Full Length Research Paper

ABSTRACT
Malaria showed a significant decline in Senegal following the scale up of malaria control around the country with epidemiological disparities. Assessment of multiplicity of infection (MOI) and genetic diversity of Plasmodium falciparum population evolution proved its efficacy in monitoring malaria control. However, to our knowledge few studies focused on the dynamic changes of genetic diversity and MOI in Senegal. The present study aimed to analyze allelic diversity of Pfmsp1 and Pfmsp2 genes and MOI on P. falciparum isolates from Kedougou and Thies regions in Senegal from 2014 to 2017. Allelic polymorphism of Pfmsp1 and Pfmsp2 were assessed by the nested PCR. Over the four years, MAD20 frequencies were significantly predominant in Kedougou (48 to 64%) than Thies (7 to 29%) (P<0.01). Monoclonal infections with K1 (56 to 70%) or RO33 (10 to 17%) were higher in Thies than Kedougou. No significant difference in frequencies of Pfmsp2 allelic families was found and IC3D7 was predominant in both areas during the study. MOI means in the two regions were similar, showing an increase between 2014 and 2015 and a decrease from 2015 to 2017. However, its averages were higher in Kedougou than Thies. The frequencies of the allelic families of Pfmsp1 and Pfmsp2 reported by the study showed a difference within the temporal and spatial evolution of P. falciparum genetic diversity from these regions between 2014 and 2017. However, the similarity observed in the MOI means’ evolution within two regions needs to be deeply investigated on malaria transmission surveillance.
Key words: Senegal, Plasmodium falciparum, Pfmsp1, Pfmsp2, allelic diversity, MOI.
Abbreviation: DBS, Dried blood samples; DNA, deoxyribonucleic acid; EIR, entomological inoculation rate; MOI, multiplicity of infection; NMCP, National Malaria Control Program; PCR, polymerase chain reaction; Pfmsp1, merozoite surface protein 1 of Plasmodium falciparum; Pfmsp2, merozoite surface protein 2 of Plasmodium falciparum; SNP, single nucleotide polymorphism; UV, ultraviolet; WHO, World Health Organization.
INTRODUCTION
Malaria remains a public health burden threat as it reached 228 million cases and was responsible for 405 000 deaths worldwide in 2018 (WHO, 2019). Therefore, over fifteen years, malaria controls were intensified in many endemic areas including Senegal (PNLP, 2016, 2017). On the other hand, scale up of malaria control may cause changes in malaria epidemiology particularly on the intensity of malaria transmission and genetic diversity of parasite population (Branch et al., 2011; Nkhoma et al., 2013; Daniels et al., 2015; Obaldia et al., 2015). Indeed, malaria transmission pattern can affect the multiplicity of infection (MOI) and genetic diversity of parasite populations (Su et al., 2009). Hence, the usefulness of genetic diversity and MOI assessment which may provide insight on malaria transmission (Daniels et al., 2015).
To date, molecular approaches showed convincing results to malaria control and elimination in numerous countries where malaria transmission is decreasing such as Senegal (Daniels et al., 2013, 2015; Niang et al., 2017), Panama (Obaldia et al., 2015) and Columbia (Chenet et al., 2012). Among these molecular approaches, genotyping of allelic families of Plasmodium falciparum merozoite surface protein 1 (Pfmsp1) and 2 (Pfmsp2) genes is a standard method for assessing the MOI (Atroosh et al., 2011; Mueller et al., 2012) and exhibit efficiency in gauging different genotypes of parasite co-infections. These two polymorphic antigenic markers are also useful in characterizing P. falciparum populations in malaria endemic areas (Salem et al., 2014; Kateera et al., 2016; Yavo et al., 2016; Niang et al., 2017; Chen et al., 2018; Metoh et al., 2020).
Following the intensification of malaria control in Senegal, its transmission revealed some disparities within the country (PNLP, 2016). Actually, the Southern part of the country shows high malaria transmission with annual incidence greater than 100/1000 inhabitants while the Northern part displays low to moderate transmission with an annual incidence rates less than 5/1000 inhabitants (PNLP, 2015). However, a spatio-temporal analysis of complexity of P. falciparum infections realized in Kedougou (a Southern region) suggested a decrease of malaria transmission from 2009 to 2013 (Niang et al., 2017). Moreover, in Thies (a Western region) the application of 24 Single Nucleotide Polymorphism (SNP) genomic barcodes for malaria surveillance combined with incidence data from the national malaria control policies (NMCP) also reported a reduction in malaria transmission from 2006 to 2010 but with a significant rebound from 2012 to 2013 (Daniels et al., 2015). These two reports raised some contradictions particularly the rise of genetic diversity of parasites in Thies region known as a low malaria transmission area (Daniels et al., 2015; Niang et al., 2017). Therefore, additional studies are needed beyond 2013 in both Thies and Kedougou to better characterize the temporal and spatial evolution of MOI and P. falciparum genetic diversity. This present study aimed to analyze the allelic diversity of Pfmsp1 and Pfmsp2 genes and MOI of P. falciparum isolates from Kedougou and Thies from 2014 to 2017.
MATERIALS AND METHODS
Ethics approval and consent to participate
The study was approved by the National Ethics Committee for Health Research of Senegal. Before patients were enrolled in the study sites, benefits and any perceived risks were explained to all participants in French or local languages. Informed consent for adults or guardians consent for children less than 18 years old were obtained before participant recruitment and sample collection.
Study sites and period
This is an observational surveillance study carried out in febrile patients visiting healthcare facilities in Kedougou and Thies over a four-year period (2014-2017), during malaria transmission season (Figure 1). Kedougou is an administrative region located in the South-East of Senegal at 685 km from Dakar (the capital city). There, malaria transmission is high and seasonal from July to December; the entomological inoculation rate (EIR) ranges from 20 to 100 infectious bites per person and per year and malaria incidence is higher than 15 malaria cases per 1,000 habitants (PNLP, 2015, 2016). Thies is also an administrative region located at the west of the country at 70 km from Dakar. Malaria transmission in Thies is low with EIR varying from 0 to 20 infections bites per person/year and malaria incidence varies from 5 to 15 malaria cases per 1,000 habitants. The malaria transmission seasonality in this area overlaps with the rainy season that takes place between September and December (PNLP, 2015, 2016).
Sample collection
Three hundred and eighty (380) dried blood spots (DBS) were collected on filter paper. Patients met the following criteria: living in a 15-km radius of health facilities, having fever (axillary temperature ≥ 37.5°C) or history of fever in the previous 48 h. Age ranging from 6 months to 75 years and uncomplicated P. falciparum malaria with parasite density ≥ 1000 asexual forms per microliter. Patients who presented signs of severe malaria as defined by World Health Organization (WHO) and pregnant women were not included (WHO, 2000).
DNA extraction and PCR genotyping
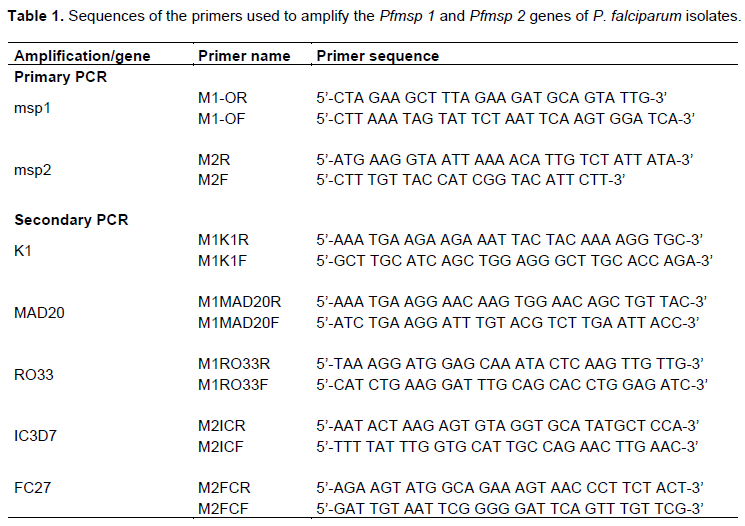
Parasite deoxyribonucleic acid (DNA) was extracted from filter papers using QIAamp DNA Mini kit (Qiagen®) following the manufacturer's instructions. The polymorphic loci of Pfmsp1 block 2 (K1, MAD20 and RO33 allelic families) and Pfmsp2 central region (IC3D7 and FC27 allelic families) were amplified by nested PCR as described in previous studies (Snounou et al., 1999; Mayengue et al., 2007) with primers described in Table 1 (Ndiaye et al., 2019). All polymerase chain reactions (PCR) were performed in a total volume of 25 μl containing: 4 μl PCR master mix (0.2 U/μl Taq Polymerase, 10 mM deoxyribonucleotide triphosphate), and 10 μM of each forward and reverse primer for each gene. In the first round reaction (nest 1), 2 μl of genomic DNA was added as a template. In the second nested reaction (nest 2), 1 μl of the nest 1 PCR product was used as a DNA template. Cycling conditions for primary PCR were as follows: initial denaturation at 95°C for 5 min, followed by 35 cycles of denaturation 94°C for 1 min, annealing at 58°C for 2 min and extension 72°C for 2 min; and a final extension was done at 72°C for 3 min. The cycling conditions for nested PCR were: initial denaturation at 95°C for 5 min. That followed by 35 cycles of denaturation 94°C for 1 min. Annealing 61°C for 2 min, extension 72°C for 2 min, with a final extension cycle of 72°C for 3 min. Positive (3D7 and Dd2) and negative (reagent grade water) controls were systematically incorporated in each PCR run. The nested PCR products were revealed by electrophoresis on 2% agarose gels stained with ethidium bromide and visualized under UV trans-illumination (VersaDoc®, BIORAD, Hercules, USA). The sizes of nested PCR products were estimated using 100 bp DNA ladder (Promega, Madison, USA).
Statistical analysis
Isolates with two or more alleles were considered as multiclonal infections and samples with one allele as monoclonal infection. The multiplicity of infection (MOI) was calculated by dividing the total number of alleles detected for Pfmsp1 and Pfmsp2 genes by the total number of samples (Mayengue et al., 2007). Chi2 test for trend was used to compare the allelic families and multiclonal isolates frequencies over the years and Student’s t test from online Biostatgv was used to compare MOI over the time. For all statistical tests, the significance level was stated if p< 0.05.
RESULTS
In total, genetic material was obtained from 380 enrolled patients from the two localities: 200 in Kedougou and 180 in Thies (Table 2). From these samples, 96% (365/380) for Pfmsp1 gene and 94% (356/380) for Pfmsp2 gene were successfully amplified.

Genetic polymorphism of Pfmsp1 gene in Kedougou and Thies
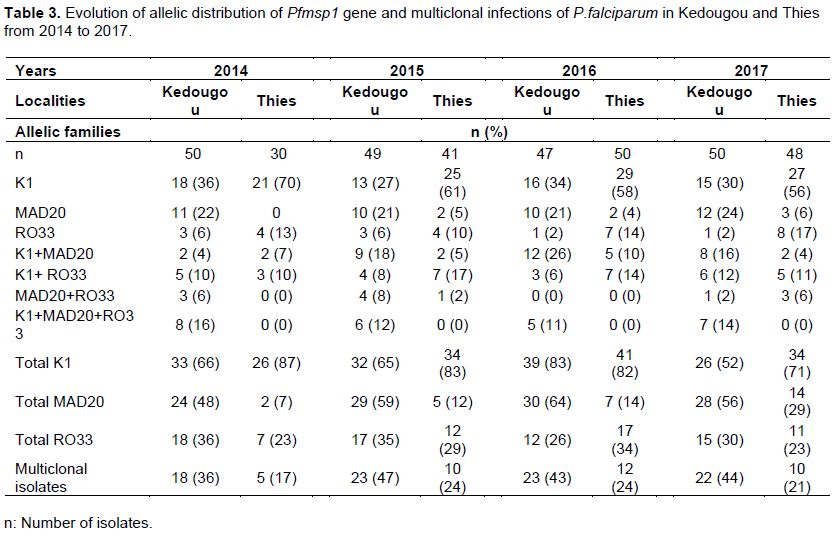
K1, MAD20 and RO33 allelic families were observed in Kedougou and Thies. The frequencies of these three allelic families, their dimorphic combinations (K1/MAD20, K1/RO33 and MAD20/RO33) as well as trimorphic combination (K1/MAD20/RO33) were described in Table 3. In Kedougou, the highest frequencies of both K1 and MAD20 allelic families were observed in 2016, 83% (39/47) and 64% (30/47), respectively. In Thies, K1 and RO33 allelic families were predominant with frequencies varying from 71% (34/48) to 87% (26/30) and from 23% (7/30) to 34% (17/50), respectively between 2014 and 2017. The K1 allelic family was significantly predominant in Thies than Kedougou (P<0.01) while MAD20 allelic family was significantly present in Kedougou than in Thies during the study period (P<0.01). Monoclonal infections with K1 allelic family were significantly higher in Thies than in Kedougou (P<0.01) with decreasing frequencies from 70% (21/30) in 2014 to 56% (27/48) in 2017. Monoclonal infections due to MAD20 allelic family were significantly higher in Kedougou than in Thies (P<0.01) and showed an irregular trend over the four years (Table 3). Frequencies of multiclonal isolates were significantly higher in Kedougou than in Thies (P<0.01) from 2014 to 2017. Dimorphic Pfmsp1 allelic combination K1/MAD20 was significantly present in Kedougou than Thies (P=0.01). Infections due to K1/RO33 and MAD20/RO33 associations showed no linear trend in Kedougou as well as in Thies between 2014 and 2017. Samples carrying the trimorphic combination were identified only in Kedougou with frequencies ranging from 12 to 16% during the four-year study period (Table 3).
Genetic polymorphism of Pfmsp2 gene in Kedougou and Thies
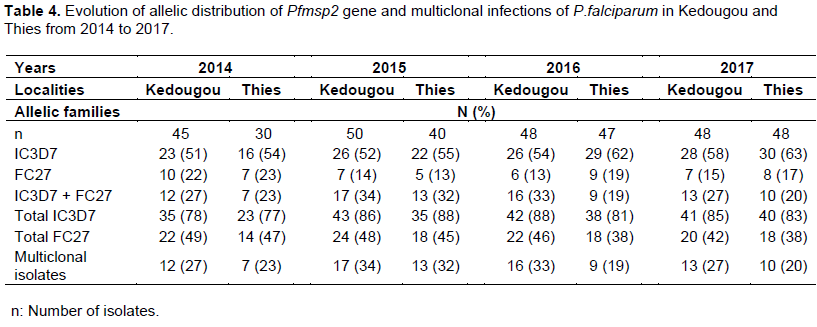
IC3D7 and FC27 allelic families were identified in Kedougou and Thies from 2014 to 2017. The frequencies of allelic families and their combinations were illustrated in Table 4. The evolution of allelic families of Pfmsp2 gene was almost similar in Kedougou and Thies all over the study period and no significant difference on allelic family’s distribution was found between the two areas. The IC3D7 allelic family remained the most represented allelic family and the highest frequencies (88%) were observed in 2015, in Kedougou as well as in Thies. In Kedougou, monoclonal infections with IC3D7 were the most predominant with increasing frequency from 51% (23/45) in 2014 to 58% (28/48) in 2017 while these increased from 53% (16/30) to 63% (30/48) in Thies. Frequencies of multiclonal isolates were always higher in Kedougou than in Thies.
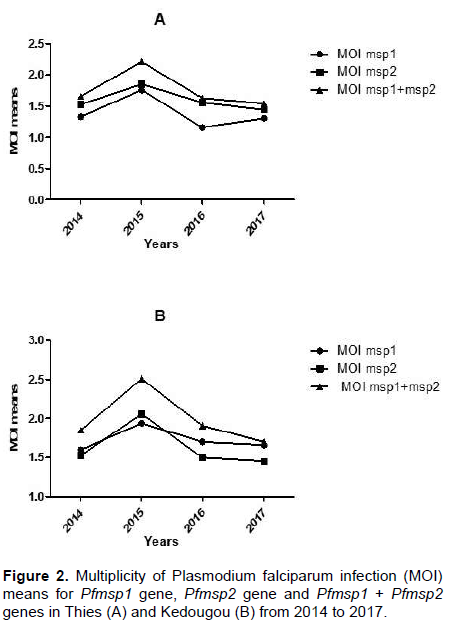
Multiplicity of P. falciparum infection in Kedougou and Thies from 2014 to 2017
MOI evolution in Kedougou and Thies over the 4-year of study are as shown in Figure 2. The number of Pfmsp1 and Pfmsp2 alleles per specimen ranged from 1 to 5. The same trends of MOI evolution were observed in both regions from 2014 to 2017; MOI means increased between 2014 and 2015 and decreased progressively from 2015 to 2017. However, the MOI means for both Pfmsp1 and Pfmsp2 found in Kedougou were higher than those found in Thies (1.85 versus 1.66 in 2014, 2.51 vs. 2.21 in 2015, 1.91 vs. 1.63 in 2016 and 1.7 vs. 1.54 in 2017). Overall, MOI means varied between both localities and showed a significant difference in 2017 only (P<0.01). The MOI means of Pfmsp1 loci were significantly more important in Kedougou than in Thies with 1.7 vs. 1.16 (P<0.01) and 1.66 vs. 1.31 (P=0.031), respectively in 2016 and 2017. The MOI means for Pfmsp2 gene followed the general MOI distribution but no significant difference was observed between Kedougou and Thies.
DISCUSSION
This study reports analysis temporal evolution of allelic polymorphism of Pfmsp1 and Pfmsp2 genes and multiplicity of P. falciparum infections across Kedougou and Thies, two area in Senegal with different malaria transmission intensities during four years from 2014 to 2017. The frequencies of the allelic families of Pfmsp1 and Pfmsp2 showed a difference within the temporal and spatial evolution of P. falciparum genetic diversity and a similarity in the MOI means’ evolution in these regions over the four years (2014-2017).
The predominance of the K1 allelic family in the Pfmsp1 gene reported in this study suggests the adaptability of these strains (or alleles) in these malaria endemic areas. Similar results were reported by Yavo et al. (2016) in sub-Saharan Africa particularly in Gabon and Ivory-Cost, underlining this strong selection of the K1 allelic family within Pfmsp1 with frequencies of 64.6 and 56.6% in Ivory Coast and Gabon, respectively. Analogous findings were also observed in Mauritania with a prevalence of 90% of K1 allelic family (Salem et al., 2014) as well as in Yemen where 58% of isolates harbored this allelic family (Al-abd et al., 2013). By contrast, a previous study carried out in Pahang, Malaysia reported predominance of RO33 allelic family with frequency of 80% while K1 was less represented (Atroosh et al., 2011). These findings about allelic polymorphism of Pfmsp1 gene could show different distributions in various geographical locations in malaria endemic areas (Mwingira et al., 2011). This observation on the difference on allelic diversity distribution of Pfmsp1 between different geographic regions has been reported between the Southeast regions and one Western state of Myanmar (Soe et al., 2017). Accordingly, we found some significant associations between allelic diversity of Pfmsp1 and endemicity of areas. The high prevalence of MAD20 allelic family in the parasite population, Southeast of Senegal was previously in 2013 (Niang et al., 2017). These authors showed that MAD20 allelic family was associated with the highest parasitaemia (Agyeman-Budu et al., 2013; Niang et al., 2017) and could explain its strong presence in this area where malaria transmission remains active over the time (PNLP, 2016). For Pfmsp2, overall, there was a lack of major differences of distribution of allelic families; IC3D7 was predominant in Kedougou as well as in Thies between 2014 and 2017. Previous studies carried out in Kedougou (Niang et al., 2017) and in Ghana in 2013 showed that IC3D7 strains were about 4, outnumbered the FC27 strains throughout the year (Agyeman-Budu et al., 2013). It is known that FC27 allelic type has been shown to be more prevalent in asymptomatic rather than symptomatic Plasmodium carrier individuals (Oyedeji et al., 2013), suggesting a lower risk to find a high prevalence of this allelic family in studies carried out on symptomatic patients in the field (A-Elbasit et al., 2007; Amodu et al., 2008).
Overall, allelic diversity of the Pfmsp1 gene appears to be more affected by malaria transmission intensity because in this study, we found significant differences in allelic distribution between the two studies areas over time and this was not observed for the Pfmsp2 gene. Indeed, it was reported that Pfmsp1 gene is more likely to be involved in malaria symptomatology by participating in erythrocyte invasion and immune evasion (Ogutu et al., 2009; Moss et al., 2012) and this situation could explained the difference found in areas with different malaria transmission level.
The higher predominance of multiclonal isolates are in Kedougou (high malaria transmission area) compared to Thies (low malaria transmission area). This was consistent with previous studies which identified a high number of multiclonal isolates in high malaria transmission areas (Yavo et al., 2016; Niang et al., 2017) suggesting that clonality of infection could increase with higher malaria transmission intensity (Anderson et al., 2000). The relatively higher MOI means for msp genes (Pfmsp1 and Pfmsp2) observed in Kedougou is consistent with a previous study comparing the clonality of malaria infection in two areas with different malaria endemicity (Grande Comore, meso-hyperendemic and Thies, hypoendemic area in Senegal) (Papa Mze et al., 2015).
However, previous studies stipulated that high MOI is common to malaria hyperendemic areas (Ghanchi et al., 2010; Mawili-Mboumba et al., 2015) and directly linked with malaria transmission intensity (Anderson et al., 2000; Agyeman-Budu et al., 2013). This suggests to reconsider the endemicity level in Thies (reported as hypoendemic area), given the rapid and dramatic increase of malaria incidence since 2012 in this region. The similarity of MOI evolution between Kedougou and Thies is probably due to the high rainfall recorded in this country during this year (574.5 mm in Thies and 1576.7 mm in Kedougou) compared to other years of studies (PNLP, 2017).
These results are consistent with the trend evolution of malaria morbidity and mortality in Senegal between 2014 and 2017 (PNLP, 2016, 2017) suggesting a uniform coverage of malaria controls such as the mass distribution of insecticide-treated bednets and rapid malaria diagnosis with appropriate treatment (PNLP, 2016) in the country. These overall results found in these both localities with different endemicities could insinuate an efficiency of intensive intervention to control malaria, resulting in slow decline of malaria transmission in these regions and continual surveillance of MOI could be a good monitor tool of malaria transmission.
As shown, Pfmsp1 and Pfmsp2 genes are robust polymorphism markers and can be used successfully to characterize genetic P. falciparum strains populations (Salem et al., 2014; Kateera et al., 2016; Yavo et al., 2016; Niang et al., 2017; Chen et al., 2018; Metoh et al., 2020). However, there is a limitation as the challenge to distinguish allelic variants presenting similar fragment size during agarose gel electrophoresis migration could underestimate the P. falciparum genetic diversity (Cattamanchi et al., 2003; Mwingira et al., 2011).
CONCLUSION
The genetic diversity of P. falciparum parasites and MOI can impact malaria transmission and malaria control strategies. Therefore, it is important to monitor the genetic diversity of parasites in endemic areas such as Kedougou and Thies. Indeed, the frequencies of the allelic families of Pfmsp1 and Pfmsp2 reported by our study showed a difference within the temporal and spatial evolution of P. falciparum genetic diversity from Kedougou to Thies regions of Senegal between 2014 and 2017. Our findings also showed similarity of MOI evolution from isolates collected in Kedougou and Thies, respectively in a high and a low malaria transmission region from 2014 to 2017. This underlines a uniform efficiency strategy introduced by NMCP within Senegal since 2006. However, the high MOI revealed by our study indicated that malaria transmission remains high in both areas despite the massive scale-up of malaria interventions. Thus, to achieve malaria elimination, the NMCP should make a continual surveillance of MOI which is a robust malaria transmission indicator, in sentinels’ sites in order to monitor the trend of malaria transmission within Senegal. Therefore, additional studies including parasites population dynamics, MOI estimation and immune status of P. falciparum-infected individuals from isolates collected in different regions of Senegal could more elucidate malaria transmission dynamics of P. falciparum in Senegal as well as to be useful to guide malaria interventions.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
The authors acknowledge all participants of the study in Senegal. They thank Younouss Diedhiou, Lamine Ndiaye, Amadou Mactar Mbaye and staff of Cheikh Anta Diop University for their contribution in this study. The work was supported by Cheikh Anta Diop University and the International Centers of Excellence for Malaria Research (ICEMR) West Africa (U19AI089696).
REFERENCES
|
A-Elbasit IE, ElGhazali G, TM AE, Hamad AA, Babiker HA, Elbashir MI, Gih HA (2007). Allelic polymorphism of MSP2 gene in severe P. falciparum malaria in an area of low and seasonal transmission. Parasitology Research 102(1):29-34 |
|
|
Agyeman-Budu A, Brown C, Adjei G, Adams M, Dosoo D, Dery D, Wilson M, Asante KP, Greenwood B, Owusu-Agyei S (2013). Trends in multiplicity of Plasmodium falciparum infections among asymptomatic residents in the middle belt of Ghana. Malaria Journal 12:22. |
|
|
Al-abd NM, Mahdy MAK, Al-Mekhlafi AMQ, Snounou G, Abdul-Majid NB, Al-Mekhlafi HM, Fong MY (2013). The Suitability of P. falciparum Merozoite Surface Proteins 1 and 2 as Genetic Markers for In Vivo Drug Trials in Yemen. Plos ONE 7:e67853. |
|
|
Amodu OK, Oyedeji SI, Ntoumi F, Orimadegun AE, Gbadegesin RA, Olumese PE, Omotade OO (2008). Complexity of the msp2 locus and the severity of childhood malaria, in south-western Nigeria. Annals of Tropical Medicine and Parasitology 102(2):95-102. |
|
|
Anderson TJ, Haubold B, Williams JT, Estrada-Franco JG, Richardson L, Mollinedo R, Bockarie M, Mokili J, Mharakurwa S, French N, Whitworth J, Velez ID, Brockman AH, Nosten F, Ferreira MU, Day KP (2000). Microsatellite markers reveal a spectrum of population structures in the malaria parasite Plasmodium falciparum. Molecular Biology and Evolution17(10):1467-1482. |
|
|
Atroosh WM, Al-Mekhlafi HM, Mahdy MA, Saif-Ali R, Al-Mekhlafi AM, Surin J (2011). Genetic diversity of Plasmodium falciparum isolates from Pahang, Malaysia based on MSP-1 and MSP-2 genes. Parasites and Vectors 4:233. |
|
|
Branch OH, Sutton PL, Barnes C, Castro JC, Hussin J, Awadalla P, HijarG (2011). Plasmodium falciparum genetic diversity maintained and amplified over 5 years of a low transmission endemic in the Peruvian Amazon. Molecular Biology and Evolution 28(7):1973-1986. |
|
|
Cattamanchi A, Kyabayinze D, Hubbard A, Rosenthal PJ, Dorsey G (2003).Distinguishing recrudescence from reinfection in a longitudinal antimalarial drug efficacy study: comparison of results based on genotyping of msp-1, msp-2, and glurp. American Society of Tropical Medicine and Hygiene 68:133-139. |
|
|
Chen JT, Li J, Zha GC, Huang G, Huang ZX, Xie DD, Zhou X, Mo HT, Eyi JUM, Matesa RA, Obono MMO, Li S, Liu XZ, Lin M (2018). Genetic diversity and allele frequencies of Plasmodium falciparum msp1 and msp2 in parasite isolates from Bioko Island, Equatorial Guinea. Malaria Journal 17:458. |
|
|
Chenet SM, Schneider KA, Villegas L, Escalante AA (2012). Local population structure of Plasmodium: impact on malaria control and elimination. Malaria Journal 11:412. |
|
|
Daniels RF, Chang HH, Séne PD, Park DC, Neafsey DE, Schaffner SF, Hamilton EJ, Lukens AK, Tyne DV, Mboup S, Sabeti PC, Ndiaye D, Wirth DF, Hartl DL, Volkman SK (2013).Genetic surveillance detects both clonal and epidemic transmission of malaria following enhanced intervention in Senegal. PLoS ONE 8:60780. |
|
|
Daniels RF, Schaffnerc SF, Wengerd EA, Proctord JL, Change HH, Wong W, Baro N, Ndiaye D, Fall FB, Ndiop M, Ba M, Milner Jr DA, Taylor TE, Neafsey DE, VolkmanSK, Eckhoff PA, Hartl DL, Wirth DF (2015). Modeling malaria genomics reveals transmission decline and rebound in Senegal.Proceedings of the National Academy of Sciences of the United States of America 112(22):7067-7072. |
|
|
Ghanchi NK, Martensson A, Ursing J, Jafri S, Bereczky S, Hussain R, Beg MA (2010). Genetic diversity among Plasmodium falciparum field isolates in Pakistan measured with PCR genotyping of the merozoite surface protein 1 and 2. Malaria Journal 9:1. |
|
|
Kateera F,Nsobya SL, Tukwasibwe S,Mens PF, Hakizimana E, Grobusch MP, Mutesa L, Kumar N, Vugt MV (2016). Malaria case clinical profiles and Plasmodium falciparum parasite genetic diversity: a cross sectional survey at two sites of different malaria transmission intensities in Rwanda. Malaria Journal 15:237. |
|
|
Mawili-Mboumba DP, Mbondoukwe N, Adande E, Bouyou-Akotet MK (2015). Allelic Diversity of PFMSP1 Gene in Plasmodium falciparum from Rural and Urban Areas of Gabon. Korean Journal of Parasitology 53(4):413-419. |
|
|
Mayengue PI, Ndounga M, Davy MM, Tandou N, Ntoumi F (2007). In vivo chloroquine resistance and prevalence of the pfcrt codon 76 mutation in Plasmodium falciparum isolates from the republic of Congo. Acta Tropica 95:219-225. |
|
|
Metoh TN, Chen JH, Gah PF, Zhou X, Somo RM, Zhou XN ( 2020). Genetic diversity of Plasmodium falciparum and genetic profile in children affected by uncomplicated malaria in Cameroon. Malaria Journal 19:115. |
|
|
Moss DK, Remarque EJ, Faber BW, Cavanagh DR, Arnot DE, Thomas AW, Holder AA (2012).Plasmodium falciparum 19-kilodalton merozoite surface protein 1 (PFMSP1)-specific antibodies that interfere with parasite growth in vitro can inhibit PFMSP1 processing, merozoite invasion, and intracellular parasite development. Infection and Immunity 80:1280-1287. |
|
|
Mueller I, Schoepflin S, Smith TA, Benton KL, Bretscher MT, Lin E, Kiniboro Benson, Zimmerman PA, Speed TP, Siba P, Felge I (2012). Force of infection is key to understanding the epidemiology of Plasmodium falciparum malaria in Papua New Guinean children. Proceedings of the National Academy of Sciences of the United States of America 109(25):10030-10035. |
|
|
Mwingira F, Nkwengulila G, Schoepflin S, Sumari D, Beck H-P, Snounou G, Felger I, Olliaro P, Mugittu K (2011). Allele frequency and diversity in sub-Saharan Africa. Malaria Journal 10:79. |
|
|
Ndiaye T, Sy M, Gaye A, Ndiaye D (2019). Genetic polymorphism of Merozoite Surface Protein 1 (msp1) and 2 (msp2) genes and multiplicity of Plasmodium falciparum infection across various endemic areas in Senegal. African Health Sciences 19(3):2446-2456. |
|
|
Niang M, Thiam LG, Loucoubar C, Sow A, Sadio BD, Diallo M, Amadou A Sall, Aissatou Toure-Balde (2017). Spatio-temporal analysis of the genetic diversity and complexity of Plasmodium falciparum infections in Kédougou, Southeastern Senegal. Parasites and Vectors 10:33. |
|
|
Nkhoma SC, Nair S, Al-Saai S, Ashley E, McGready R, Phyo AP, Nosten F, Anderson TJC (2013). Population genetic correlates of declining transmission in a human pathogen. Molecular Ecology 22(2):273-285. |
|
|
Obaldia N , Baro NK, Calzada JE, Santamaria AM, Daniels R, Wong W, ChangHH, Hamilton EJ, Arevalo-HerreraM, HerreraS, Wirth DF, Hartl DL, Marti M, Volkman SK (2015). Clonal Outbreak of Plasmodium falciparum Infection in eastern Panama. Journal of Infectious Diseases 211(7):1087-1096. |
|
|
Ogutu BR, Apollo OJ, McKinney D, Okoth W, Siangla J, Dubovsky F, Tucker K, Waitumbi JN, Diggs C, Wittes J, Malkin E, Leach A, Soisson LA, Milman JB, Otieno L, Holland CA, Polhemus M, Remich SA, Ockenhouse C, Cohen J, Ballou WR, Martin SK, Angov E, Stewart VA, Lyon JA, Heppner DG, Withers MR, MSP-1 Malaria Vaccine Working Group (2009). Blood stage malaria vaccine eliciting high antigen-specific antibody concentrations confers no protection to young children in Western Kenya. Plos One 4:e4708. |
|
|
Oyedeji SI, Awobode HO, Anumudu C, Kun J (2013). Genetic diversity of Plasmodium falciparum isolates from naturally infected children in northcentral Nigeria using the merozoite surface protein-2 as molecular marker. Asian Pacific Journal of Tropical Medicine 6(8):589-594. |
|
|
Papa Mze N, Ndiaye YD, Diedhiou CK, Rahamatou S, Dieye B, Daniels RF, Hamilton EJ, Diallo M, Bei AK, Wirth DF, Mboup S, Volkman SK, Ahouidi AD, Ndiaye D (2015). RDTs as a source of DNA to study Plasmodium falciparum drug resistance in isolates from Senegal and the Comoros Islands. Malaria Journal 14:373. |
|
|
Programme National de Lutte contre le Paludisme (PNLP) (2015). Bulletin épidémiologique annuel du paludisme au Senegal. Dakar. |
|
|
Programme National de Lutte contre le Paludisme (PNLP) (2016). Plan stratégique national de lutte contre le paludisme au Sénégal (2016-2020). |
|
|
Programme National de Lutte contre le Paludisme (PNLP) (2017). Bulletin épidémiologique annuel du paludisme au Senegal. |
|
|
Programme National de Lutte contre le Paludisme (PNLP) (2016). Bulletin épidémiologique annuel du paludisme au Senegal. Dakar. |
|
|
Salem MSOA, Ndiaye M, OuldAbdallahi M, Lekweiry KM, Bogreau H, Konaté L, Faye B, Gaye O, Faye O, Boukhary AOMSO (2014). Polymorphism of the merozoite surface protein-1 block 2 region in Plasmodium falciparum isolates from Mauritania. Malaria Journal 13:26. |
|
|
Snounou G, Zhu X, Spiripoon N, Jarra W, Thaithong S, Brown KN, Viriyakosol S (1999). Biased distribution of Pfmsp1 and Pfmsp2 allelic variant in Plasmodiumfalciparum populations in Thailand. Transactions of the Royal Society of Tropical Medicine and Hygiene 93:369-374. |
|
|
Soe TN, Wu Y, Tun MW, Xu X, Hu Y, Ruan Y, Han KT, Aye KM, Morris J, Su P, Yang Z, Kyaw PM, Cui L (2017) . Genetic diversity of Plasmodium falciparum populations in southeast and western Myanmar. Parasites and Vectors 10:322. |
|
|
Su X-Z, Jiang H, Yi M, Mu J, Stephens RM (2009). Large-scale genotyping and genetic mapping in Plasmodium parasites. The Korean Journal of parasitology 47(2):83. |
|
|
World Health Organization (WHO) (2000). Communicable diseases cluster: severe falciparum malaria. Transactions of the Royal Society of Tropical Medicine and Hygiene 94:S1-S9. |
|
|
World Health Organization (WHO) (2019). World malaria report 2019. World Health Organization. |
|
|
Yavo W, Konaté A, Mawili-Mboumba DP, Kassi FK, Tshibola Mbuyi ML, Angora EK, Menan EIH, Bouyou-Akotet MK (2016). Genetic Polymorphism of Pfmsp1 and Pfmsp2 in Plasmodium falciparum Isolates from Côte d'Ivoire versus Gabon. Journal of Parasitology Research 2016:3074803. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


