Full Length Research Paper

ABSTRACT
Common bean (Phaseolus vulgaris L.) is an important food consumed in every region in Republic of Benin; however, it remains a neglected and under-utilized crop. In order to assess the value of bean germplasm of the central region in the Republic of Benin for useful breeding programs, 57 accessions were collected from 23 villages. After a classification based on the morphological variables of the seeds, these accessions were evaluated using 30 morphological traits (18 qualitative and 12 quantitative) following the IBPGR descriptors in experimental field at Faculty of Sciences and Technology of Dassa. Based on the seeds morphological variability, the accessions have been grouped in 8 morphotypes. However, 9 morphological types were obtained with cluster analysis based on UPGMA classification method using qualitative variables, whereas in Principal Component Analysis (PCA) they were gathered into 4 clusters using quantitative variables. The accessions in cluster 1 (11 accessions) were identified as possessing the highest values in quantitative traits like days to maturity, and number of pods per plant. While accessions in cluster 2 (3 accessions) have had the lowest number of days to flowering and the highest pods length, the accessions of cluster 3 (23 accessions) presented the highest 100-seed weight. Correlation coefficient of 100-seed weight was positively significant (p ≤ 0.001) with leaf length, pod width, and seeds length while it was negatively correlated (p ≤ 0.05) with days to flowering, pod length and the number of seeds per pod. Cluster 3 accessions may serve as useful genetic material in future, for any breeding programmes to improve the productivity of other common bean accessions through hybridization.
Key words: Common bean, cluster analysis, diversity, landraces, qualitative traits, quantitative traits.
INTRODUCTION
Common bean (Phaseolus vulgaris L.) is the most cultivated and consumed legume throughout the world (Blair et al., 2009; Coelho et al., 2009). This legume is an annual and self-pollinated crop (Ferreira et al., 2000), which intensely grows throughout the whole tropical area and some temperate regions of the planet (Coelho et al., 2009; Hegay et al., 2012). It is widely cultivated in the tropics for its green edible leaves and green pods consumed as vegetables, and dried seeds harvested at maturity (De Luque et al., 2014). Common bean is an important food and source of dietary minerals that potentially provide all the 15 essential minerals (Welch et al., 2000), and daily protein requirements for humans (Broughton et al., 2003; Ulukapi and Onus, 2014). This legume is presumed as one of the basic components of African food, mostly in form of grain (Bode et al., 2013).
In the Republic of Benin, beans are important staple, but its production has declined since 2008 from 143,625 T to 95,794 T in 2014 (Food and Agricultural Organization [FAO], 2014). This decline in production can be explained by several biotic and abiotic stresses that affect the crop during its cultivation and storage (Beebe et al., 2013; Hinkossa et al., 2013; Yaqoob et al., 2013; De Luque et al., 2014; Asfaw and Blair, 2014; Polania et al., 2016). Moreover, these biotic and abiotic stresses lead to the disappearance of common bean landraces and their genetic erosion in Central region of Benin Republic (Missihoun et al., 2017). The preservation and efficient utilisation of existing common bean landraces in this region of Benin Republic requires detailed knowledge of their genetic variability such as agro-morphological characteristics. In addition, to increase farmers’ production and productivity of this legume, the development of cultivars with improved resistance to biotic and abiotic stresses is necessary (Miklas et al., 2006; Doumbia et al., 2013). Unfortunately, there are no reported studies on collection and characterisation of these common bean genetic resources in the central region of Benin Republic. Consequently, there is a need to collect, and characterise common bean populations in this region before they disappear for an efficient conservation and to guide breeding programs.
Common bean is known for its adaptability to different environments, creating a wide range of landraces (Bitocchi et al., 2012). An important varietal diversity would be thus managed by farmers. To be useful for plant breeders, genetic resources must be characterised by morphological and agronomic traits (Martins et al., 2006; Stoilova et al., 2013). In fact, morphological characterization allows clarification of some problems of synonymies that exist in local denomination of landraces and permits knowing the correlations between agronomic performances for breeding programmes (Balkaya and Karagac, 2005; Balkaya et al., 2010; Karaagac and Balkaya, 2013). The characterization of accessions also allows quantification and structuring of the genetic variability in the germplasm (Bode et al., 2013). The objective of this study is to assess the morphological diversity and agronomic performances of common bean landraces of the central region in the Republic of Benin which is highly important for breeding programmes, for the conservation and the preservation of this genetic resource.
MATERIALS AND METHODS
Plant material and study location
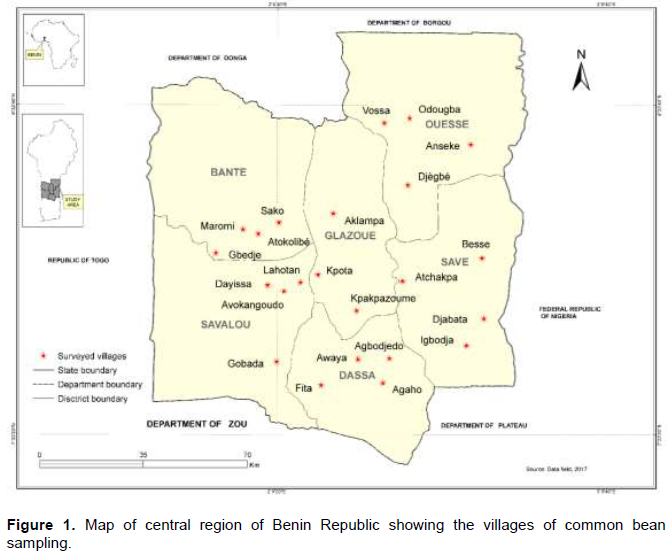
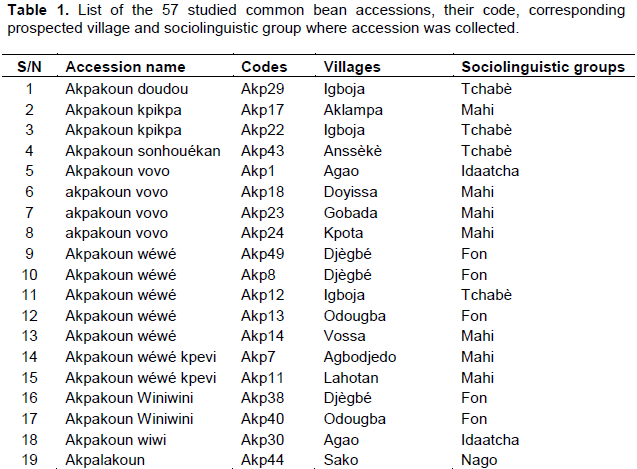
Fifty-seven common bean accessions were collected from 23 villages in the central region of the country (Figure 1 and Table 1). A trial was conducted under open field conditions at the experimental site of Faculty of Sciences and Technology of Dassa. The region is characterized by 4 seasons including two rainy seasons and two dry seasons with an annual rainfall ranging from 900 to 1100 mm (Akoègninou et al., 2006). The temperature varies from 24 to 29°C with an average of 27°C (Yabi and Afouda, 2012).
Morpho-agronomic characterisation
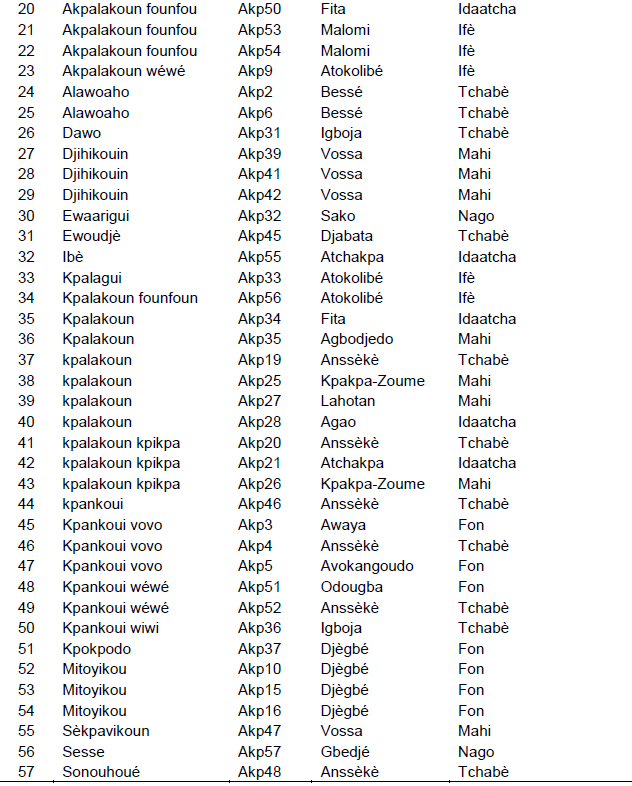
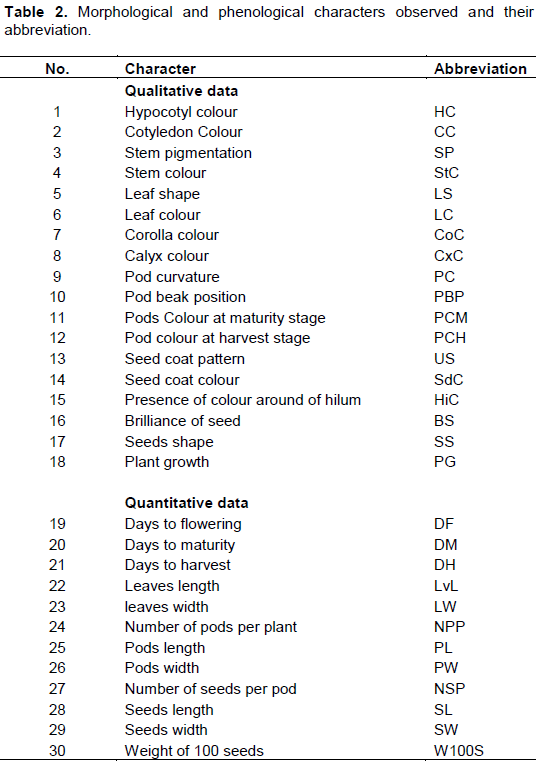
The 57 common bean accessions were firstly classified using visual technique following Mohammed et al. (2016) based on seed’s morphological description characteristics (coat colour, size, coat pattern and hilum colour). The agro-morphological characterization was conducted at the experimental site of the Faculty of Sciences and Techniques of Dassa, during the common bean cropping season of 2016 to 2017. The experimental design was a randomised complete block with five replicates. Each experimental unit consists of 4 rows measuring 4 m in length. The distance between rows of common beans was 1 m with 0.8 m between plants in the row using traditional farmers' production management in the study area. Three seeds were put in each hole, and 15 days after germination, seedlings were removed to leave only one seedling per hole. Weeds were removed manually from experimental plots. All accessions were assigned to creeping plants, staked and trained to climb the stakes (Rana et al., 2015). The descriptors used included eighteen (18) qualitative and twelve (12) quantitative variables (Table 2) among those recommended by IBPGR (1982). All of the observations were made according to the methods of IBPGR (1982).
Statistical analyses
Qualitative data were used to build a dendrogram with UPGMA (unweighted pair group method with arithmetic average) algorithm (Sneath and Sokal, 1973) using the sequential agglomerative hierarchical nested clustering (SAHN) of the NTSYS-pc software (Rohlf, 2000). Quantitative data were analysed by descriptive statistics with Minitab 17.1.0 software to determine the mean, minimum, maximum, standard deviation, and coefï¬cient of variation. To measure the degree of association between pairs of quantitative variables, Pearson correlation coefficient was calculated using Minitab 17.1.0 software. Principal Component Analysis (PCA) was also performed with Minitab 17.1.0 software to project on the axis the analysed accessions. For quantitative morphological traits, an analysis of variance (ANOVA) was performed with the aid of SPSS software Version 17.0. Significant differences between means were separated using Student Newman Keuls test (p < 0.05).
RESULTS
Distribution of phenotypic characters
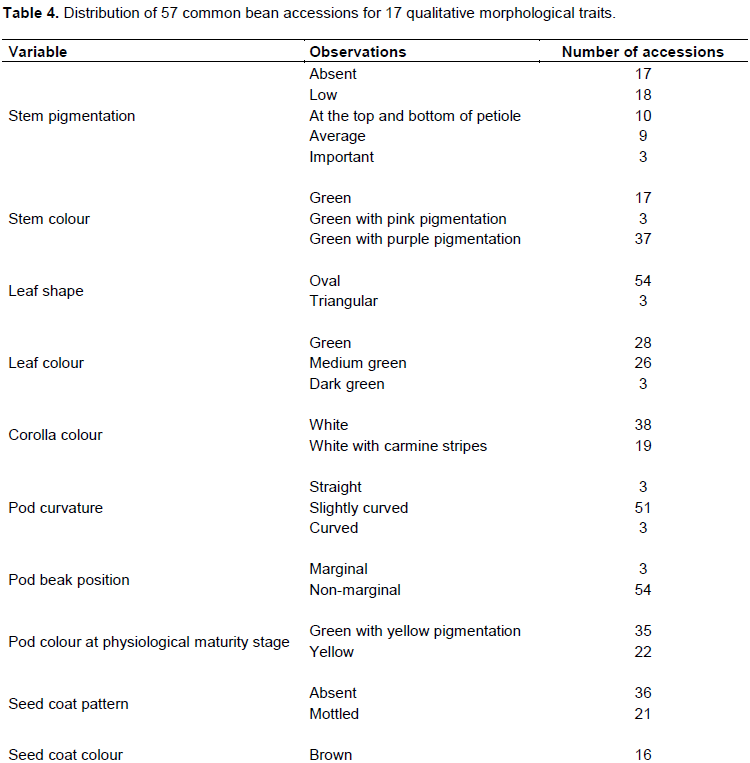
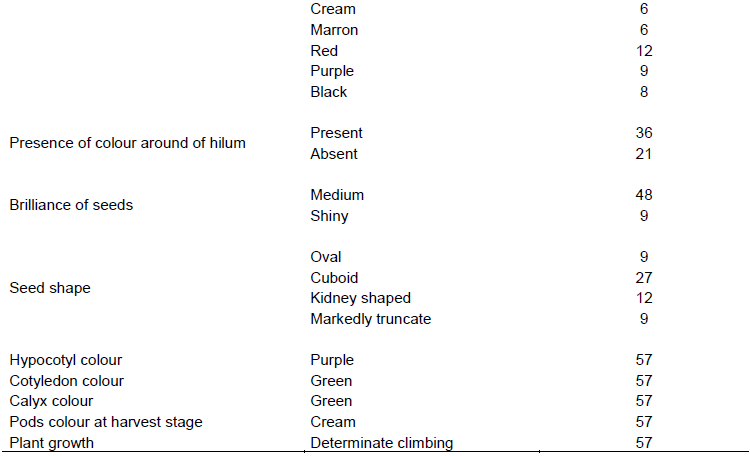
The 57 common bean accessions were classified in eight morphotypes according to the seed morphological description characteristics (Figure 2). The characteristics corresponding to the eight morphotypes were presented in Table 3. The analysis of qualitative data has shown that five traits (hypocotyl colour (purple), cotyledon colour (green), calyx colour (green), pods colour at harvest stage (cream), and plant growth (determinate climbing)) were not polymorphic, while the remaining showed a considerable level of variation (Table 4). Various seed’s colours were found with the brown (28.1% of accessions) and red (21.1% of accessions) seed’s coat colour as the most dominant (Table 4). A total of 63.2% of the germplasm had no seed coat pattern. Most accessions had cuboid seed shape (47.4%), medium brilliance (84.2%), and presence of colour around hilum (63.2%). A greater proportion of common bean accessions had green stem with purple pigmentation (64.9%), oval leaves shape (94.7%), green leaves colour (49.1%), white corolla colour (66.7%), slightly curved pod curvature (89.5%), non-marginal pod beak position (94.7%), and green pod with yellow pigmentation at physiological maturity stage (61.4%) (Table 4).

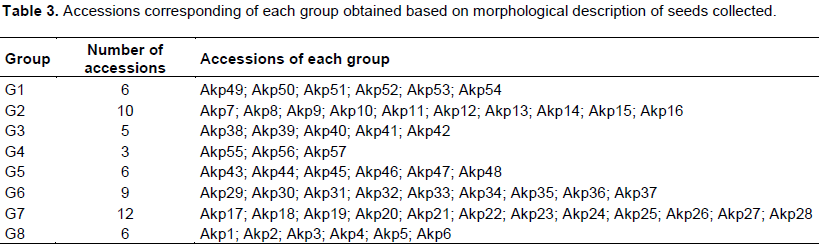
Morphological diversity
Cluster analysis based on morphological qualitative traits grouped the 57 accessions into 9 morphological types (Figure 3):
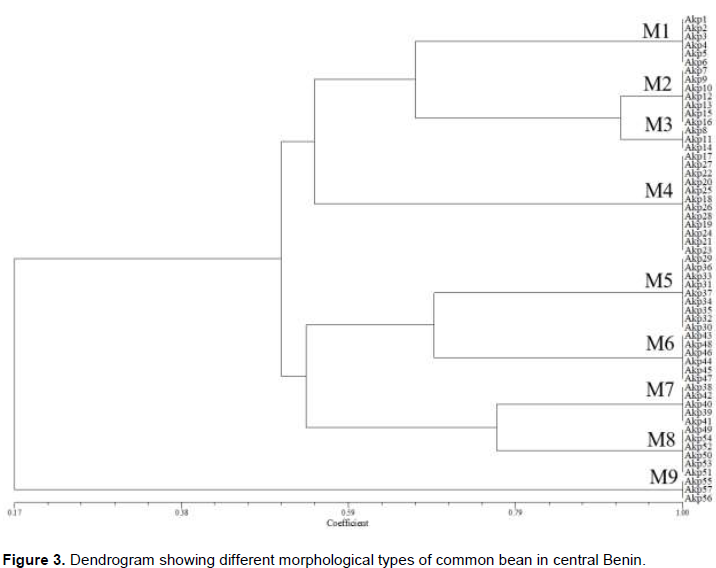
1) The first one (M1: 6 accessions) is characterized by low pigmentation of stem which have green colour with purple pigmentation, oval and green leaves, corolla coloured in white with carmine stripes, slightly curved pods, non-marginal pod beak position, pods having a green with yellow pigmentation at physiological maturity stage, mottled seeds coat pattern, cream seeds coat colour, presence of colour around hilum of seeds, medium brilliance, and oval seeds.
2 The second morphological type (M2: 7 accessions) was different from the first one by stem having pigmentations at the top and bottom of petiole, pods having a yellow pigmentation at physiological maturity stage, absence of mottle in seeds coat, white colour of seeds coat, cuboid shape of seeds, and slightly curved pods.
3) The characteristics of the third morphological type (M3: 3 accessions) are similar to those of the second morphological type (M2) except the curved pods.
4) The fourth morphological type (M4: 12 accessions) is characterized by green with pink pigmentation colour of leaves, white colour of corolla, red colour of seeds coat, and presence of colour around seeds of hilum.
5) The fifth morphological type (M5: 9 accessions) have an average pigmentation of stem, pods having a green with yellow pigmentation at physiological maturity stage, mottled seeds coat, purple colour of seeds coat, absence of colour around seeds of hilum, and markedly truncated shape of seeds.
6) The sixth morphological type (M6: 5 accessions) is characterized by the absence of stem’s pigmentation, green colour of stem, green leaves, absence of mottle on seeds coat, cream colour of seeds coat, and cuboid seeds.
7) The characteristics of the seventh morphological type (M7: 6 accessions) are mottled seeds coat, chestnut colour of seeds’ coat, and kidney shape of seeds.
8) The eighth morphological type (M8: 6 accessions) is characterized by a medium green colour of leaves, absence of mottle on seed’s coat, absence of colour around hilum seeds, and shiny brilliance of seeds.
9) The ninth morphological type (M9: 3 accessions) is characterized by an important pigmentation of the stem, green with pink pigmentation of stem, triangular shape of the leaves, dark green colour of leaves, corolla having white colour with carmine stripes, straight curvature of pods, marginal beak position of pods, purple colour of the seeds’ coat, and oval seeds.
Quantitative variations
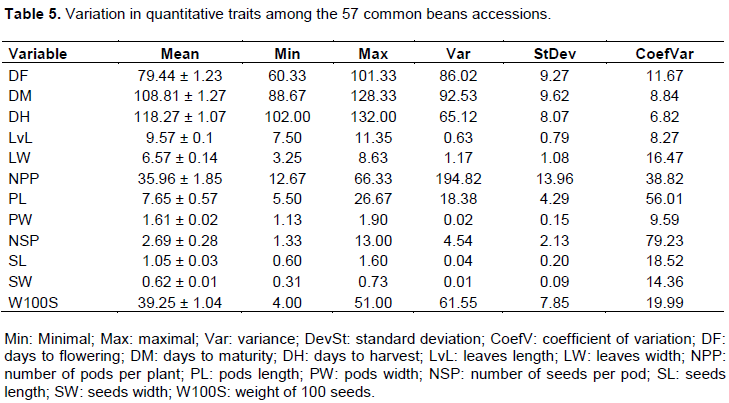
The mean, maximum, minimum, standard deviation and coefficient of variation of each of the 12 quantitative measured variables are presented in Table 5. The results indicated that variability among the accessions was significant (p=0.000) for the twelve traits. The results show that the variables such as the number of seeds per pod, the length of pods, and the number of pods per plant have the highest coefficient of variation (Table 5). According to results, days to flowering ranged from 60.3 (Akp38) to 101.3 days (Akp56) with a mean of 79.4 days and a standard deviation of 9.2. Mean days until physiological maturity was 108.81 and days to harvest were 118.27. The length of the leaves ranged from 7.50 (Akp20) to 11.35 cm (Akp55), with an average of 9.57 and a variation coefficient of 8.27%. Regarding the pods width, the range was 1.13 (Akp56) to 1.90 cm (Akp33), with an average of 1.61 cm and a variation coefficient of 9.59%. The length of the pods varied from 5.50 (Akp54) to 26.67 cm (Akp55) with an average of 7.65 cm. The length of the seeds varied from 0.60 (Akp57) to 1.60 cm (Akp8, Akp10 and Akp13) while seeds width ranged from 0.31 (Akp55, Akp56 and Akp57) to 0.73 cm (Akp9 and Akp14) with an average of 0.62 cm. The weight of 100 seeds ranged from 4 (Akp36) to 51 g (Akp14) with a mean number of seeds per pod of 2.69.
Phenotypic trait correlations
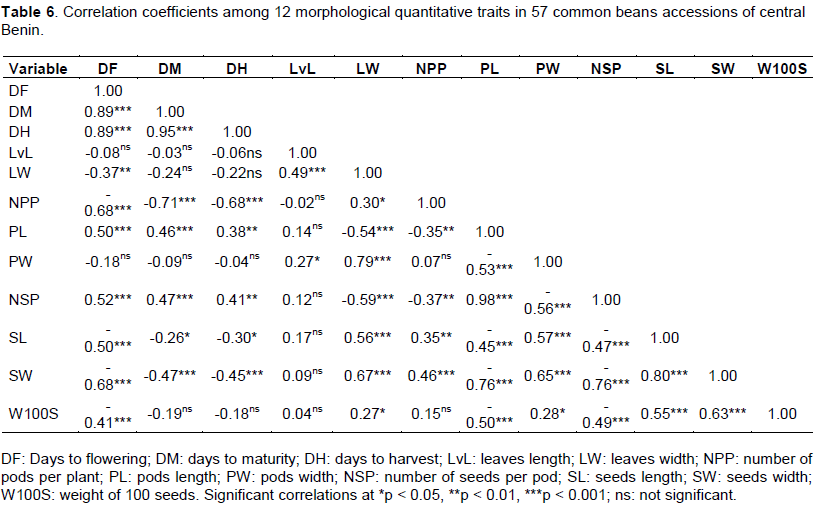
Correlation coefficients among traits are shown in Table 6. Indeed, the days to flowering (DF) was positively correlated with days to maturity (DM) (r = 0.894***), days to harvest (DH) (r = 0.890***), the length of pods (PL) (r = 0.503***), and the number of seeds per pod (NSP) (r = 0.518***); whereas, the days to flowering (DF) was negatively correlated with the width of leaves (LW) (r = -0.370**), the number of pods per plant (NPP) (r = -0.683***), the length seeds (SL) (r = -0.505***), the width of seeds (SW) (r = -0.679***), and the weight of 100 seeds (W100S) (r = -0.411***). The pods width was positively correlated with seeds length (SL) (r = 0.569***), seeds width (SW) (r = 0.652***), weight of 100 seeds (W100S) (r = 0.283*), leaves length (LvL) (r = 0.272*), and leaves width (LW) (r = 0.789***). However, the pods width was negatively correlated with the number of seed per pod (NSP) (r = -0.562***), and pods length (PL) (r = -0.534***). The weight of 100 seeds has a positive correlation with leaves width (LW) (r = 0.271*), pods width (PW) (r = 0.283*), seeds length (SL) (r = 0.549***), and seeds width (SW) (r = 0.627***), while it is negatively correlated with the days to flowering (DF) (r = -0.411***), pods length (PL) (r = -0.505***), and number of seeds per pod (NSP) (r = -0.487***).
Principal component analysis (PCA)
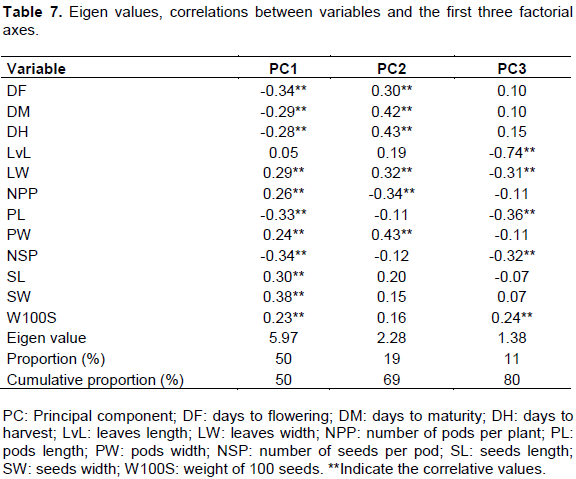
PCA showed that only the first three axes had an Eigen value higher than 1 and represent about 80% of total variability or dispersion (Table 7). Thus, most of the data structure can be captured in these dimensions. The remaining principal components account for a very small proportion of the variability and are probably less important. This has been confirmed by the fact that all measured variables are in correlation with these three axes. Indeed, except leaves length variable, the other eleven variables are in correlation with the first axis (LW, NPP, PW, SL, SW and W100S are positively correlated while DF, DM, DH, PL and NSP are negatively correlated) because the correlation of each of them is not close to zero (higher than 0.2). Leaves length variable has negative correlation with the third axis. Six variables (DF, DM, DH, NPP and PW) are simultaneously in correlation with the first and second axis while three variables (PL, NSP and W100S) are in correlation with the first and third axis. No variable is simultaneously in correlation with the second and the third axis but one variable (LW) is in correlation with the three axes alone. This analysis of the principal component showed that all variables are important for spatial representation of accessions study.
The correlation of the variables about the first and the second axis (loading plot) is as shown in Figure 4. Furthermore, observing the loading plot and the score plot obtained from principal component analysis, the studied accessions have been grouped in 4 clusters (Figure 5). The first one (11 accessions) is characterized by a high length of pods, number of seeds per pod, days to flowering, days to maturity and days to harvest while it is represented by low pods width, leaves width, seeds length, weight of 100 seeds, seeds width and number of pods per plant. The second cluster (3 accessions) contains accessions that have opposite performances to the first cluster. It is characterised by a high pods width, leaves width, seeds length, weight of 100 seeds, seeds width, and number of pods per plant while the pods length, number of seeds per pod, days to flowering, days to maturity and days to harvest are very low. The third and fourth clusters (23 and 20 accessions respectively) seem to belong to the same cluster because both clusters are located near the origin of the first axis; thus, they present variables that the values are closed to the means. However, the days to flowering, days to maturity and days to harvest of the third cluster are lower than the ones of the fourth cluster. The values of the pods width, leaves width, seeds length, weight of 100 seeds, and seeds width of the third cluster are on the other hand, higher than the ones of the fourth cluster.
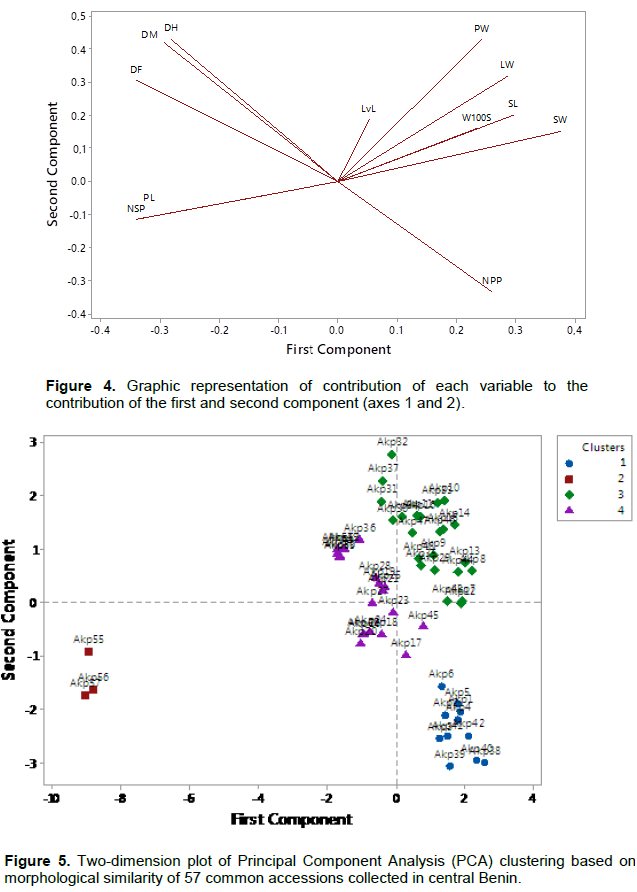
The comparison of the means of each variable using ANOVA enabled confirmation that the four clusters are significantly different (p ≤ 0.001). Therefore, the characteristics and accessions of each cluster are respectively presented in Table 8. This analysis showed that accessions of the Clusters 1, 3 and 4 have a high performance of weight for 100 seeds (W100S). But regarding the precocity of landraces, accessions of Cluster 1 have the best productivity.
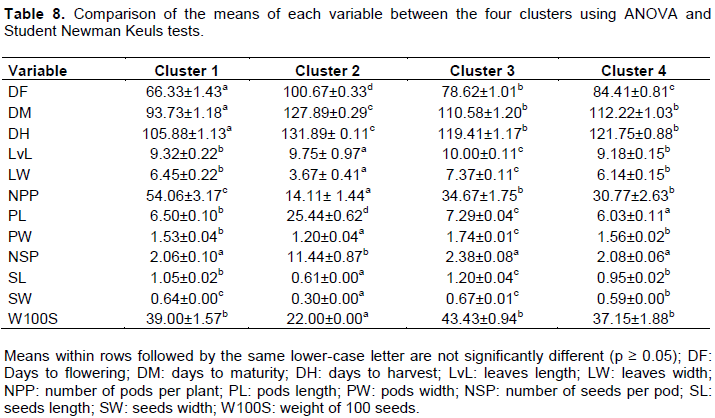
Comparing the four clusters coming from the quantitative data analysis with the ones from the qualitative data analysis, we have notified that: Cluster 1 gathers the morphological types M1 and M7, Cluster 2 is the same with morphological type M9, Cluster 3 contains morphological types M2, M3, M5 without Akp36 and M6 without Akp45, and Cluster 4 regroups the morphological types M4, M8, and accessions Akp36 of M5 and Akp45 of M6.
DISCUSSION
The results of this study showed that the common bean landraces collection of the central region of the Republic of Benin have a wide range of morphological and agronomic characteristics. The number of morphological types obtained from the studied accessions using the 17 qualitative variables (9 morphological types) was almost similar to the number of groups obtained only considering seed’s morphological description characteristics (8 morphotypes). Showing the importance of seed’s morphological characteristics as indicator of common bean diversity (Ulukapi and Onus, 2014), and to understand why folk nomenclature and taxonomy of this legume in central region of Benin Republic were mainly based on seeds’ coat colour (Loko et al., 2018). Moreover, some studies showed that descriptors linked to seeds are the most discriminant traits of common beans (Hegay et al., 2013; Stoilova et al., 2013; Ulukapi and Onus, 2014), and considered as highly heritable traits, therefore important for breeding programmes (Kumar et al., 2014).
The accessions bearing brown-red colour and cuboid shape dominated in common bean collection of central region of Benin Republic. Similar observations on the dominance of these colour and shape in common bean collections have been recorded in other parts of the world (Meza et al., 2013; Saba et al., 2016). Knowing that the colour, shape and size of common bean seeds are of special attention for consumers (Stoilova et al., 2013), these dominant seeds characteristics could reflect farmers and consumers preferences. Thus, to orient breeders with the morphological type of common bean to be improved for adoption by farmers and consumers, the findings of Loko et al. (2018) suggested that selection based on common bean seed coat colour would have a definite role in the framework of on-farm conservation of this legume in central Benin.
The growth habit of all common bean accessions was climbing type. The predominance of this growth habit type is probably related to ecological adaptation as well as to the cropping system being followed in the study area (Rana et al., 2015). Indeed, Missihoun et al. (2017) observed that in central Benin, common bean production generally involves intercropping in the fields with other crops such as maize, cassava, pigeon pea or oil palm tree. Aware of the fact that the scarcity of suitable materials for stacking is the main constraint of bean production in Central region of Benin Republic (Missihoun et al., 2017), it is important to create semi-climbers varieties for reduction of this constraint. The hybridisation of climbing bean type found in central Benin by a bush bean type namely Houintakpakun found in southern Benin (Missihoun et al., 2017) is necessary for enlarging the genetic base for increasing levels of resistance to both biotic and abiotic stress factor affecting common bean production in this region.
The cluster analysis based on morphological qualitative traits showed a significant number of similarities between accessions which seem to attest the presence of duplicates. Duplication of common bean landraces in numerous collections throughout the world were often highlighted by several authors (Chiorato et al., 2006; Madakbas and Ergin, 2011; Akhshi et al., 2014; Rana et al., 2015). In fact, in central region of Benin Republic, folk nomenclature of common bean landraces varies from one socio-linguistic group to another, and several folk varieties could be attributed to a single landrace and many landraces could have a similar name (Loko et al., 2018). To detect and identify these duplicates in common bean collections, molecular analysis was recommended by several authors (Singh et al., 1991; Madakbas and Ergin, 2011; Meza et al., 2013; Rana et al., 2015).
Descriptive statistics of quantitative data reflected high level of variation among the 57 common bean accessions. These finding could reflect their genetic differences and provide a good material for breeders. The average days of flowering (79.44) and the days until physiological maturity (108.81) of common bean accessions of central Benin is higher than those obtained in similar studies of common beans (Garcia et al., 1997; Gomez et al., 2004; Stoilova et al., 2005; Meza et al., 2013; Ulukapi and Onus, 2014). These differences could be explained by the fact that these physiological traits are influenced by environmental conditions, mainly temperature and photoperiod length (Meza et al., 2013).
Different correlations existing among quantitative variables showed that it is possible to improve several performances from breeding one of them. In fact, strongly correlated traits are possibly under the influence of the same genes which during selection could be selected simultaneously based on one of the traits (Okii et al., 2014). The significant positive correlation of seed weight with leaves width (LW), pods width (PW), seeds length (SL), and seeds width (SW) indicated that these characters are efficient in yield determination. Similar findings were reported by Rana et al. (2015). However, seeds’ weight was negatively correlated with days to flowering, pod length and number of seeds per pod. These findings are in certainty with Okii et al. (2014) and Rana et al. (2015) who found negative and significant correlations between days to flowering and number of seeds per pod, with grains yielded. That means the early maturity landraces have a good productivity. Similar results have been observed in Turkey (Madakbas and Ergin, 2011) and in Albania (Bode et al., 2013) but the correlative variables are not identical.
Quantitative analysis regrouped the studied accessions in 4 clusters which facilitates the selection of diverse parents for common beans breeding programs. The comparison of clusters showed that Cluster 1 and 3 contains the best accessions which must be recommended to the farmers in central region of Benin. However, these accessions have presented the highest number of pods per plant but pods length of these clusters is lower than general mean (7.65 cm). It is therefore very important to set up a breeding programme taking into account the accessions of these clusters and the ones of cluster 2 which have the longest pods (25.44 cm). Agronomic tests in other region of the country are also of great importance to compare the performances of these accessions collected in central Benin. Further studies in north and south Benin are recommended to collect and characterize all landraces cultivated in Benin.
CONCLUSION
This study points out the diversity and agronomic performances within common bean in the central region of Benin Republic. This characterization allowed us to group the landraces cultivated in this region in 8 morphotypes according to the seed traits. Regarding qualitative variables of IBPGR descriptors, the collected accessions have been classified in 9 morphotypes while the quantitative variables regrouped the accessions in 4 clusters. Common bean accessions of Cluster 1 and 3 have shown some good agronomical characteristics and could be recommended to farmers. However, breeding programs could be initiated to ameliorate some agronomic traits of common bean landraces of these clusters for the happiness of farmers and consumers. The difference between the number of clusters generated by qualitative variables on one hand and quantitative data on the other hand has revealed that environmental conditions influences morphological characterization.
Molecular characterization is thus important for evaluating common bean diversity cultivated in central Benin. The study should also be widened to other regions of the country.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
The authors are grateful to farmers who gave their common beans seeds for this study along with anonymous reviewers for their comments on previous versions of the manuscript.
REFERENCES
|
Akhshi N, Cheghamirza K, Ahmadi H, Firouzabadi FN (2014). Generation mean analysis to estimate genetic parameters for morphological traits in common bean (Phaseolus vulgaris L.). Journal of Biodiversity and Environmental Sciences 4(4):254-261. |
|
|
Akoègninou A, Van der Burg WJ, Van der Maesen LJG (2006). Flore Analytique du Bénin. Backhuys Publishers, Leiden. |
|
|
Asfaw A, Blair MW (2014). Quantification of drought tolerance in Ethiopian common bean varieties. Agricultural Sciences 5(2):124-139. |
|
|
Balkaya A, Karaagac O (2005). Vegetable Genetic Resources of Turkey. Journal of Vegetation Science 11(4):81-102. |
|
|
Balkaya A, Özbakir M, Kurtar ES (2010). The phenotypic diversity and fruit characterization of winter squash (Cucurbita maxima) populations from the Black Sea Region of Turkey. African Journal of Biotechnology 9(2):152-162. |
|
|
Beebe SE, Rao IM, Blair MW, Acosta-Gallegos JA (2013). Phenotyping common beans for adaptation to drought. In: Ribaut JM, Monneveux P (eds) Drought Phenotyping in Crops: from theory to practice. Generation Challenge Program Special issue on Phenotyping. Physiology 4(35): 311-334. |
|
|
Bitocchi E, Nanni L, Bellucci E, Rossi M, Giardini A, Zeuli PS, Logozzo G, Stougaard J, Mcclean P, Attene G, Papa R (2012). Mesoamerican origin of the common bean (Phaseolus vulgaris L.) is revealed by sequence data. Proceeding of National Academy of Sciences 109(14):E788-E796. |
|
|
Blair MW, Diaz LM, Buendía HF, Duque MC (2009). Genetic diversity, seed size associations and population structure of a core collection of common beans (Phaseolus vulgaris L.). Theoretical and Applied Genetics 119(6):955-972. |
|
|
Bode D, Elezi F, Gixhari B (2013). Morphological characterisation and interrelationships among descriptors in Phaseolus vulgaris accessions. Agriculture and Forestry 59(2):175-185. |
|
|
Broughton WJ, Hernandez G, Blair MW, Beebe SE, Gepts P, Vander-leyden J (2003). Beans (Phaseolus spp)-model food legumes. Plant and Soil 252(1):55-128. |
|
|
Chiorato AF, Carbonell SAM, Dias LADS, Moura RR, Chiavegato MB, Colombo CA (2006). Identification of common bean (Phaseolus vulgaris) duplicates using agromorphological and molecular data. Genetics and Molecular Biology 29:105-111. |
|
|
Coelho RC, Faria MA, Rocha J, Reis A, Oliveira MBPP, Nunes E (2009). Assessing genetic variability in germplasm of Phaseolus vulgaris L collected in Northern Portugal. Scientia Horticulturae 122(3):333-338. |
|
|
De Luque R, José J, Bernardo C (2014). Principal constraints and trends for common bean production and commercialization; establishing priorities for future research. Agronomia Colombiana 32(3):423-431. |
|
|
Doumbia IZ, Akromah R, Asibuo JY (2013). Comparative study of cowpea germplasms diversity from Ghana and Mali using morphological characteristics. Journal of Plant Breeding Genetics 1(3):139-147. |
|
|
Food and Agricultural Organization (FAO) (2014). Food and Agricultural Organization of the United Nations http://www.fao.org/news/archive/news-by-date/2014/en/ |
|
|
Ferreira JJ, Alvarez E, Fueyo MA, Roca A, Giraldez R (2000). Determination of the outcrossing rate of Phaseolus vulgaris L. using protein markers. Euphytica 113(3):259-263. |
|
|
Garcia EH, Valdivia CB, Aguirre JR, Muruaga JS (1997). Morphological and agronomic traits of a wild population and an improved cultivar of common bean (Phaseolus vulgaris L.). Annals of Botany 79(2):207-213. |
|
|
Gomez OJ, Blair MW, Frankow-Lindberg BE, Gullberg U (2004). Molecular and phenotypic diversity of common bean landraces from Nicaragua. Crop Science 44:1412-1418. |
|
|
Hegay S, Geleta M, Bryngelsson T, Asanaliev A, Garkava-Gustavsson L, Hovmalm HP, Ortiz R (2013). Genetic diversity analysis in Phaseolus vulgaris L. using morphological traits. Genetic Resources and Crop Evolution 61(3):555-566. |
|
|
Hegay S, Geleta M, Bryngelsson T, Gustavsson L, Persson-Hovmalm H, Ortiz R (2012). Comparing genetic diversity and population structure of common beans grown in Kyrgyzstan using microsatellites. Scientific Journal of Crop Science 1(4):63-75. |
|
|
Hinkossa A, Gebeyehu S, Zelleke H (2013). Differential effects of post-flowering drought stress on Growth and yield of the basic generations of two common bean (Phaseolus Vulgaris L.). Science, Technology and Arts Research Journal 2(1):22-31. |
|
|
International Board for Plant Genetic Resources (IBPGR) (1982). Phaseolus vulgaris descriptor list. Secretariat, Rome, Italy. |
|
|
Karaagac O, Balkaya A (2013). Interspeciï¬c hybridization and hybrid seed yield of winter squash (Cucurbita maxima Duch) and pumpkin (Cucurbita moschata Duch) lines for rootstock breeding. Scientia Horticulturae 149:9-12. |
|
|
Kumar A, Singh PK, Rai N, Bhaskar GP, Datta D (2014). Genetic diversity of French bean (Phaseolus vulgaris L.) genotypes on the basis of morphological traits and molecular markers. Indian Journal of Biotechnology 13(2):207-213. |
|
|
Loko LEY, Toffa J, Adjatin A, Akpo AJ, Orobiyi A, Dansi A (2018). Folk taxonomy and traditional uses of common bean (Phaseolus vulgaris L.) landraces by the sociolinguistic groups in the central region of the Republic of Benin. Journal of Ethnobiology and Ethnomedicine 14:52. |
|
|
Madakbas SY, Ergin M (2011). Morphological and phenological characterization of Turkish bean (Phaseolus vulgaris L.) genotypes and their present variation states. African Journal of Agricultural Research 6(28):6155-6166. |
|
|
Martins SR, Vences FJ, Miera E, Barroso MR, Carnide V (2006). RAPD analysis of genetic diversity among and within Portuguese landraces of common white bean (Phaseolus vulgaris L.). Scientia Horticulturae 108(3):133-142. |
|
|
Meza N, Rosas JC, Martin JP, Ortiz JM (2013). Biodiversity of common bean (Phaseolus vulgaris L.) in Honduras, evidenced by morphological characterization. Genetic Resources and Crop Evolution 60(4):1329-1336. |
|
|
Miklas PN, Kelly JD, Beebe SE, Blair MW (2006). Common bean breeding for resistance against biotic and abiotic stresses: From classical to MAS breeding. Euphytica 147:105-131. |
|
|
Missihoun AA, Milognon HW, Montcho D, Agbo RI, Sedah P, Agbangla C (2017). Diversité variétale et gestion paysanne des haricots cultivés du genre Phaseolus cultivés au Centre et au Sud Bénin (en Afrique de l'Ouest), Journal of Applied Biosciences 118:1817-11828. |
|
|
Mohammed MS, Shimelis HA, Laing MD (2016). Phenotypic characterization of diverse Bambara groundnut (Vigna subterranea [L.] Verdc) germplasm collections through seed morphology. Genetic Resources and Crop Evolution 63(5):889-899. |
|
|
Okii D, Tukamuhabwa P, Odong T, Namayanja A, Mukabaranga J, Paparu P, Gepts P (2014). Morphological diversity of tropical common bean germplasm. African Crop Science Journal 22(1):59-67. |
|
|
Polania JA, Poschenrieder C, Beebe S, Rao IM (2016). Effective use of water and increased dry matter partitioned to grain contribute to yield of common bean improved for drought resistance. Frontiers in Plant Science 7(660):1-10. |
|
|
Rana JC, Sharma TR, Tyagi RK, Chahota RK, Gautam NK, Singh M Sharma PN, Ojha SN (2015). Characterisation of 4274 accessions of common bean (Phaseolus vulgaris L.) germplasm conserved in the Indian gene bank for phenological, morphological and agricultural traits. Euphytica 205(2):441-457. |
|
|
Rohlf FJ (2000). NTSYS-pc version 2.2: numerical taxonomy and multivariate analysis system. Exeter Software, Setauket, New York. |
|
|
Saba I, Sofi PA, Zeerak NA, Bhat MA, Mir RR (2016). characterisation of a core set of common bean (Phaseolus vulgaris L.) germplasm for seed quality traits. SABRAO Journal of Breeding and Genetics 48(3):359-376. |
|
|
Singh SP, Gepts P, Debouck DG (1991). Races of common bean (Phaseolus vulgaris, Fabaceae). Economic Botany 45:379-396. |
|
|
Sneath PHA, Sokal RR (1973). Numerical taxonomy-the principles and practice of numerical classification. Freeman, San Francisco. |
|
|
Stoilova T, Pereira G, De-sousa MT (2013). Morphological characterization of a small common bean (Phaseolus vulgaris L.) collection under different environments. Journal of Central European Agriculture 14(3):1-11. |
|
|
Stoilova T, Pereira G, Souso de MMT, Carnide V (2005). Diversity in common bean landraces (Phaseolus vulgaris L.) from Bulgaria and Portugal. Journal of Central European Agriculture 6(4):443-448. |
|
|
Ulukapi K, Onus AN (2014). Phenotypic evaluation of some Turkish green bean (Phaseolus Vulgaris L.) Genotypes. Pakistan Journal of Botany 46(4):1415-1420. |
|
|
Welch RM, House WA, Beebe S, Cheng Z (2000). Genetic selection for enhanced bioavailable levels of iron in bean (Phaseolus vulgaris L.) seeds. Journal of Agricultural and Food Chemistry 48(8):3576-3580. |
|
|
Yabi I, Afouda F (2012). Extreme rainfall years in Benin (West Africa). Quaternary International 262(7):39-43. |
|
|
Yaqoob M, Hollington PA, Mahar BA, Gurmani ZA (2013). Yield performance and responses studies of chickpea (Cicer arietinum L.) genotypes under drought stress. Emirates Journal of Food and Agriculture 25(2):117-123. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


