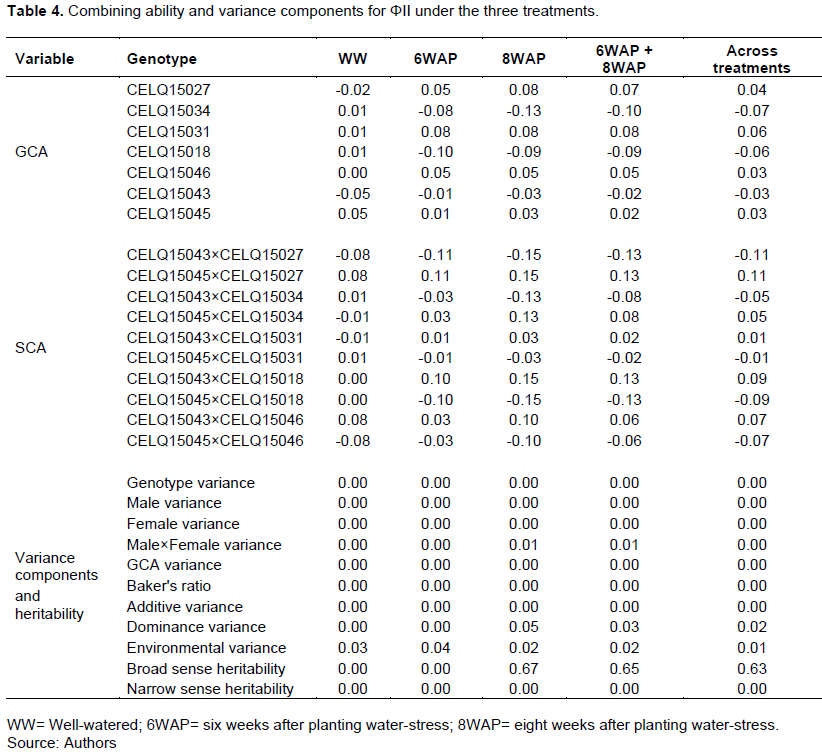
The results of combining ability analysis and variance components for LEF are shown in Table 5. All the genotypes had insignificant GCA for LEF under all treatments. GCA and SCA for LEF were generally higher under the water stress treatments than in the WW treatment. CELQ15031 had the highest positive GCA in all the environments.
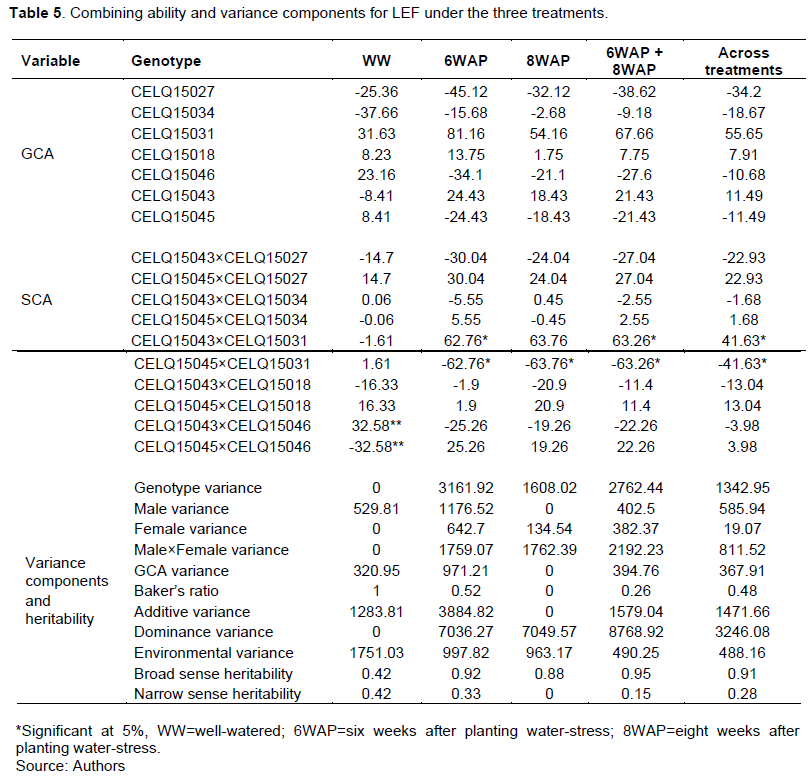
Crosses CELQ15045×CELQ15046, CELQ15043× CELQ15031 and CELQ15045×CELQ15031 had positive significant SCA (P≤0.05) for LEF under the WW, 6WAP and 8WAP treatments, respectively. The trait also had a higher broad sense heritability especially under the water stress treatments.
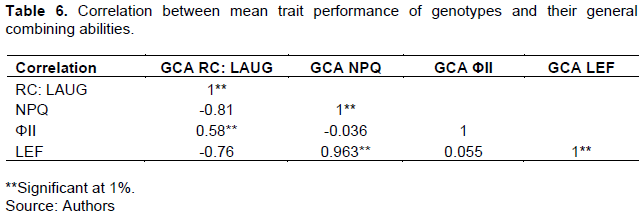
The results for the correlation studies between mean performance of genotypes for particular traits and their GCA for those respective traits are shown in Table 6. Mean ΦII had a significant (P<0.001) positive correlation with GCA for RC: LAUG. Mean LEF had a significant (P≤0.05) positive correlation with GCA for NPQ. Mean LEF had a positive insignificant correlation with GCA for ΦII. The means of all parameters apart from ΦII had a positive significant (P<0.001) correlation with the GCAs of the genotypes for those respective traits.
DISCUSSION
Maximum quantum yield of photosystem II photochemistry (ΦII) inheritance was predominantly under the control of the environment with a small percentage controlled by inter-allelic interactions. This suggested that where-as heterosis could be exploited to improve germplasm for this trait as reported by Zhang et al. (2000), a great deal of the performance of the resultant genotypes had to do with how favorable the environment they were exposed to was in terms of photosynthetic efficiency. Malik et al. (1999) however, reported that inheritance for net-photosynthesis was mainly under the control of additive gene effects, probably owing to the different methods of measurement for the trait used in their study. The significant positive correlation between mean ΦII and GCA for RC: LAUG suggested that these two traits could be selected for simultaneously to hasten the breeding process (Banziger et al., 2000).
Generally, the variation for NPQ and LEF among the F1 hybrids was more under the 6WAP treatment followed by the 8WAP typically owing to the relative length of periods of exposure to water-stress (Efeo?lu et al., 2009). The variation for NPQ only under the water-stress treatments implied that the trait could best be selected under this treatment. Additionally, variation for this trait was mainly due to additive effects implying that it could effectively be improved by re-current selection among germplasm (Badu-Apraku et al., 2011). Selection for high mean NPQ among lines could imply selection for lines with a high GCA for LEF (Banziger et al., 2000). NPQ had a moderate heritability across environments with higher heritability under the water-stress treatments affirming the need to select for it under water-stress treatments (Badu-Apraku et al., 2013). Crosses CEL15043×CEL15031, CEL15045×CEL15034 and CEL15045×CEL15046 showed positive though insignificant SCA for this trait suggesting that these crosses could be studied in different environments to further understand and exploit their heterosis (Bekavac et al., 2007). This heterosis suggested that where-as inheritance for NPQ was mainly additive, a significant percentage of it was under the control of inter-allelic interactions. Similar findings were reported by Holá et al. (2010) that non-additive gene effects were just as important as additive gene effects in the inheritance of photosynthetic parameters.
The F1 hybrids showed significant variation for LEF only under the water stress treatments, with the highest heritability recorded under the same suggesting that inheritance for this trait would be better studied under similar environmental conditions. Heritability for this trait was largely due to dominance variance implying that it was mainly under the control of inter-allelic interactions and would be better exploited through heterosis rather than re-current selection (Rojas and Sprague, 1952). Particularly, the cross CELQ15043XCELQ15031 showed positive significant SCA for this trait under the water stress treatments suggesting that it was the best performing for this trait among the germplasm screened (Cairns et al., 2013). Crosses inclusive of CELQ15045XCELQ15027, CELQ15045XCELQ15046 and CELQ15045XCELQ15034 also showed positive though insignificant SCAs for this trait suggesting the need to screen them in even more environments for further exploration of their heterosis (Cukadar?Olmedo and Miller, 1997). Inbred lines CELQ15031, CELQ15018 and CELQ15043 also showed positive though insignificant GCA for LEF implying the need to cross them in even more diverse populations to further exploit their genetic ability to improve germplasm for this trait (Veeresha et al., 2015).
RC: LAUG was moderately under the control of additive effects under the water-stress treatments used in this study, a finding that was similar to those by Cukador-Olmedo and Miller (1997), Joshi et al. (2007),Belicuas et al. (2014), Sayed and Bedawy (2016), and Annor and Badu-Apraku (2016) who reported that retention of leaf pigmentation was mainly under the control of additive gene effects. The significant SCAs for RC: LAUG in crosses CELQ15043XCELQ15046 and CELQ15045XCELQ15046 were in agreement with the findings by Mhike et al. (2012) and suggested that these crosses could be studied further for heterosis in regards to this trait. The high heritability for this trait under the well-watered treatment was also in agreement to the findings by Joshi et al. (2007) who reported heritability for leaf pigmentation between 0.73 and 0.80; but disagreed with the findings by Mhike et al. (2012), who reported that leaf pigmentation retention had a higher heritability under water-stress conditions, leaving the conflicting results un-resolved.
The higher GCAs in water-stress treatments for all traits were an indication that these inbred lines had a higher breeding value for these traits under water-stress (Jiang et al., 2004). Broad sense heritability estimates for ΦII, LEF and NPQ were higher than 0.5 suggesting the need for further selection for each trait (Sayed and Bedawy, 2016).
CONCLUSION
The study therefore, established that non-additive gene effects were just as important as additive gene effects as regards inheritance of the physiological determinants for the stay green trait, and that the effect the environment had on the inheritance of these traits should be taken into account. The crosses CELQ15045XCELQ15046 and CELQ15043XCELQ15031 exhibited positive and highly significant SCA effects and could be considered as good specific combiners for leaf pigmentation (RC: LAUG) and LEF across treatments.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENT
This work was accomplished with financial support from the Regional Universities Forum for Capacity in Agriculture (RUFORUM) together with the World Bank through the African Centre of Excellence in Materials, Product Development and Nanotechnology (MAPRONANO ACE II).
REFERENCES
|
Annor B, Badu-Apraku B (2016). Gene action controlling grain yield and other agronomic traits of extra-early quality protein maize under stress and non-stress conditions. Euphytica 212(2):213-228. |
|
|
Badu-Apraku B, Oyekunle M, Akinwale RO, Lum AF (2011). Combining ability of early-maturing white maize inbreds under stress and nonstress environments. Agronomy Journal 103(2):544-557. |
|
|
Badu-Apraku B, Oyekunle M, Fakorede MAB, Vroh I, Akinwale RO, Aderounmu M (2013). Combining ability, heterotic patterns and genetic diversity of extra-early yellow inbreds under contrasting environments. Euphytica 192(3):413-433. |
|
|
Banziger M, Edmeades GO, Beck D, Bellon M (2000). Breeding for drought and nitrogen stress tolerance in maize: From theory to practice. CIMMYT, Mexico, DF 68 p. |
|
|
Bekavac G, Purar B, Stojakovi? M, Jockovi? D, Ivanovi? M, Nastasi? A (2007). Genetic analysis of stay green trait in broad-based maize populations. Cereal Research Communications 35(1):31-41. |
|
|
Belicuas PR, Aguiar AM, Bento DAV, Câmara TMM, de Souza Junior CL (2014). Inheritance of the stay green trait in tropical maize. Euphytica 198(2):163-173. |
|
|
Borrell AK, Jordan D, Mullet J, Klein P, Klein R, Nguyen H, Henzell B (2008). Discovering stay green drought tolerance genes in sorghum: a multidisciplinary approach. In: 14th Australian Agronomy Conference pp. 21-25. |
|
|
Cairns JE, Crossa J, Zaidi PH, Grudloyma P, Sanchez C, Araus JL, Menkir A (2013). Identification of drought, heat, and combined drought and heat tolerant donors in maize. Crop Science 53(4):1335-1346. |
|
|
Comstock RE, Robinson HF (1948). The components of genetic variance in populations of biparental progenies and their use in estimating the average degree of dominance. Biometrics 4(4):254-266. |
|
|
Costa EF, Santos MF, Moro GV, Alves GF, de Souza Junior CL (2008). Inheritance of the delayed senescence in maize. Pesquisa Agropecuária Brasileira 43(2):207-213. |
|
|
Cukadar?Olmedo B, Miller JF (1997). Inheritance of the stay green trait in sunflower. Crop Science 37(1):150-153. |
|
|
Dabholkar RR (1992). Elements of biometrical genetics. Ashok, Kumar. |
|
|
Edmeades GO, Bolanos J, Chapman SC (1997). Value of secondary traits in selecting for drought tolerance in tropical maize. In. Edmeades GO, Banziger M, Mickelson HR, Pena-Valdivia CB (Eds.), Developing drought -and Low N-Tolerant maize. Proceedings of a symposium, March 25-29, 1996, CIMMYT, El Batan, Mexico pp. 222-234. |
|
|
Efeo?lu B, Ekmekçi YA, Çiçek NU (2009). Physiological responses of three maize cultivars to drought stress and recovery. South African Journal of Botany 75(1):34-42. |
|
|
Epaku GT, Rubaihayo P, Kagoda F, Bomet KD, Badji A, Chapu I, Ogwal G (2021). Stay green physiological capacity of drought tolerant maize inbred lines. African Crop Science Journal 29(2):309-323. |
|
|
Guei RG, Wassom CE (1996). Genetic analysis of tassel size and leaf senescence and their relationships with yield in two tropical lowland maize populations. African Crop Science Journal 4(3):275-281. |
|
|
Holá D, Benešová M, Honnerová J, Hnili?ka F, Rothová O, Ko?ová M, Hnili?ková H (2010). The evaluation of photosynthetic parameters in maize inbred lines subjected to water deficiency: Can these parameters be used for the prediction of performance of hybrid progeny? Photosynthetica 48(4):545-558. |
|
|
Inoue M, Taniguchi N, Hata Y, Hoda T, Kudo Y, Ueda H (1988). U.S. Patent No. 4,728,978. Washington, DC: U.S. Patent and Trademark Office. |
|
|
Jiang GH, He YQ, Xu CG, Li XH, Zhang Q (2004). The genetic basis of stay green in rice analyzed in a population of doubled haploid lines derived from an indica by japonica cross. Theoretical and Applied Genetics 108(4):688-698. |
|
|
Joshi AK, Kumari M, Singh VP, Reddy CM, Kumar S, Rane J, Chand R (2007). Stay green trait: variation, inheritance and its association with spot blotch resistance in spring wheat (Triticum aestivum L.). Euphytica 153(1-2):59-71. |
|
|
Kuhlgert S, Austic G, Zegarac R, Osei-Bonsu I, Hoh D, Chilvers MI, Roth MG, Bi K, TerAvest D, Weebadde P, Kramer DM (2016). MultispeQ Beta: a tool for large-scale plant phenotyping connected to the open PhotosynQ network. Royal Society open science 3(10):160592. |
|
|
La Rovere R, Abdoulaye T, Kostandini G, Guo Z, Mwangi W, MacRobert J, Dixon J (2010). Economic, production, and poverty impacts of investing in maize tolerant to drought in Africa: an ex-ante assessment. The Journal of Developing Areas 48(1):199-225. |
|
|
Lee EA, Ahmadzadeh A, Tollenaar M (2005). Quantitative genetic analysis of the physiological processes underlying maize grain yield. Crop science 45(3):981-987. |
|
|
Luo PG, Deng KJ, Hu XY, Li LQ, Li X, Chen JB, Tan FQ (2006). Chloroplast ultrastructure regeneration with protection of photosystem II is responsible for the functional 'stay?green 'trait in wheat. Plant, Cell and Environment 36(3):683-696. |
|
|
Malik TA, Wright D, Virk DS (1999). Inheritance of net photosynthesis and transpiration efficiency in spring wheat, Triticum aestivum L., under drought. Plant Breeding 118(1):93-95. |
|
|
Mhike X, Okori P, Magorokosho C, Ndlela T (2012). Validation of the use of secondary traits and selection indices for drought tolerance in tropical maize (Zea mays L.). African Journal of Plant Science 6(2):96-102. |
|
|
Otim MH, Tay WT, Walsh TK, Kanyesigye D, Adumo S, Abongosi J, Asea G (2018). Detection of sister-species in invasive populations of the fall armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae) from Uganda. Plos One 13(4):0194571. |
|
|
Rojas BA, Sprague OF (1952). A Comparison of Variance Components in Corn Yield Trials: III. General and Specific Combining Ability and Their Interaction with Locations and Years 1. Agronomy Journal 44(9):462-466. |
|
|
Sayed MA, Bedawy IM (2016). Heterosis and inheritance of some physiological criteria imparting drought tolerance of grain sorghum in the irrigated and water-limited environments. Egyptian Journal of Agronomy 38(2):293-318. |
|
|
Tenkouano A, Miller FR, Frederiksen RA, Rosenow DT (1993). Genetics of nonsenescence and charcoal rot resistance in sorghum. Theoretical and Applied Genetics 85(5):644-648. |
|
|
Thomas H, Smart CM (1993). Crops that stay green 1. Annals of applied Biology 123(1):193-219. |
|
|
Uganda National Meteorological Authority (2020). |
|
|
Veeresha BA, Hanamaratti NG, Salimath PM (2015). Heterosis and combining ability studies for yield and productivity traits in rice. International Journal of Current Agricultural Research 4(5):120-126. |
|
|
Walulu RS, Rosenow DT, Wester DB, Nguyen HT (1994). Inheritance of stay green trait in sorghum. Crop Science 34:970-972. |
|
|
Wang AY, Li Y, Zhang CQ (2012). QTL mapping for stay green in maize (Zea mays). Canadian Journal of Plant Science 92(2):249-256. |
|
|
Zaidi PH, Rashid Z, Vinayan MT, Almeida GD, Phagna RK, Babu R (2015). QTL mapping of agronomic waterlogging tolerance using recombinant inbred lines derived from tropical maize (Zea mays L.) germplasm. Plos One 10(4):e0124350. |
|
|
Zhang MQ, Chen RK, Luo J, Lu JL, Xu JS (2000). Analyses for inheritance and combining ability of photochemical activities measured by chlorophyll fluorescence in the segregating generation of sugarcane. Field Crops Research 65(1):31-39. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


