Full Length Research Paper

ABSTRACT
A study was conducted to determine the mode of inheritance for grain shape and aroma traits of lines in the Uganda rice germplasm. Five aromatic and four non-aromatic rice lines were crossed in screen house using North Carolina Design II at the National Crops Resources Research Institute, Uganda. The resulting F2 seeds and their parents were planted in the field using complete randomized block design in two replications. The grain shape and aroma intensity of the harvested F3 seeds on per hill basis were determined in laboratory using instrumental and sensory evaluation procedures, respectively. The results revealed that the inheritance of grain shape and aroma traits were influenced by both additive and non-additive gene actions, but with the grain shape being dominated by additive gene action and the aroma by non-additive gene action. In addition, Supa 3, Supa 5 and Supa 1052 rice lines exhibited good general combining ability effects for the shape trait while Supa 3 and MET 13 were good general combiners for the aroma trait. The superior cross combination from Supa 3 x Supa 5 for grain shape and Supa 3 x MET 13 for aroma trait could be used for selection of the relevant traits at advanced stages of the breeding cycles.
Key words: General combining ability effects, gene action, North Carolina design II, selection, sensory evaluation.
INTRODUCTION
Rice (Oryza sativa L.) is an important food crop that has grown to be a significant cash crop in Uganda (Kijima and Otsuka, 2017) and in particular, the more preferred aromatic rice (Mogga et al., 2018) reported to offer supplementary health benefits (Prodhan et al., 2017). The utilization of the crop has also broadened in different industries such as food manufacturing (Plengsaengsri, 2019), health (Sen et al., 2020), energy (Olupot et al., 2015; Menya et al., 2018) and the construction sector (Yiga et al., 2019). In regards to the food sector, rice is “Not Just a Grain’’ (Fitzgerald et al., 2008) as consumers tend to make choices based on the rice grain qualities (Candia, 2013) and with the continuous strengthening of the economies of rice producing countries, demand for superior rice grain quality also increases. The most important rice grain traits in Uganda include grain length, shape, colour, stickiness when cooked, aroma and taste (Calingacion et al., 2015). Although rice grain qualities differ around the world, long and slender shape of rice grains is preferred in Africa (Asante, 2017). Rice aroma, characterized by the sweet popcorn smell (Buttery, 1983), in both raw and cooked grains is the most important trait (Mogga et al., 2018). In addition, the rice grain traits such as grain shape defined by the ratio of grain length to the grain width (Singh et al., 2000) is considered important (Kitara et al., 2018) with preference mostly for slender grain shape. The grain shape gives the first impression of the grain quality thus influencing the choice of the rice consumers (Calingacion et al., 2015).
Consequently, the improvement of the grain aroma and appearance for rice lines in Uganda was recommended by the Uganda National Rice Development Strategy with target of achieving the results by 2018 (MAAIF, 2009). Both the aroma and the grain appearance are influenced by genotype and environmental conditions (Xie et al., 2019). In regards to presentational qualities, some grain attributes of rice from Uganda including grain length were suggested to have met the market requirement specification (Lamo et al., 2017), but grain aroma which was also reported to be associated with the grain shape (MAAIF, 2020) was not one of the attributes for the recently released varieties in Uganda. Furthermore, despite of some efforts being directed at identification of aromatic rice varieties in Uganda (Akwero et al., 2020), the issue of adaptability has affected use of primary introduction as a breeding strategy and yet deliberate study on inheritance of aroma trait has so far been limited. One of the main problems faced by plant breeders for improving varieties is to select good parents and better insights on the combining abilities of the parents as pre-requisite for developing progenies for trait selection. The objective of this study was to establish the inheritance patterns for grain appearance and aroma traits.
MATERIALS AND METHODS
Description of the study area
The experiment was conducted both under screen house and field conditions at National Crops Resources Research Institute (NaCRRI), Namulonge, Uganda during the second rainy season of 2018. The site is characterized by bimodal rainfall, with the first rainy season starting from March to May and the second rainy season running from September to November with an annual rainfall intensity of 1,170 mm (Nsubuga, 2011). The soils are classified as Acric Ferralsol and are characterized by slightly acidic (pH of between 5.3 and 6.2) and low fertility with predominantly clay-loam textural class (Lamo et al., 2017).
Experimental materials
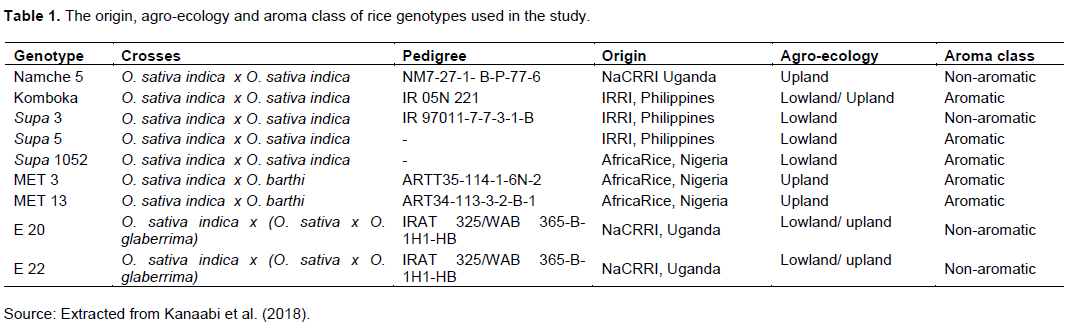
Parental materials used in the study comprised of nine rice lines with diverse backgrounds of Oryza sativa, Oryza barthi and Oryza glaberrima selected from the rice germplasm collection centre of NaCRRI with five highly-aromatic and four non-aromatic lines belonging to five sets of Multi-Environment Trial (MET) from Africa Rice Centre and Entry set from Uganda (‘E’), Namche and Komboka (Table 1).
Plant establishment and mating design
The seeds of each rice lines comprising of E 20, E22, Namche 5 and Supa 3 as the females and Komboka, MET 3, MET 13, Supa 5 and Supa 1052 used as male parents, were planted 2017B at a rate of three hills per individual line in 10 L, capacity plastic buckets filled with loam soil and raised in NaCRRI under screen house condition. At flowering, the plants were crossed using North Carolina Design II from the adjacent crossing block following the manual emasculation and hooking method (Peter et al., 1964). The resulting F1 seeds were planted in the plastic buckets in 2018A and raised in the screen house and the F2 seeds were harvested, dried and stored in paper bags separately for each bucket. The F2 seeds, alongside their parents, were planted in the field at NaCCRI in 2018B using randomized complete block design (RCBD) with two replications at a spacing of 20 cm × 20 cm (Lamo et al., 2017). The recommended agronomic practices of hand weeding (PRiDe, 2011) and bird control were followed to raise good crop stands. The F3 seeds harvested from panicles of the mature stands (Counce et al., 2000) on the account of each individual hill, comprising the tillers derived from a single seed were sun-dried until the moisture content determined using a moisture-meter was less than 14% (He et al., 2018). The dried grains per plant were kept individually in paper bags at room temperature of 25°C in the Bio-analytical and Nutrition Laboratory unit at NaCRRI until laboratory analyses. Two days prior to laboratory analyses, one sample of the stored F3 seeds on per hill basis were drawn from paper bags and dehulled using a porcelain mortar and pestle.
Assessment of grain appearance
Grain shape was determined by measuring the grain length and width (mm) using transparent plastic ruler and an average length of ten whole rice grains lined up lengthwise along the edge of a transparent plastic ruler and the breadth similarly determined (Singh et al., 2000). The mean grain length to mean width ratio of the grains for each sample was then determined as a measure of the grain shape (Graham, 2002). The grain length was described as short (< 5.5 mm), medium (5.51 - 6.60 mm), long (6.61 - 7.5 mm) and very long (> 7.5 mm) and the grain shape (length to width ratio) as: bold (< 2.1), medium (2.1 - 3.0), slender (> 3.0) (Khush et al., 1979).
Grain sensory aroma evaluation
Seven trained panelists in sensory aroma evaluation from the Nutrition and Bio-analytical Laboratory of National Crops Resources Research Institute were used to evaluate the rice grain aroma. The training consisted of two-hour sessions for 3 days prior to the grain aroma sensory evaluation. Forty grains of each cross and lines were soaked in 10 ml of 1.7% KOH solution in a covered and previously coded glass petri-dish for 30 min at room temperature following the method of Golam et al. (2010). The grain samples were randomized and aroma evaluation were done immediately after removing the petri-dish cover by using short sniffs. Four-point category scales (1 = no aroma, 2 = slight aroma, 3 = moderate aroma and 4 = high aroma) (Golam et al., 2010), were used to score the aroma intensity. The mean aroma score for each sample was obtained and classified into one of the four categories, namely non-aromatic (< 1.5); slightly aromatic (between 1.5 and 2.5) (Kamara et al., 2016); moderately aromatic (between 2.5 and 3.5) and highly aromatic (> 3.5) (Ocan et al., 2019; Srigastava et al., 2019).
Statistical analyses
Data generated from individual plants were entered in Microsoft Excel sheet and summarized into average on the replication basis. Unbalanced general analysis of variance (ANOVA) for the mean individual genotype data was carried out using GenStat statistical software (Version 12th Edition; VSN International Ltd., UK, 2009) and the means separated using Fishers protected least significance difference (LSD) at 5% significance level. The ANOVA followed a RCBD statistical model:
Yijk = Y + Gi + Rj + eijk
Where: Yijk = observation of genotype in replication j, and block k; Y = the general mean; Gi = effect of genotype i; Rj= effect of replication j and eijk = error of the observation ijk.
The genotypic difference among grain parameters of F2 segregates was determined by F test:
F(df1, df2) = MSg/MSe
Where: df1and df2 are the degrees of freedom associated with the numerator and denominator of the F ratio and MSg and MSe are the genotype and error mean squares, respectively.
For the analysis of combining ability and gene action, in relation to NCDII, RCBD consisting of 2 replications, each contained 20 F2 segregates and nine parental lines, the individual data were subjected to general ANOVA using linear model shown below:
Yijk= Y + GCAi + GCAj + SCA + Bk + eijk
Where: Yijk = mean of a specific cross; Y = Grand mean; GCA = general combining ability, Effect of the parent in the genotypic mean of its crosses, i = female, j = male; SCA = specific combining ability; Phenotypic value of a specific cross compared to the value predicted from parental GCA values; Bk = Block effects; Eijk = error effect.
The mean of F3 seeds were subjected to general ANOVA and regression using GenStat computer package (Release 12.1, PC windows 7, VSN International Ltd, 2009) to determine the variance of general combining ability (GCA) and specific combining ability (SCA) and direction effects. The SCA were calculated by subtracting predicted means from observed means and the significance of GCA and SCA were tested using a standard t test [T=effect/ (standard error of the effect)]. The error variance (error mean square, MSe) obtained from ANOVA of individual replication data analysis was used to determine the F-values for the sources of variations. The MSe was used as denominator in the F-values for testing combining abilities as:
F value m MSg/MSe; F value = MSs/MSe
Where: MSg, MSs and MSe are mean square due to GCA, SCA and error, respectively.
F-probability for the respective F values were tested using F-distribution function based on the degree of freedom for GCA and SCA as nominator (df1), each tested against degree of freedom associated with the denominator (MSe) of the F ratio, respectively. The variance of GCA and SCA for the variables and mean squares were estimated and used to calculate coefficient of genetic determination and Bakers ratios according to Baker (1978). Heritability was estimated according to the relationship between additive (s2a), genotypic (s2g) and phenotypic (s2p) variances as: Narrow sense coefficient of genetic determination (NS-CGD, h2), a proportion of phenotypic variation (s2p or phenotypic variance) attributed only to additive gene effect (s2a = additive variance) (Falconer and Makay, 1996), was calculated as:
h2 = (σ2GCAf + σ2GCAm)/(σ2GCAf + σ2GCAm +σ2SCA +σ2e) = s2a / s2p
Where, e = Sample error; (σ2GCAf + σ2GCAm) = Additive gene effect and (σ2GCAf + σ2GCAm +σ2SCA + σ2e) = Phenotypic effect.
Broad sense coefficient of genetic determination (BS-CGD, H2), a proportion of all phenotypic variation (s2p) due to all genetic effect (s2g) (Falconer and Makay, 1996), was calculated as:-
H2 = (σ2GCAf + σ2GCAm +σ2SCA)/ (σ2GCAf + σ2GCAm +σ2SCA +σ2e)
Where, e = Sample error; (σ2GCAf + σ2GCAm +σ2SCA) = Total genetic effect; and (σ2GCAf + σ2GCAm +σ2SCA + σ2e) = Phenotypic effect.
Baker’s ratio was calculated as:
Baker’s ratio (BR) = (σ2GCAf + σ2GCAm)/(σ2GCAf + σ2GCAm +σ2SCA)
Where, σ2 = sample variance; (σ2GCAf + σ2GCAm) = additive gene effect; and (σ2GCAf + σ2GCAm + σ2SCA) = Total genetic effect.
In addition, estimates of the relative significance between additive and non-additive effects was calculated based on the ratio of GCA to SCA, where a ratio of greater than unity (1) would mean predominance of GCA effect.
RESULTS
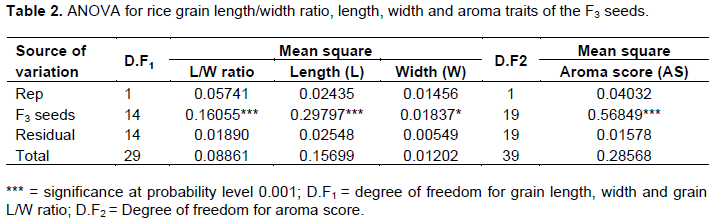
The results for the analysis of variance (ANOVA) for rice grain traits and aroma traits of the F3 seeds are presented in Table 2. The F3 seeds exhibited a significant difference (P < 0.001) among themselves for the rice grain length/width ratio, length, and aroma traits and (P < 0.05) for width.
Phenotypic performance of F2 plants and their parents
The mean scores for grain traits and aroma of F3 seeds and their parents are shown in Table 3. The parental lines had grain length/width ratio ranging from 2.94 to 3.84 and for the F3 seeds a range of between 2.92 and 3.70. Of the 24 genotypes including the parents, the F3 seeds of four genotypes (20%) and 20 genotypes (80%) were classified as medium and slender shaped grains, respectively. In the case of the grain length, a range of between 6.85 and 7.87 mm were determined whereas a range of between 6.75 and 8.05 mm, and 6.85 and 7.87 mm, respectively were recorded for the parental lines and the F3 seeds. Out of the 24 parental and F3 seeds, 13 (54.2%) had long grain and 11 (45.83%) parental and F3 seeds were classified as very long grains (Khush et al., 1979). For the grain width, the parental and F3 seeds registered values ranging from 1.95 to 2.50 mm and from 2.09 to 2.41 mm, for the parental and F3 seeds, respectively.
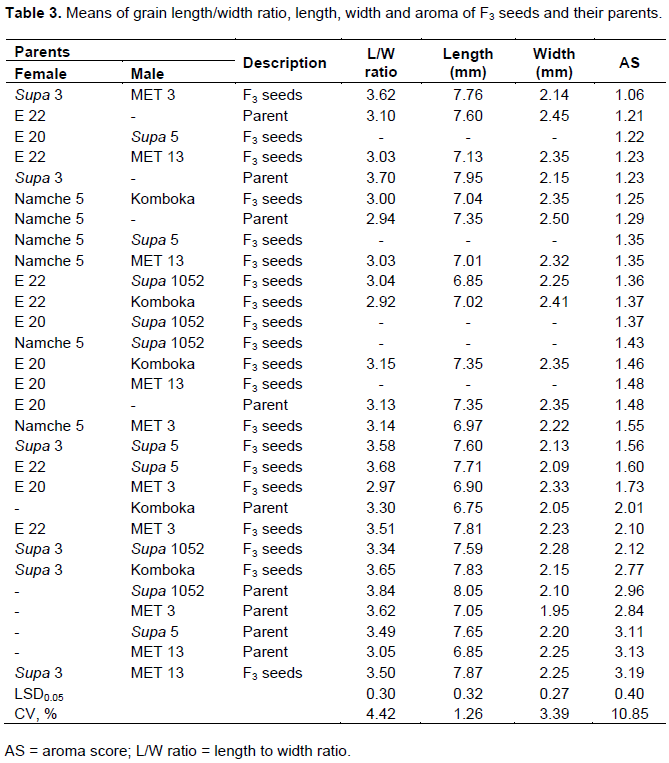
The aroma intensity of the genotype determined through sensory evaluation (Golam et al., 2010), ranged from 1.06 to 3.19 and the perceived aroma score for the aromatic parents of Komboka, Supa 1052, MET 3, Supa 5 and MET 13 from 2.01 to 3.11. The parental seeds registered aroma score ranging from 1.21 to 3.13, as opposed to the crosses with values ranging from 1.06 to 3.19. Four crosses (20%) of E 20 x MET 3, Supa 3 x Supa 1052, Supa 3 x Komboka and Supa 3 x MET 13 recorded aroma intensity higher than the aromatic parent of Komboka rice line which exhibited the aroma intensity of 2.01. The F3 seeds from Supa 3 and MET 13 had the highest perceived aroma intensity of 3.19 in relation to all the genotypes studied. Based upon Hien et al. (2006) classification for aroma intensity, only two F3 seeds of Supa 3 x MET 13 and Supa 3 x Komboka (score > 2.5) were highly aromatic and Supa 3 x Supa 1052 and E 22 x MET 3 (score of 2.0 - 2.5) exhibited moderate aroma intensities. F3 seeds derived from crosses between Namche 5 x MET 3, Supa 3 x Supa 5, E 22 x Supa 5 and E 20 x MET 3 showed slight aroma (score of 1.50 - 1.99) and the remaining 13 genotypes were non-aromatic. The results for analysis of variance for GCA of the parents and SCA of the crosses and the respective variance components, coefficient of genetic determination and Baker’s ratio for the grain quality traits are presented in Table 4.
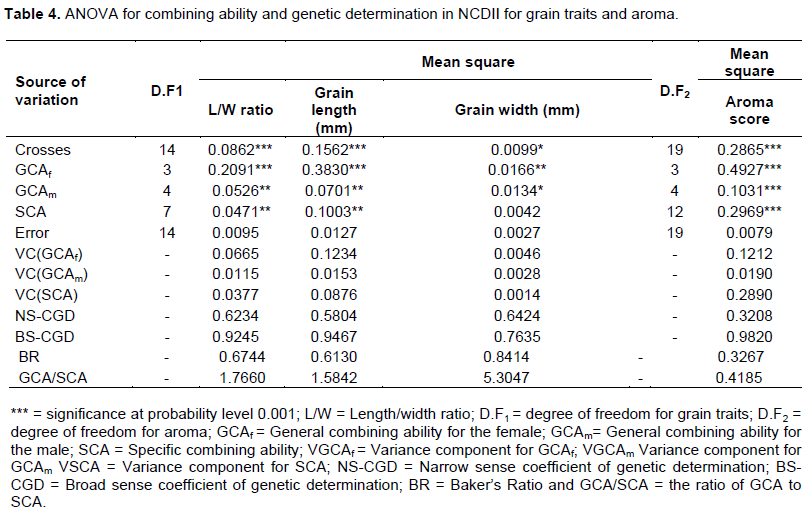
The analysis of variance of 20 F2 segregates and their 9 parents for general and specific combining ability showed that the grain length/width ratio, length, width and aroma traits were significantly (P < 0.05) different, but the results of the SCA for the grain width trait were non-significant. The combined GCA for the female and male, were higher than that for the SCA for all the traits except for the aroma. The genetic determination of the NSCGD ranged from 0.58 to 0.64 for grain traits, but aroma recorded a relatively lower value of 0.32. Considering BSCGD, all the traits recorded high values falling between 0.76 and 0.98. The Baker ratio and the grain traits recorded values ranging between 0.61 and 0.84 while the aroma trait registered a lower value of 0.33. The results also showed high GCA/SCA ratio for grain traits, ranging from 1.6 to 5.3 and a lower GCA/SCA ratio of 0.42 for aroma trait.
Combining ability effects
General combining ability effect
The general combining ability effects for the grain length, grain width, grain length/width ratio and aroma of parents estimated during the second season of 2018 are presented in Table 5.
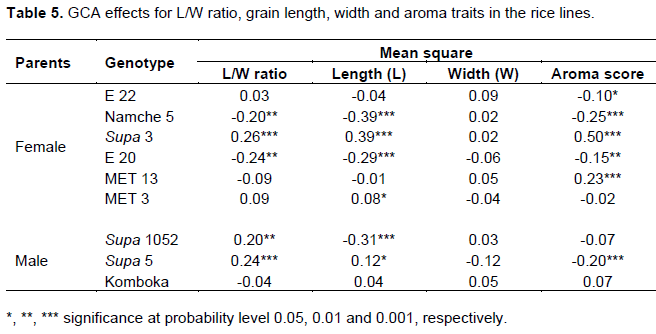
The results of analyses of combining ability effects for the L/W ratio indicated a significant (P<0.0015) positive GCA effect for Supa 3, Supa 5 and Supa 1052, and a significant (P<0.0105) negative GCA effect of Namche 5 and E 20. Similarly, the GCA effects for the grain length for Supa 3, Supa 5 and MET 3 were also significant (P < 0.05). Namche 5, E 20 and Supa 1052 however, recorded a negative significant (P < 0.05) GCA effects. The grain width unlike the rest of the grain traits showed non-significant GCA effect. In the case of aroma, Supa 3 and MET 13 exhibited high positive significant (P < 0.001) SCA effect (P < 0.001), while E 22, Namche 5, E 20 and Supa 5 expressed negative significant (P < 0.05) SCA effects.
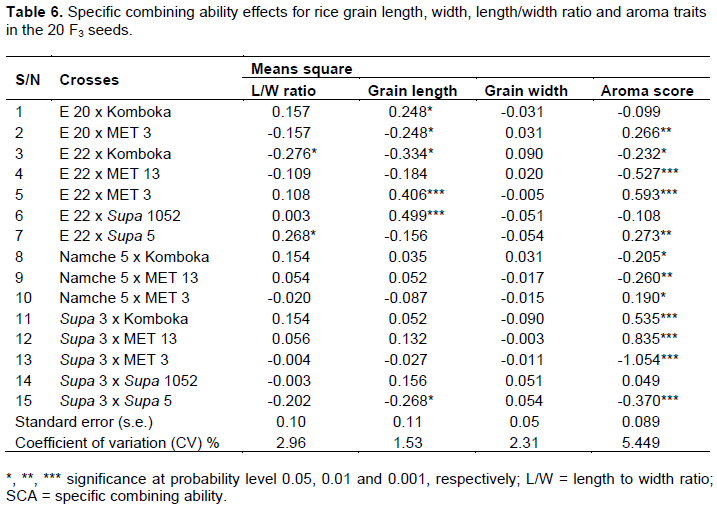
Specific combining ability effect
The results of specific combining ability effects in the crosses for grain length, width, length/width ratio and aroma traits are presented in Table 6. The SCA values for L/W ratio ranged between -0.276 and 0.268 with only one cross from E 22 x Supa 5 showing a significant (P < 0.05) positive SCA effect and also one F3 from E 22 x Komboka with a negative significant (P<0.05) SCA effect. The SCA for the grain length ranged from -0.334 to 499 and with the cross combination from E 20 x Komboka, E 22 x MET 3 and E 22 x Supa 1052 exhibiting positive significant (P<0.05) SCA whereas a negative significant (P<0.05) SCA effect were observed for the F3 seeds of Supa 3 x Supa 5, E 20 x MET 3 and E 22 x Komboka. The grain length did not show any significant SCA effects. The SCA effects of the F3 seeds for aroma trait ranged from -1.054 to 0.835 and six (40 %) crosses were observed to exhibit a positive significant (P < 0.05) positive SCA effects for the aroma trait.
DISCUSSION
The significant (P < 0.05) variation observed in the grain length/width ratio, length and width and the grain aroma intensity of the F3 populations, indicated sufficient existence of diversity in the parents used in the study implying that the crosses could be employed in the grain quality improvement through selection for superior grain traits. The variability in rice grain traits including length/width ratio, length, width as well as aroma trait among rice genotypes have previously been reported (Kamara et al., 2017; Kitara et al., 2018).
Grain shape for the crosses were broadly within the range of the parental limits, which is in line with Kumar (2016) who reported L/W ratio of crosses to be defined by the parental range. Furthermore, 80% of the gains for all the genotypes conformed to the preferred slender grain shape of the parents. Considering the mean grain length for the F3 seeds and their parents, all the genotypes possessed this long grains with 73.3% in this category of long grain representing grains classified as very long implying that grain length as a quality attribute was not a major limitation as was reported by Lamo et al. (2017) who suggested that Uganda rice genotypes had already met some grain quality specifications desired in the grain market. Genotypes from E 20 x Supa 1052 had grain length below the range of the parental lines, but the rest of the crosses were intermediate to the parental lines. Kumar (2016) reported grain length of some crosses to be slightly below or above those of the parental limits and in addition transgressive segregation for grain length was observed as was reported by Asante (2017), also suggesting possibility for improvement of this trait. According to Sharma and Jaiswal (2020), the performance of the crosses in regards to the quality rice grain traits of the shape, length, width and aroma were dependent on the choice of the parents. In the case of aroma, 25 % of the crosses exhibited moderate to high aroma intensity, suggesting that aroma trait was inherited from parents used which was consistent with the results reported by Kanaabi et al. (2018). A cross from Supa 3 x MET 13 expressed a higher score than the best aromatic parental line, which is an example of transgressive segregation, suggesting that some cross combinations could be employed in rice breeding program for aroma enhancement. According to Singh et al. (2017), crosses having both parents as good general combiner could result into superior transgressive segregants. The results further suggested that Supa 3 as female parent produced highly aromatic crosses compared to E 20, E 22 and Namche 5 backgrounds when used as female parents.
The significant (P<0.005) general combining ability (GCA), which defines average performance of parents (Griffing, 1956) as well as the significant GCA interaction effects implied involvement of both additive and non-additive gene effects in the inheritance of the rice grain traits (Sharma and Jaiswal, 2020). Recent study by Sharma and Jaiswal (2020) and Singh et al. (2020) indicated high significant GCA and SCA effects of the rice grain L/W ratio, length, width and aroma. According to Singh et al. (2020), GCA/SCA ratio of less than 1 implied preponderance of the non-additive gene action and therefore the higher GCA/SCA ratio (> 1) of the variance components for GCA to the SCA observed for all the traits apart from the grain aroma, implies predominance of additive gene effect. This finding is inconsistent with study by Srigastava et al. (2012) who reported that the grain length was conditioned by non-additive gene effects. A similar study conducted on Basmati for grain attributes reported the GCA/SCA ratio for the grain length to be above 1 and values lower than 1 in the case of the grain L/W ratio, width and aroma traits (Sharma and Jaiswal, 2020). In this study, with the GCA/SCA value for the appearance traits being above 1, it implies preponderance of additive gene action (Singh et al., 2020).
The moderate values for narrow sense coefficient of determination (NS-CGD) defining proportion of phenotypic variation attributed to additive gene variance (Falconer and Makay, 1996) and Baker’s ratio (BR) measuring the proportion of total genetic variance attributed to additive variance (Baker, 1978; Babu et al., 2012) implied moderate predictability for the inheritance of grain appearance traits (Falconer and Makay, 1996; Baker, 1978). On the other hand, the relatively lower NSCGD and GCA/SCA ratio for the aroma traits (Table 4) further indicated even much lower predictability for the expression of the aroma trait in rice.
The higher value for broad sense coefficient of determination (BS-CGD), which is defined by the proportion of total variance attributed to genetic variance (Falconer and Makay, 1996) across all the traits meant that both the appearance and aroma traits (Kamara et al., 2017; Xie et al., 2019) were mainly determined by genetic factors and minimally influenced by environmental conditions. It is also worth noting that although aroma presented comparatively lower values of the genetic determination compared to the appearance traits, the inheritance is still plausible (Kamara et al., 2017). Similarly, other researchers have reported aroma intensity to be influenced mainly by the genetic factors (Fasahat et al., 2016; Xie et al., 2019), besides environmental effects (Mo et al., 2019).
Combining ability effects for the parents and crosses
For the general combining ability effects, the parents with at least significant (P<0.05) positive GCA effects in this study were considered good combiners for the L/W ratio that is Supa 3, Supa 5 and Supa 1052, which are lowland rice types while Namche 5 and E 20 with significant and negative SCA effects were poor combiners. Following the same principle, Supa 3 and Supa 5 were good combiners for the grain length, while Supa 3 and MET 13 were good combiners for aroma, but a commonly grown Namche 5 upland rice variety had demonstrated a poor GCA implying limited possibilities of obtaining a superior aromatic upland rice variety based on deploying it as an aroma donor. On the other hand, E 22 and MET 3 both with poor GCA presented a cross combination with acceptable aroma intensity. In an inheritance study of grain traits, Singh et al. (2020) reported parents with good and poor GCA effects for length/ width ratio, length and width traits in rice.
Considering SCA in relation to the L/W ratio, cross E 22 x Supa 5 with significant (P < 0.05) positive SCA was a good combiner as opposed to E 22 x Komboka with significant (P < 0.05) negative SCA effect. In the case of the grain length, E 20 x Komboka, E 22 x MET 3 and E 22 x Supa 1052 with significant positive SCA values were good combiners unlike the Supa 3 x Supa 5, E 20 x MET 3 and E 20 x Komboka with significant negative SCA, hence designated poor combiners. Considering the aroma trait, the genotypes from Namche 5 x MET 3, E 20 x MET 3, E 22 x Supa 5, E 22 x MET 3, Supa 3 x Komboka and Supa 3 x MET 13 were good combiners due to their significant positive SCA effects, but Namche 5 x Komboka, Namche 5 x MET 13, E 22 x Komboka, Supa 3 x Supa 5, E 22 x MET 13 and Supa 3 x MET 13 with significant negative SCA values were considered poor combiners. Considering the best three scorers for L/W ratio in ascending order of magnitude were the crosses Supa 3 x Supa 5, Supa 3 x MET 13 and Supa 3 x Komboka, these were combination of parents with good x good, good x poor and good x poor GCA, respectively. Similarly, the top scoring crosses for grain length trait in ascending order of magnitude were the crosses E 22 x MET 3, Supa 3 x Komboka and Supa 3 x MET 13 corresponding to poor x good, good x poor and good x poor GCA, respectively. In the case of aroma, the best three scorers in ascending order of magnitude were the crosses Supa 3 x Supa 1052, Supa 3 x Komboka and Supa 3 x MET 13, which corresponded to the parental combination of good x poor, good x poor and good x good GCA, respectively showing that the case of transgressive segregation observed for the aroma trait was the result of the combination of two parents both with good general combining ability for the aroma trait. Sharma and Jaiswal (2020) reported transgressive segregation to be the result of a parental combination with both good combining ability, however, some F3’s with significant L/W ratio, for example from cross E 22 x Komboka, a cross combination of parents with moderate
x moderate GCA effects and cross E 22 x MET 3 with high significant SCA for grain length were from parents with moderate GCA effects. Similarly for the aroma, cross E 22 x Supa 5 was of parents which were both poor general combiners, but the F2 segregates had similar effects. A similar study on the grain quality traits of rice reported parents with various GCA combinations for the crosses exhibiting significant SCA effects (Sharma and Jaiswal, 2020). According to Akanksha and Jaiswal (2019), the significant SCA which resulted from the parents with high GCA × high GCA, high GCA × low GCA and low GCA × low GCA were attributed to additive × additive gene action, additive × non-additive gene action and non-additive × non-additive gene action interactions, respectively.
CONCLUSION
The results showed that both additive and non-additive gene effects were controlling the expression of the grain traits and aroma of rice seeds, but with preponderance of additive gene effect for the grain traits and prevalence of non-additive gene effect in the case of aroma indicating that early selection for aroma based on sensory evaluation technique would not be possible. In the case of aroma trait, the cross of Supa 3 x MET 13 was best for the advancement of the trait.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
The authors are grateful to Makerere-Sida Programme for the research funds and National Crops Resources Research Institute, Uganda for providing facilities for development and evaluation of the crosses. Professor Paul Gibson and Dr. Angele Pembele Ibanda of Makerere University are acknowledged for backstopping in the data analyses process.
REFERENCES
|
Akanksha, Jaiswal HK (2019). Combining ability studies for yield and quality parameters in basmati rice (Oryza sativa L.) genotypes using diallel approach. Electronic Journal of Plant Breeding 10(1):9-17. |
|
|
Akwero A, Ocan D, Akech W, Lamo J, Ochwo-Ssemakula M, Rubaihayo PR (2020). Allelic variations in aroma gene in cultivated rice varieties. African Crop Science Journal 28(2):241-254. |
|
|
Asante DM (2017). Breeding rice for improved quality. In: Li J (ed) Advances in International Rice Research. IntechOpen pp. 69-89. |
|
|
Babu RV, Shreya K, Singh DK, Usharani G, Shankar AS (2012). Correlation and path coefficient analysis studies in popular rice hybrids of India. International Journal of Scientific and Research Publications 2(6):1-5. |
|
|
Baker RJ (1978). Issues in Diallel Analysis. Crop Science 18(4):533-536. |
|
|
Candia AOT (2013). Effect of paddy drying depth using open-sun drying on drying time and mill recovery of Kasio variety in eastern Uganda. Uganda Journal of Agricultural Sciences 14(2):37-47. |
|
|
Counce PA, Keisling TC, Mitchell AJ (2000). A uniform, objective, and adaptive system for expressing rice development. Crop Science 40(2):436-443. |
|
|
Calingacion M, Fang L, Quiatchon-Baeza L, Mumm R, Riedel A, Hall RD, Fitzgerald MA (2015). Delving deeper into technological innovations to understand differences in rice quality. Rice 8(6):1-10. |
|
|
Falconer DS, Mackay TFC (1996). Introduction to Quantitative Genetics. 4Th ed., Longman Group. New York. |
|
|
Fasahat P, Rajabi A, Rad JM, Derera J (2016). Principles and utilization of combining ability in plant breeding. Biometrics and Biostatistics International Journal 4(1):1-22. |
|
|
Fitzgerald MA, McCouch, SR, Hall RD (2008). "Not just a grain of rice: the quest for quality". Trends in Plant Science 14(3):133-139. |
|
|
GenStat (2009). GenStat Release 12.1 (PC/Windows Vista). GenStat Procedure Library Release PL20.1. VSN International Ltd, UK. |
|
|
Golam F, Norzulaani K, Jennifer AH, Subha B, Zulqarnain M, Osman M, Nazia AM, Zulqarnian M, Mohammad O (2010). Evaluation of kernel elongation ratio and aroma association in global popular aromatic rice cultivars in tropical environment. African Journal of Agricultural Research 5(12):1515-1522. |
|
|
Graham R (2002). IRRI discussion paper series. In: A proposal for IRRI to establish a grain quality and nutrition research center. pp. 1-15. |
|
|
Griffing B (1956). Concept of General and Specific Combining Ability in Relation to Diallel Crossing Systems, pp. 464-492. |
|
|
He F, Qian Y, Zhang Y, Zhang M, Qian MC (2018). Aroma compounds generation in brown and polished rice during extrusion. Flavour Science pp. 103-106. |
|
|
Hien N, Yoshihashi T, Sarhadi WA, Thanh VC, Oikawa Y, Hirata Y (2006). Evaluation of aroma in rice (Oryza sativa L.) using KOH method, molecular markers and measurement of 2-acetyl-1-pyrroline concentration. Japan Journal of Tropical Agriculture 50(4):190-98. |
|
|
Kamara N, Asante MD, Akromah R (2017). Inheritance studies of aroma and yield attributes in rice (Oryza sativa L.) genotypes. African Journal of Agricultural Research 12(50):3461-3469. |
|
|
Kanaabi M, Tusiime G, Tukamuhabwa P, Andaku J, Ocan D, Jimmy L (2018). Evaluation of rice germplasm reveals sources of bacterial leaf streak disease resistance in Uganda. Journal of Food Security 6(4):163-169. |
|
|
Khush GS, Paule CM, DeLa-Cruz MN (1979). Rice quality evaluation and improvement at IRRI. In Proceedings of the workshop on chemical aspects of rice grain quality, International Rice Research Institute. Los Banos, Laguna, Philippines pp. 21-31. |
|
|
Kijima Y, Otsuka K (2017). Causes and consequences of NERICA adoption in Uganda: In: K. Otsuka and D.F. Larson (eds.), An African Green Revolution: Finding Ways to Boost Productivity on Small Farms pp. 123-142. |
|
|
Kitara IO, Lamo J, Edema R, Gibson, P, Rubaihayo PR (2018). Amylose content and grain appearance traits in rice genotypes. African Crop Science Journal 27(3):501-513. |
|
|
Kumar V (2016). Characterization, inheritance and allelic relationship study of gene(s) governing aroma and quality traits in rice (Oryza Sativa L.). Ph.D. Thesis, College of Agriculture, Indira Gandhi Krishi Vishwavidyalaya, Raipur, Chhattisgarh pp. 1-231. |
|
|
Lamo J, Tongoona P, Sie M, Semon M, Onaga G, Okori P (2017). Upland Rice Breeding in Uganda: Initiatives and Progress. Advances in International Rice Research pp. 214-246. |
|
|
Ministry of Agriculture Animal Industries and Fisheries (MAAIF) (2020). Draft annual performance report, financial year 2019-2020. pp. 1-150. |
|
|
Ministry of Agriculture Animal Industries and Fisheries (MAAIF) (2009). Uganda National Rice Development Strategy (UNRDS) (2nd Draft). Entebbe, Uganda: Ministry of Agriculture Animal Industry and Fisheries (MAAIF). |
|
|
Menya E, Olupot PW, Storz H, Lubwama M, Kiros Y (2018). Characterization and alkaline pretreatment of rice husk varieties in Uganda for potential utilization as precursors in the production of activated carbon and other value-added products. Waste Management 81:104-116. |
|
|
Mo Z, Li Y, Nie J, He L, Pan S, Duan M, Tian H, Xiao L, Zhong K, Tang X (2019). Nitrogen application and different water regimes at booting stage improved yield and 2-acetyl-1-pyrroline (2AP) formation in fragrant rice. Rice 12(74):1-16. |
|
|
Mogga M, Sibiya J, Shimelis H, Lamo J, Ochanda N (2018). Appraisal of major determinants of rice production and farmers' choice of rice ideotypes in south Sudan: implications for breeding and policy interventions. Experimental Agriculture 55(1):1-14. |
|
|
Nsubuga FNW (2011). Climatic trends at Namulonge in Uganda, JGG 3(1):119-131 |
|
|
Ocan D, Zhang R, Odoch M, Kanaabi M, Ibanda AP, Akwero A, Khizzah BW, Nuwamanya E, Lamo J, Fitzgerald MA, Daygon VD, Rubaihayo PR (2019). Relationship between 2-acetyl-1-pyrroline and aroma in Uganda rice populations with Oryza (barthi, glaberrima and sativa) backgrounds. African Journal of Biotechnology 18(31):1016-1024. |
|
|
Olupot PW, Candia A, Menya E, Walozib R (2015). Characterization of rice husk varieties in Uganda for biofuels and their techno-economic feasibility in gasification. Chemical Engineering Research and Design 107:63-72. |
|
|
Peter RJ, Beachall HM, Chuaviroj M (1964). An Improved Rice Hybridization Technique 1. Crop Science 4(5):524-526. |
|
|
Plengsaengsri P (2019). Optimization of process conditions for the development of rice milk by using response surface methodology. IOP Conf. Ser., Environmental Earth Sciences 346(1):012080. |
|
|
Prodhan ZH, Faruq G, Taha RM, Rashid KA (2017). Agronomic, transcriptomic and metabolomic expression analysis of aroma gene (badh2) under different temperature regimes in rice. International Journal of Agriculture and Biology 19(3):569-576. |
|
|
Promotion of Rice Development Project (PRiDe) (2011). Rice cultivation handbook. National Crops Resources Research Institute (NaCRRI)/Japan International Cooperation Agency (JICA), Uganda. pp. 1-45. |
|
|
Sen S, Chakraborty R, Kalita P (2020). Rice - not just a staple food: A comprehensive review on its phytochemicals and therapeutic potential. Trends in Food Science and Technology 97:265-285. |
|
|
Sharma A, Jaiswal HK (2020). Combining ability analysis for grain yield and quality traits in basmati rice (Oryza sativa L.). Plant Arch 20(1):2367-2373. |
|
|
Singh RK, Singh US, Singh GS (2000). Aromatic Rices. Oxford and IBH publishing Co. Pvt. Ltd. New Delhi, India pp. 1-300. |
|
|
Singh MK, Singh S, Nautiyal MK, Pandey ID, Gaur AK (2017). Variability, heritability and correlation among grain quality traits in basmati rice (Oryza sativa L.). International Journal of Chemical Studies 5(5):309-312. |
|
|
Singh MK, Banshidhar, Kumar K, Tigga A, Singh SK (2020). Exploration of possibilities to identify heterotic cross combinations in aromatic rice (Oryza sativa L.) for grain yield and quality parameters. Current Journal of Applied Science and Technology 39(11):92-98. |
|
|
Srigastava AK, Jaiswal HK, Agrawal HK (2012). Genetics of grain quality traits in indigenous aromatic rice (Oryza sativa L.). Crop Research 44(1):135-140. |
|
|
Srigastava D, Shamim M, Mishra A, Yadav P, Kumar D, Pandey P, Khan NA, Singh KN (2019). Introgression of semi-dwarf gene in Kalanamak rice using marker-assisted selection breeding. Current Science 116(4):597-603. |
|
|
Xie LH, Tang SQ, Wei XJ, Jiao GA, Sheng ZH, Hu PS (2019). An optimized analysis of 2?acetyl?1?pyrroline content diversity in the core collection of aromatic rice germplasm (Oryza sativa L.). Cereal Chemistry 96(4):698-707. |
|
|
Yiga VA, Lubwama M, Olupot FW (2019). Effect of Alkaline Surface Modification and Carbonization on Biochemical Properties of Rice and Coffee Husks for Use in Briquettes and Fiber-Reinforced Plastics. Journal of Natural Fibers 18(4):620-629. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


