ABSTRACT
The protective effect of probiotic strain Enterococcus faecium EF55 in chicks challenged with Salmonella enterica serovar Enteritidis 147 (SE147) was assessed in caecum and blood. Quantitative reverse transcription-polymerase chain reaction (RT-PCR) detection of mRNAs of chicken intestinal mucin gene (MUC2), Interleukin (IL-15) and Interleukin (IL-17) cytokines, together with flow cytometry determination of intraepithelial lymphocytes (IEL) were investigated in caecum. White blood cell counts were measured haematologically. One-day-old chicks were divided into 4 groups. The probiotic (EF) and probiotic+salmonella groups (EFSE) received perorally 109 CFU EF55 7 days. The challenged SE and EFSE groups were single perorally infected with 108 CFU SE147 on day 4. No bacteria were given to controls (C). Administration of EF55 to chicks challenged with SE147 (EFSE group) ultimately resulted in increased number of blood heterophils, caecal IgA+ IEL, and decreased caecal expression of MUC compared to SE group on 7 days post infection (d.p.i.). Detection of IL-15 and IL-17 mRNAs cytokine level in caecum showed a tendency to their increase on 1 d.p.i. in EFSE group. The results demonstrated beneficial effect of EF55 on the expression of MUC and production of IgA+ IEL in caecum after SE147 infection. Furthermore, the results suggest certain relation between expression of cytokines, expression of MUC and IgA+ cells.
Key words: blood, chicks, immunity, infection, intestine, probiotics.
Enteric salmonellosis is a major foodborne disease caused by Salmonella and its serovars, the most common being Enteritidis and Typhimurium. Chicken is recognized as one of the most important reservoir of Salmonella. The colonization of the chicken gut by S. enterica could result in the contamination of the environment and food chain. Broiler chicks are most susceptible to colonization by pathogens during the early period of their life, when a stable gut-microflora is not established, and an introduced stabilized gut-microflora can provide them with vital protection against these undesirable organisms (Chambers and Lu, 2002).
Potential probiotic strains have been found among the intestinal microbiota of food producing animals. Among them, some bacteria have been shown to exert antimicrobial activity against enteric pathogens due to the production of bacteriocins and bacteriocin-like molecules. The E. faecium EF55 was shown to produce a bacteriocin-like substance, and the inhibitory activity of this probiotic strain of bacteria was reported against number of enteropathogenic bacteria, including S. enterica (Levkut et al., 2009).
Mucosal tissues represent the route of access and site of infection for the majority of pathogens. Mucosal glycoproteins are secreted in large quantities by mucosal epithelia and cell surface mucins are a prominent feature of the apical glycocalyx of all mucosal epithelia. Expression of cell-surface mucins can be upregulated by inflammatory cytokines and other inflammatory factors (Linden et al., 2008). Similarly, some inflammatory cytokines have been demonstrated to play a critical role in induction and expression of SIgA responses at mucosal surfaces (Husband, 2002). Interleukin-15 was shown to be proinflammatory cytokine produced by mononuclear phagocytes (Lillehoj et al., 2001), activated T cells, and stimulates growth and proliferation of intestinal epithelial cells, natural killer cells and activated B cells (Kaiser et al., 2002). Interleukin-17 is a proinflammatory cytokine produced by activated and memory T cells (Hong et al., 2006). Functional studies in mice indicated that IL-17 is involved into stimulation of granulopoiesis, proliferation and differentiation of cells into neutrophils (Fossiez et al., 1996). In chickens IL-17 showed protective role in local immunity against salmonellae (Crhanova et al., 2011). However, the local immune response in the gastrointestinal tract has been scarcely studied in relation to competitive exclusion. It is likely to involve a variety of factors such as the role of native microflora, secretory IgA, mucin barrier, and products of inflammatory cells.
In order to find a certain relation to the quantity of cells and level of cytokines as well as mucin gene expression involved in natural and acquired intestinal immunity in chicks after protection of a probiotic strain E. faecium EF55 and challenged with S. enterica serovar Enteritidis 147, we chose to study in caecum, the intraepithelial lymphocytes, chicken intestinal mucin gene mRNA expression like human MUC2, as well as proinflammatory cytokines IL-15 and IL-17 mRNA expression together with white blood cell counts in the peripheral blood.
Animals
One-day-old chicks (220) of Cobb-500 hybrids were randomly divided into 4 groups (n=55) and placed in large pens with a floor-covering of wood shaving for 11 days. The bedding was changed daily during the course of experiment. The temperature was based on the requirement for chicks of this age group: 32°C in the first week and reduction of 2°C each successive day. Water and feed (commercial diet BR1, Barbara Comp., ÄŒaňa, Slovak Republic) were available ad libitum. The chicks were kept in the menagerie of the Department of Pathological Anatomy, University of Veterinary Medicine and Pharmacy, Košice, Slovakia (SK P 52004), in accordance with the rules and approval of the Ethics Committee, and the experiment was authorized by the State Veterinary and Food Administration of the Slovak Republic (ÄŒ.k.Ro-270710-221).
Experimental design and samplings
The chicks of group 1 (Control) were fed only with commercial diet without application of any bacteria (negative control). Enterococcus faecium EF 55 in the dose of 109 CFU/ml was administered in 0.2 ml phosphate buffer saline (PBS) to chicks of group 2 (EF) per os daily from 1 to 7 days. The birds of group 3 (SE) were individually inoculated per os on day 4 of the experiment with Salmonella enteritidis SE 147 (provided by Dr. Rychlik from Veterinary Research Institute, Brno, Czech Republic) with a single dose 1x108 CFU/ml in 0.2 ml PBS. The chicks of group 4 (EFSE) were administered with E. faecium EF 55 during the first 7 days in the same manner as those of group 2, and were inoculated with S. Enteritidis on day 4 of the experiment in the same way as the chicks of group 3. Ten chicks from each group were killed and samples were taken on days 1, 2, 3, 4, and 7 post infection with salmonellae (d.p.i.) for cytokines detection, as well as 4 and 7 d.p.i. for other examinations. The time from 1-4 d.p.i. was termed as early phase, and 7 d.p.i. the late phase of infection. The chicks were anaesthetized with intraperitoneal (i.p.) injection of xylazine (Rometar 2%, SPOFA, Czech Republic) and ketamine (Narkoman 5%, SPOFA, Czech Republic) at the doses of 0.6 and 0.7 ml.kg-1 body weight, respectively. After laparotomy, blood was collected into heparinised tubes by intracardial punction and was immediately used for determination of leukocyte numbers. The samples from jejunum and caecum for qRT-PCR assays and caecum for flow cytometry were taken during the necropsy from 5 randomly chosen chicks.
Microbial strains preparation and analysis
Probiotic and bacteriocin-producing strain Enterococcus faecium EF55 (by rifampicin marked to differ it from other enterococci) was cultivated in MRS broth (pH 7.0, Merck, Germany) for 18 h at 37°C, the broth culture was centrifuged at 10,000 × g for 30 min. Cells pellet was resuspended in Ringer solution (pH 7.0) to have cell count 109 colony forming unit (CFU/ml). Bacterial cell count was confirmed by the standard microbiological dilution method. Appropriate dilutions were spread onto M-Enterococcus agar (Difco, USA) enriched with rifampicin as previously described by
Strompfová et al. (2003) and cultivated at 37°C for 24 h. Moreover, PCR confirmation (Woodford et al., 1997) was performed. Salmonella Enteritidis SE147 strain was cultivated in Trypticase soy broth (TSY, Becton, Dickinson and Company [BD], Cockeysville, USA) overnight at 37°C; dilutions were spread onto Brilliant green agar (Becton & Dickinson, USA) and grown colonies were confirmed by PCR (Chiu and Ou, 1996).
For microbial analysis, caeca and faeces were sampled. Fecal samples were taken at day 1 before the experiment (from each group including 55 chicks, 10 mixture samples was examinated) to exclude salmonella infection and to know microbial backround, then at day 7 and at the end of experiment. As mentioned previously, samples were treated by he standard microbiological dilution method (ISO, ratio 1:9), diluted in Ringer solution and the appropriate dilutions were spread onto the media as follows: M-Enterococcus agar (Difco, USA) to count enterococci, M-Enterococcus enriched with rifampicin to count E. faecium EF55, MacConkey agar (BD, USA) for coliforms, CLED agar (Oxoid, Germany) to check enterobacteriae. To check salmonellae including SE147 strain, faecal and caecal samples were pre-cultivated in Rappaport-Vassiliadis medium (Merck, Germany) at 37°C for 48 h; appropriate dilutions (100 µl) were spread onto Brilliant green agar (BD, USA). The colonies grown from the highest dilution of each sample were counted and confirmed by PCR (Chiu and Ou, 1996). The counts of microbiota are expressed in CFU g-1 as mean ± SD.
White blood cell count
Leukocytes were counted in a haemocytometer using Fried-LukaÄová solution (475 μl of solution plus 25 μl of blood). Differential cell counts of 200 cells per slide were done by light microscopy at 1000 magnification using blood smears stained with Hemacolor (Merck, Germany). The total numbers of different subtypes of white blood cells was then calculated: total leukocytes count x proportion of differential cells counted (%)/100.
Caecal intraepithelial lymphocyte isolation and detection
The ceca were placed into cold PBS and processed immediately after sampling. Modified procedure by Swinkels et al. (2007) was used. Whole caeca (3 cm) were opened longitudinally, washed 3 times with cold PBS and cut into pieces of 0.5 cm. Incubation of pieces was done for 45 min at 37°C in PBS containing 1 mM EDTA and 5 mM DTT. The suspension of IEL was centrifuged for 10 min at 250 g and washed twice in cold PBS. The cells were resuspended in 5.104 per 50 μl for flow cytometry imuno-phenotyping. Labelled primary mouse anti-chicken monoclonal antibodies were used in the protocol recommended dilution (Southern Biotechnology Associates, USA). Cells were stained by the direct immunofluorescent method at one of the monoclonal combinations CD3-PE/MHC II-FITC, CD4-PE/CD8αα-FITC, TCRγδ-PE/CD45-FITC, IgM-FITC/IgA-PE. Fifty μl of cell suspension and working solution of specific monoclonal antibodies (MoAbs) were mixed and incubated for 15 min at 37°C. Then, the cells were washed once in PBS (0.5 ml), centrifuged (5 min at 250 g) and resuspended in 0.2 ml PBS with 0.1 % paraformaldehyde. After staining the cells were measured with FACScan flow cytometer (Becton Dickinson, Germany) when a live gate was set for the lymphocyte population in the forward and side scatter plot. The fluorescence data were collected on at least 10,000 lymphocytes and analysed using the Cell Quest programme (BD, Germany). The results were expressed as the relative percentage of lymphocyte subpopulation, which was positive for a specific MoAb.
Homogenization of tissue and isolation of mRNA
The samples of the caecum taken during necropsies were cut into 20 mg pieces, immediately placed into RNALater (Qiagen, UK) and stored at −70°C prior to RNA purification. Single tissue fragment was transferred into 1 ml of TRI reagent (Molecular Research Centre, USA) and homogenized using zirconia silica beads (BioSpec Products, USA) in vortex mixer (Labnet, USA). To separate the phases, 50 μl of 4-bromanisole (Molecular Research Centre, USA) was added and centrifuged at 12000 rpm for 15 min at 4°C. The upper aqueous phase was collected for RNA purification using the RNeasy mini kit (Qiagen, UK) according to the manufacturer's instructions. Turbo DNA-free kit (Ambion, USA) was used for treatment of RNA samples to remove genomic DNA. Purity and concentration of RNA samples was determined specro-photometrically on NanoDrop 200c (Thermo Fischer Scientific, USA) and 1 μg of total RNA was immediately reverse transcribed using iScript cDNA synthesis kit (Bio-Rad, USA). The resulting cDNA was 10× diluted in UltraPure™ DNase/RNase-Free distilled water (Invitrogen, USA) and used as a template in real-time PCR or stored at −20°C until used.
Quantitative RT-PCR
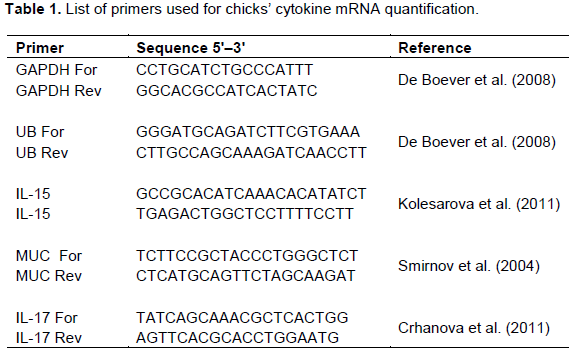
The mRNA levels of mucin, IL-15, and IL-17 were determined separately. The primers used for the quantitative RT-PCR (qRT-PCR) in this study are listed in Table 1. The expression levels of two housekeeping genes, glyceraldehyde-3-phosphate dehydro-genase (GAPDH) and ubiquitin (UB) were used for data normalisation. Amplification and detection of specific products were performed using Maxima SYBR GREEN qPCR mix (Thermo Fisher Scientific, USA) and the CFX 96 RT thermocycler (Bio-Rad, USA) with the temperature-time profile for 45 cycles: initial denaturation 15 min 95°C then denaturation 95°C for 20 s, annealing 60°C for 30 s and elongation 72°C for 30 s. A melting curve from 50 to 95°C with reading at every 0.5°C was performed for each individual RT-PCR plate. The Ct (cycle threshold) values of genes of interest were normalised to an average Ct value of the house-keeping genes (ΔCt) and the relative expression of each representative was calculated as 2–ΔCt.
Statistical analysis
All data were tested by one-way analysis of variance (ANOVA) and Tukey test in Minitab 16 (SC&C Partner, Brno, Czech Republic). The results are given as means ± SD. Differences between mean values for the groups of chickens were consider significant with P < 0.05.
White blood cell counts
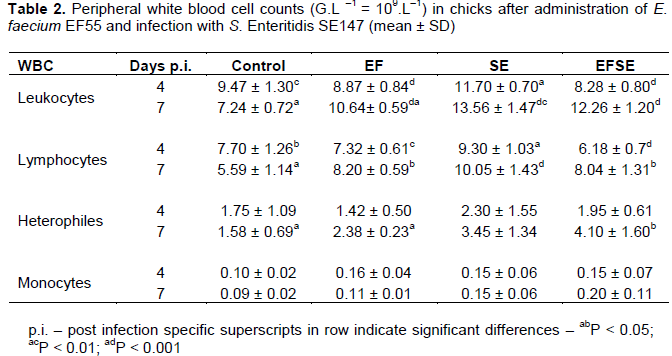
Values of peripheral blood leukocytes increased in the birds of SE group compared to the control group (P < 0.01) with higher decrease in EF and EFSE groups (P < 0.001) on day 4 p.i. (Table 2). On day 7 p.i. an increase of peripheral blood leukocytes in EF, SE and EFSE groups (P < 0.001) was observed compared to controls. Moreover, lower significance (P < 0.01) of those cells was found between EF and SE groups. The numbers of lymphocytes in SE group were higher than those in controls (P < 0.05), EF group (P < 0.01), and EFSE group (P < 0.001) on day 4 p.i. On the other hand, the number of lymphocytes increased in SE group (P < 0.001), as well as in EF and EFSE (P < 0.05) groups compared to the control chicks on day 7 p.i. There was an increase in the number of heterophils in chicks of EFSE group (P < 0.05) compared with the birds in EF group and controls on day 7 p.i.
Phenotyping of intraepithelial lymphocytes in caecum
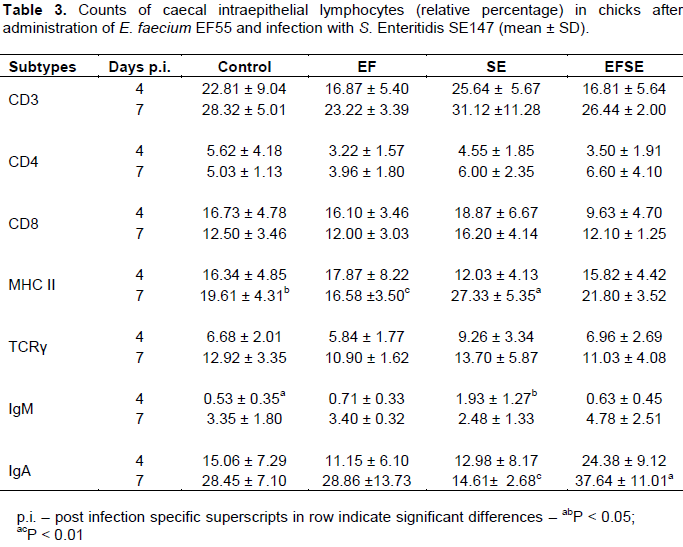
Only MHC II+, IgM+ and IgA+ subpopulations showed statistical significance. The numbers of intraepithelial MHC II+ cells were higher in birds of SE group (P < 0.05) than those in controls and chicks of EF group (P < 0.01) on 7 day p.i (Table 3). Intraepithelial IgM+ lymphocytes increased in SE group (P < 0.05) compared to control chicks on 4 day p.i. There was an increase of IgA+ cells in the group EFSE (P < 0.01) compared to the chicks of the SE group on day 7 p.i.
Intestinal mucin gene mRNA expression
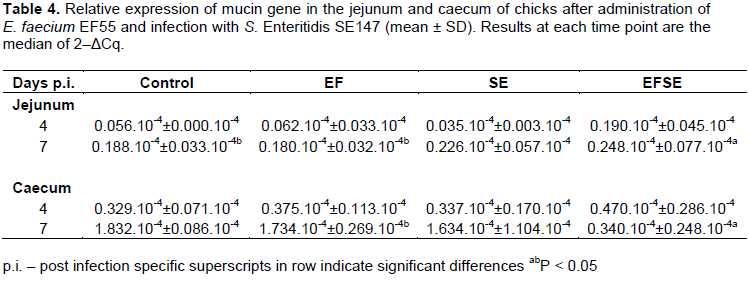
The mRNA expression of mucin gene was increased in the jejunum of EFSE group (P < 0.05) compared to EF group and controls on day 7 p.i. However, mucin gene mRNA expression was the lowest in the caecum in EFSE group in comparison with other groups of chickens, with significance (P < 0.05) to EF group on day 7 p.i. (Table 4).
Cytokines mRNA expression in caecum
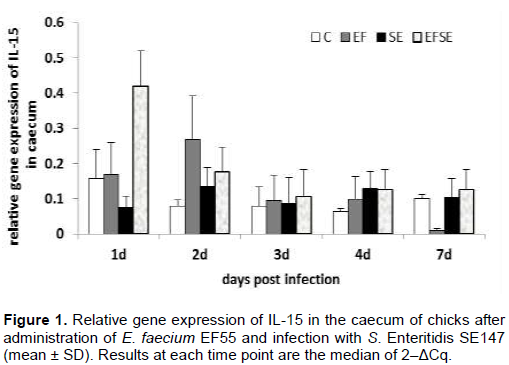
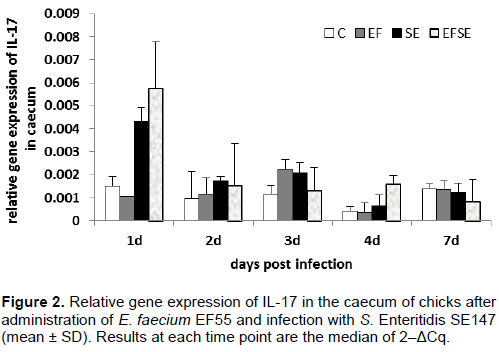
The infection with S.Enteritid is SE147 and administration of E. faecium EF55 did not significantly influence the mRNA expression of IL-15 and IL-17 in the caecum of broilers (Figures 1 and 2). There was only a tendency to increase the mRNA expression of IL-15 and IL-17 on the first day after infection in EFSE group.
Microbial evaluation
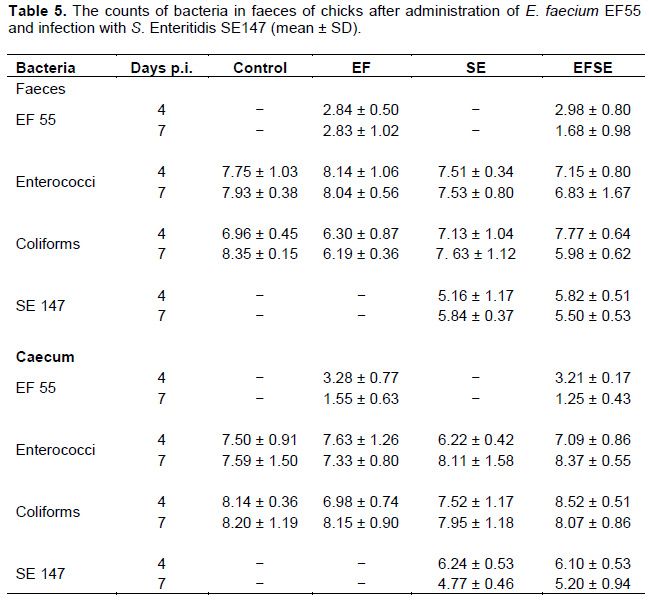
Enterococcus faecium EF55 sufficiently colonized the gastrointestinal tract of chicks. Seven days after probiotic strain administration (4 d.p.i.) faeces and caecum had counts of nearly 103 CFU/g in EF and EFSE groups. However, during the experiment no significant decrease, but slightly reduction of S. Enteritidis SE147 in faeces and caecum in combined EFSE group occurred. No significant differences were found in the counts of enterococci and coliforms bacteria between the experimental and control groups (Table 5).
Earlier works demonstrated that infection of chickens with S. Enteritidis PT4 increased the total number of peripheral blood leukocytes and lymphocytes (Asheg et al., 2003). Similar quantitative changes in the evaluation of blood leukocytes were found in the current study.
Heterophils are critical effector cells in the innate defence of the host against Salmonellae infections. The infiltration of heterophils often accompanies the early phase of infection in the gut (Van Immerseel et al., 2002) and these cells can kill invading pathogens and contribute to recruitment of other immune cell types. Our earlier observations indicate that salmonella infection attracts heterophils to the place of infection, cells which have the ability to kill Salmonella organisms by releasing antimicrobial peptides (ŠevÄíková et al., 2003). The results in the current study revealed that administration of E. faecium EF55 had immunostimulatory effect on the number of heterophils in the peripheral blood of the chicks infected with S. Enteritidis SE147. This suggests the beneficial effect of probiotic against pathogen. Previously, we demonstrated the Enterococcus faecium EF55 capability to increase the percentage of phagocytic activity of heterophils in the chickens infected with Salmonellae (Levkut et al., 2012). Modulation of heterophils by administration of E. faecium EF55 could be one of the important factors in the protection against many intracellular pathogens.
The present results demonstrated milder antimicrobial effect of E. faecium EF55 against S. Enteritidis SE147 presented by tendency to decrease of followed bacteria counts in caecum and faeces at 7 dpi. On the other hand, previous experiments with administration of EF55 and challenge with S. Enteridis PT4 showed significant antimicrobial effect in caecum of chicks 14 dpi (Levkut et al., 2009).
To obtain more information about the numbers and types of lymphocytes that could be recruited for acquired immune responses after infection with S. Enteritidis SE147 and administration of E. faecium EF55, different intraepithelial lymphocyte subpopulations obtained from caecum were examined. In the current experiment increased frequency of MHC II+ cells, and IgM+ lymphocytes was induced with Salmonellae infection, which can be associated with severe host immune response against intracellular bacteria. It is known that MHC II as polymorphic glycoprotein, expressed at the surface of antigen presenting cells, binds exogenously derived antigens and presents them to T lymphocytes. Dendritic cells are able to place class II MHC-antigen complexes on their surface at far higher densities than it can be achieve by macrophages (Kasper et al., 2008). It appears that higher density of MHC II+ cells in caecal epithelium of SE group was associated with alteration of epithelial barrier function and cell damage. On the other hand, administration of Enterococcus faecium EF55 has shown in S. Enteridis SE147 challenged chicks an increase of the IgA+ subpopulation of caecal intraepithelial lymphocytes in later phase of infection in the current experiment. Other investigators demonstrated that IgA is secreted via mucosal epithelial cells and needs to be retained in the immediate mucosal environment to maximize exclusion of pathogens (Linden et al., 2008). Secretory IgA promotes the clearance of antigens and pathogenic microorganisms from the intestinal lumen. Activity of IgA is directed by blocking access of pathogens to epithelial receptors, entrapping them in mucus, and facilitating their removal by peristaltic and mucociliary activities (Mantis et al., 2011).
Higher jejunal mRNA mucin gene expression in later phase of infection, and on the contrary lower caecal mRNA mucin gene expression in current trial, suggests the modulation in the production of mucin into mucosal environment. The higher jejunal mRNA mucin gene expression (non-specific region for S. Enteritidis) can be influenced by the activity of E. faecium EF55. Our earlier work demonstrated that jejunum is very sensitive for mucin secretion after infection (Levkut et al., 2012). Decreased mRNA expression of MUC in the chicks of EFSE group in the late phase of infection can be related to decreased frequency of gramnegative Enterobacteriaceae (Strompfová et al., 2003) and Salmonellae (Levkut et al., 2012) in the caecum and lack of mucin degrading bacterial species (Macfarlane et al., 2005). The result could be lower mRNA expression of observed mucin gene in the late phase of infection. However, it remains an open question about the correlation of the increased number of IgA+ IEL in current trial and the quantity of SIgA antibodies in biofilm formation. Bollinger et al. (2006) have shown that SIgA and mucin facilitate the formation of biofilm by non-pathogenic E. coli on the epithelial cell monolayer grown in vitro.
A wide range of cell types respond strongly to exposure of live Salmonellae by production of proinflammatory cytokines. Administration of E. faecium EF55 revealed a tendency to increase mRNA of IL-15 in the early phase of salmonella infection in both groups with applied probiotic bacteria. Similarly, a tendency to modulate mRNA expression of IL-17 after administration of E. faecium EF55 was observed in the current trial in the early phase of infection. Our results suggest that the increased tendency of mRNA expression IL-15 and IL-17 could participate in attraction of mucosal immunocompetent cells including heterophils and lymphocytes into intestinal mucosa. Interleukin-17 has the ability to upregulate synthesis and secretion of numerous cytokines such as IL-6 which is needed for the differentiation of IgM+ cells into plasma cells (Lillehoj and Okamura, 2003). Finally, the tendency of increased density of caecal IgM+ IEL was found in the groups with administration of E. faecium EF55 in late phase of salmonella infection. Higher frequency of caecal IgA+ IEL could suppress the mRNA expression of IL-15 and IL-17 in that late phase of infection with S. Enteritidis. Previous investigators demonstrated the influence of SIgA on the composition of the intestinal microbiota, downregulation of proinflammatory responses which are normally associated with the uptake of highly pathogenic bacteria (Mantis et al., 2011). Finally, the changes in the composition of intestinal bacteria after application of EF55 were observed in our other experiment (Strompfová et al., 2003).
The results of the current study suggest that there was influence of E. faecium EF55 on mRNA expression of MUC2 gene and production of IgA+ IEL with the anti-inflammatory effect in birds presented by decline salmonellae counts in caecum and faeces of challenged (EFSE) group. This suggestion is supported by the beneficial effect of E. faecium EF55 on the chicks challenged with S. Enteritidis SE147 having higher number of lymphocytes and heterophils in the early or late phase of infection. E. faecium EF55 also increased the number of IgA+ IEL in S. Enteritidis challenged group, which suggests higher immune exclusion. Finally, the administration of E. faecium EF55 had a tendency to increase mRNA expression of IL-15 and IL-17 in the chicks infected with salmonellae in the early phase of infection. The results require further characterization of mucin formation in relation to immunocompetent cells, expressed cell molecules and biofilm formation after application of E. faecium 55 to the chickens infected with pathogen.
The authors have not declared any conflict of interests.
REFERENCES
|
Asheg AA, Levkut M, Revajová V, ŠevÄíková Z, Kolodzieyski L, Pistl J, PilipÄinec E (2003). Spreading of Salmonella enteritidis in the cecum of chickens. Folia Microbiol. 48(2):277-279.
Crossref
|
|
|
|
Bollinger RR, Everett ML, Wahl SD, Lee YH, Orndorff PE, Parker W (2006). Secretory IgA and mucin-mediated biofilm formation by environmental strains of Escherichia coli: role of type 1 pili. Mol. Immunol. 43:378-387.
Crossref
|
|
|
|
|
Chambers JR, Lu X (2002). Probiotic and maternal vaccination for Salmonella control in broiler chickens. J. Appl. Poult. Res. 14:106-116.
|
|
|
|
|
Chiu CH, Ou JT (1996). Rapid identification of Salmonella serovars in faeces by specific detection of virulence genes, invA and spvC, by an enrichment broth culture-multiples CR combination assay. J. Clin. Microbiol. 34(10):2619-2622.
|
|
|
|
|
Crhanova M, Hradecka H, Faldinova M, Matulova M, Havlickova H, Sisak F, Rychlik I (2011). Immune response of chicken gut to natural colonization by gut microflora and to Salmonella enterica serovar Enteritidis infection. Infect. Immun. 79(7):2755-2763.
Crossref
|
|
|
|
|
De Boever S, Vangestel C, De Backer P, Croubels S, Sys SU (2008). Identification and validation of housekeeping genes as internal control for gene expression in an intravenous LPS inflammation model in chickens. Vet. Immunol. Immunopathol. 122(3-4):312-317.
Crossref
|
|
|
|
|
Fossiez F, Djossou O, Chomarat P, Flores-Romo L, Ait-Yahia S, Maat C, Garrone P, Garcia E, Saeland S, Blanchard D, Gaillard C, Das Mahapatra B, Rouvier E, Golstein P, Banchereau J, Lebecque S (1996). T cell interleukin-17 induces stromal cells to produce proinflammatory and hematopoietic cytokines. J. Exp. Med. 183(6):2593-2603.
Crossref
|
|
|
|
|
Hong YH, Lillehoj HS, Lillehoj EP, Lee SH (2006). Changes in immune-related gene expression and intestinal lymphocyte subpopulations following Eimeria maxima infection of chickens. Vet. Immunol. Immunopathol. 114(3-4):259-272.
Crossref
|
|
|
|
|
Husband AJ (2002). Mucosal memory-maintenance and recruitment. Vet. Immunol. Immunopathol. 87(3-4):131-136.
Crossref
|
|
|
|
|
Kaiser P, Rothwell L, Vasicek D, Hala K (2002). A role for IL-15 in driving the onset of spontaneous autoimmune thyriditis. J. Immunol. 168(8):4216-4220.
Crossref
|
|
|
|
|
Kasper B, Kothlow S, Butter C (2008). Avian antigen presenting cells. Davison F, Kasper B, Schat KA (Ed.) Avian Immunology. Academic Press, Elsevier. pp. 183-202.
Crossref
|
|
|
|
|
Kolesarova M, Spisakova V, Matulova M, Crhanova M, Sisak F, Rychlik I (2011). Characterization of basal expression of selected cytokines in the liver, spleen, and respiratory, reproductive and intestinal tract of hens. Vet. Med. Czech 56(7):325-332.
|
|
|
|
|
Levkut M, Pistl J, Lauková A, Revajová V, Herich R, ŠevÄíková Z, Strompfová V, Szabóová R, KokinÄáková T (2009). Antimicrobial activity of Enterococcus faecium EF55 against Salmonella Enteritidis in chickens. Acta Vet.-Hung. 57(1):13-24.
Crossref
|
|
|
|
|
Levkut M, Revajová V, Lauková A, ŠevÄíková Z, Spišáková V, Faixová Z, Levkutová M, Strompfová V, Pistl J, Levkut M (2012). Leukocytic responses and intestinal mucin dynamics of broilers protected with Enterococcus faecium EF55 and challenged with Salmonella Enteritidis. Res. Vet. Sci. 93(1):195-201.
Crossref
|
|
|
|
|
Lillehoj HS, Min W, Choi KD, Babu US, Burnside J, Miyamoto T, Rosenthal BM, Lillehoj EP (2001). Molecular, cellular, and functional characterization of chicken cytokines homologous to mammalian IL-15 and IL-2. Vet. Immunol. Immunopathol. 82(3-4):229-244.
Crossref
|
|
|
|
|
Lillehoj HS, Okamura M (2003). Host immunity and vaccine development to coccidian and Salmonella infections in chickens. Poult. Sci. 40(3):151-193.
Crossref
|
|
|
|
|
Linden SK, Sutton P, Karlsson NG, Korolok V, McGuckin MA (2008). Mucins in the mucosal barrier to infection. Mucosal Immunol. 1(3):183-197.
Crossref
|
|
|
|
|
Macfarlane S, Woodmansey EJ, Macfarlane GT (2005). Colonization of mucin by human intestinal bacteria and establishment of biofilm communities in a two-stage continuous culture system. Appl. Environ. Microbiol. 71(11):7483-7492.
Crossref
|
|
|
|
|
Mantis NJ, Rol N, Corthesy B (2011). Secretory IgA's complex roles in immunity and mucosal homeostasis in the gut. Mucosal Immunol. 4(6):603-611.
Crossref
|
|
|
|
|
ŠevÄíková Z, Asheg AA, Kolodzieyski L, Ciganková V, Komorová T, Levkut M (2003). Heterophils and macrophage-like cells in the caeca of chicks after experimental infection with Salmonella enteritidis PT4. Acta Vet.-Brno 72:565-570.
Crossref
|
|
|
|
|
Strompfová V, Mudroňová D, Lauková A (2003). Effect of bacteriocin-like substance produced by Enterococcus faecium EF55 on the composition of avian gastrointestinal microflora. Acta Vet.-Brno 72:559-564.
Crossref
|
|
|
|
|
Swinkels WJ, Post J, Cornellisen JB, Engel B, Boersma WJ, Rebel JM (2007). Immune response to an Eimeria acervulina infection in different broilers lines. Vet. Immunol. Immunopathol. 117(1-2):26-34.
Crossref
|
|
|
|
|
Van Immerseel F, De Buck J, De Smet I, Mast J, Haesebrouck F, Ducatelle R (2002). Dynamics of immune cell infiltration in the caecal lamina propria of chickens after neonatal infection with a Salmonella enteritidis strain. Dev. Comp. Immunol. 26(4):355-364.
Crossref
|
|
|
|
|
Smirnov A, Sklan D, Uni Z (2004). Mucin dynamics in the chick small intestine are altered by starvation. J. Nutr. 134(4):736-742.
|
|
|
|
|
Woodford N, Egelton MC, Morrison D (1997). Comparison of PCR with phenotypic methods for the speciation of enterococci. Adv. Exp. Med. Biol. 418:405-408.
Crossref
|
|