Full Length Research Paper

ABSTRACT
This study was designed to monitor the development of the hormonal profiles of the Nubian bucks at puberty. Thirty two (32) Nubain male kids between 1 day and 24 weeks having body weights of 2.3 to 13.5 kg were utilized in this study. All kids were injected with human chorionic gonadotropin (hCG). Blood samples were collected from the jugular veins of the animals immediately before hCG injection and 1 h after the injection. Plasma was separated from the samples and stored at 20°C until it was assayed for determination of hormonal levels. After hCG injection, the plasma level of testosterone was significantly higher (p< 0.01) than the level of the hormone before hCG treatment. Significant increase was also observed in luteinizing hormone (LH) values after hCG injection (P≤ 0.01). Plasma follicles stimulating hormone (FSH) had one peak in week 8 in samples taken before hCG injection. This was followed after two weeks by the first elevation of LH. Plasma prolactin (PRL) concentration showed a gradual increase with age, reaching the peak in week 18. Based on the first appearance of spermatozoa in the lumina of the seminiferous tubules, penile separation and attainment of maximal levels of testosterone, LH, FSH and PRL, pubertal age was reached between 18 to 20 weeks.
Key words: Nubian bucks, hormonal profiles, puberty.
INTRODUCTION
Goat population in Sudan is around 39.952.000 heads. Nubian goats constitute 60% of this population (MARAF, 2001). They are found in north and central parts of the country around the River Nile banks. They are well reputed as good dairy goats in the tropics. They are the progenitor of the famous Anglo-Nubian goats. They are middle-sized, have black hairy coat and long droopy white ears spotted with black. Both males and females have medium horns and sometimes wattles. Nubian goats are polyoestous with twining rate of 2 to 4 kids. Females produce around 73 kg of milk during lactation period of 143 days with a daily yield of 1.5 to 2.0 kg (Williamson and Payne, 1987).
Hormonal profiles of goat have been investigated by several authors (Goyal et al., 1994; Walkden-Brown et al., 1994; Vera-Avila et al., 1997; Delgadillo et al., 2002; Al-Daker and Salha, 2004; Barkawi et al., 2006; Al-Sobayil et al., 2008; Omari, 2012).
Hormones as endocrine secretion are transported by the vascular system to aid in the integration of body processes by their stimulatory or inhibitory effects on target organs (Haenlein, 1992).
Testosterone is the most important of androgens and is secreted by the testicular interstitial cells of Leydig. Testosterone and its by-products, androsterone and dehydrosterone are bound to plasma proteins and are rapidly used by target organs and degraded by the liver and kidneys. Testosterone and related hormones are responsible for male secondary sex characteristics, body conformation, muscular development and libido. They are also responsible for the growth and development of secondary sex glands of the males, as well as maintenance of the viability of the spermatozoa; they stimulate penile growth and separation from the prepuce.
The luteinizing (LH) and follicle stimulating hormones are glycoproteins secreted by the β-cells (basophils) gonadotrophs of the anterior pituitary and consist of two subunits. The α-subunit is identical in the hormones whereas the β subunit is biochemically unique and determines the bioactivity. The circulating concentration of LH and FSH is controlled by a negative feedback effect on the hypothalamus due to steroidal hormones, so they are secreted in pulses rather than constant amounts. The male LH stimulates interstitial cells to produce androgen. FSH in mature males is associated with the stimulation and maintenance of spermatogenesis.
Prolactin (PRL) is a single-stranded polypeptide hormone secreted by the mammotrophs of the pituitary gland and is composed of 198 amino acids. Regarding structure and biology, PRL, growth hormone (GH) and placental lactogen (HPL) are of the same type. PRL plays an essential role in the secretion of milk and has the ability to suppress gonadal function.
Studies on puberty have been reported for bull (Abdel Raouf, 1960; Renaville et al., 1993), sheep and goat (Louw and Joubert, 1964; Lord et al., 1991), ram (Dun, 1955), camel (Abdel Rahim, 1997), rhesus macaques (Bercovitch, 1993) and man (Martha and Reiter, 1991). Puberty, the culmination of a multitude of developmental processes at the hypothalamic, pituitary and gonadal levels, is essentially manifested by the episodic release of testosterone ((Renaville et al., 1983; Schams et al., 1988) which serves as a primary regulator for other major physiological changes during this period (Martha and Reiter, 1991). Associated with this is an increase in plasma concentrations of testosterone, and growth hormone (GH) discharge may precede the onset of puberty (Thompson et al., 1972). Puberty is reached when sexual organs have become fully developed, sexual instincts are prominent and reproduction is possible (Abdel-Raouf, 1960). Puberty age is considered as an economic trait. If the animals reach puberty and sexual maturity at early age, more benefit would be obtained from their productive life. Thus, remarkable physiological changes and/orphenotypic features associated with occurrence of puberty constitute helpful tools for the selection of breeding males.
Literature on age at puberty and the associated internal and external changes in goats (Babiker and Makawi, 2005; Elhammali et al., 2013; Shaaeldin et al., 2018) is scarce generally.
The objective of this study is to monitor the changes in hormonal profiles of Nubian bucks and the inter-relationship between them during the age of puberty.
MATERIALS AND METHODS
This study was conducted at Animal Production Research Centre in Hilat Kuku, Khartoum North, Sudan. Sudan is located at latitude 15° 36’ N, longitude 32° 33’ E and altitude of 380 m above sea level. Thirty-two Nubian bucks were assigned for this study. Their ages ranged between birth and 6 months.
Up to the age of weaning, the kids were dependent on their mothers. After weaning the animals were fed on green alfalfa and a concentrate ration composed of wheat bran (43%), molasses (32%), ground nut cake (20%), urea (3%) and salt (2%). Water and minerals licks were offered to them ad-libitum.
Blood samples were obtained from 32 Nubian bucks from birth up to six months of age. Samples were taken at 15 days interval for investigation. The different groups were represented by at least two animals each.
For hormonal assay, an initial sample of 5 ml of blood was collected from the jugular vein of the animals; then the animals were injected (1/m) with hCG (300 IU/animal). A second sample was withdrawn one hour later to monitor the post-hCG injection level of hormones. The samples were then centrifuged at 3000 rpm for 15 min. The supernatant was decanted into fresh test tubes and stored at -20°C to be used for hormone assay.
Measurement of hormones
Immunoradiometric assay (IRMA) and radioimmunoassay (RIA) techniques were used for the estimation of hormones. In the IRMA assay, the antibody was the labeled tracer, and in the RIA assay, the antigen was the labeled tracer.
Measurement of hormones by IRMA technique
Measurement of plasma LH and FSH: For the assay of LH and FSH, the following reagents were introduced into the reaction tube:
50 UL sample or standard, 300 UL assay buffer, 50 UL labeled antibody (anti-LH or anti-FSH) and total tube (approx. 60,000 rpm). 50UL solid phase first antibody tubes were vortexed and placed on rotary mixer overnight; 2 ml of wash buffer was added and centrifuged for 5 min at 1000 rpm; supernatant was aspirated. The washing step was repeated, aspirated again and counted for 1 min.
Measurement of plasma prolactin: The solid phase was reacted with the sample or standard at room temperature for approximately 3 h following a washing step. The radio-iodinated antibody was added and the reaction continued overnight.
This assay method was the same as that used for plasma FSH and LH except 350 UL of assay buffer was used instead of 300 UL and the centrifugation took a long time, up to 10 min.\
Measurement of testosterone by RIA technique
All reagents were placed at room temperature, mixed gently and used in the assay. The reagents were added as follows:
Standard or sample (50 UL)
125 (1) Testosterone (tracer) (500 UL)
Testosterone Antiserum (500 UL)
All tubes were vortexed gently avoiding foaming and incubated for 2 h at 37°C. Assay buffer of 500 UL was added. The tubes were again vortexed and incubated for 15 min at room temperature; they were centrifuged for 15 min at 100 g. Supernatants were decanted; the tubes were blotted gently to get rid of the remaining drops of liquid. Radioactivity was counted for one minute.
The data were subjected to analysis of variance using Statistical Package for Social Science. Mean comparisons were made using Fisher’s least significant test, while the relationship between various parameters was expressed using the Pearson correlation coefficient (Kinner and Gray, 1999).
RESULTS
The results included the plasma concentrations of some hormones (testosterone, LH, FSH, and prolactin) before and after hCG injection. The results are summarized in Table 1.
Concentrations of plasma hormones
The blood hormones levels before and after hCG injection are illustrated in Table 1.
Testosterone
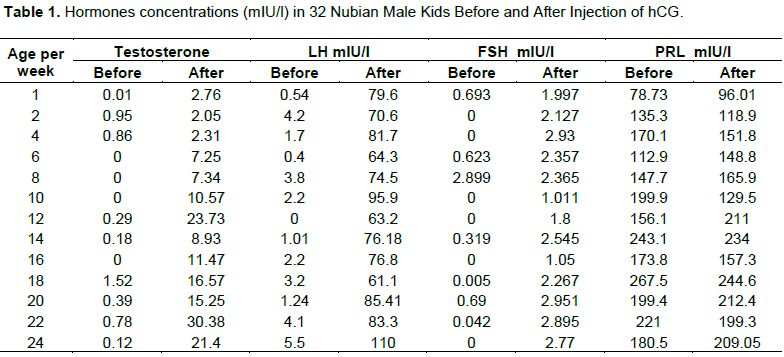
The plasma level of testosterone after hCG injection was significantly higher (p< 0.01) than the level of the hormone before hCG treatment. There was a gradual increase in the testosterone concentration with advancing age throughout the study period (Figure 1). The most important feature in testosterone pattern of secretion was the occurrence of two spikes at the age of 12 and 22 weeks.
Luteinizing hormone (LH)
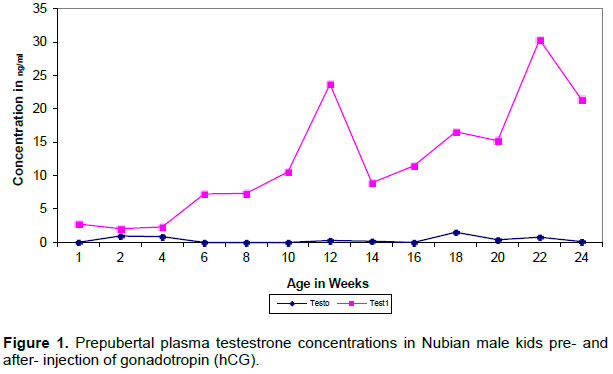
Plasma luteinizing hormone concentrations before hCG injection showed basal levels throughout the prepubertal period (Figure 2).
Significant increase was observed in LH values after hCG injection (p≤0.01). The pattern of LH secretion after hCG injection maintained levels around 70 ng/ml, with age, except for two sudden significant rises around 10 and 20 weeks of age.
Follicle stimulating hormone (FSH)
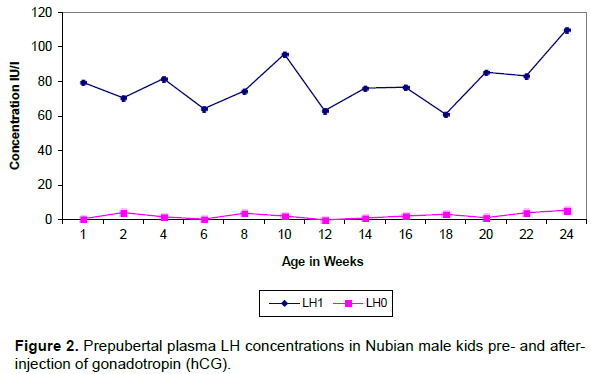
The result showed basal levels of plasma FSH before hCG injection except for a sudden rise (peak) around the age of 8 weeks (Figure 3). After hCG injection FSH values were significantly (p≤0.01) higher compared with the values before hCG injection. Plasma FSH concentration followed an irregular pattern with its values fluctuating between 1 a
Prolactin (PRL)
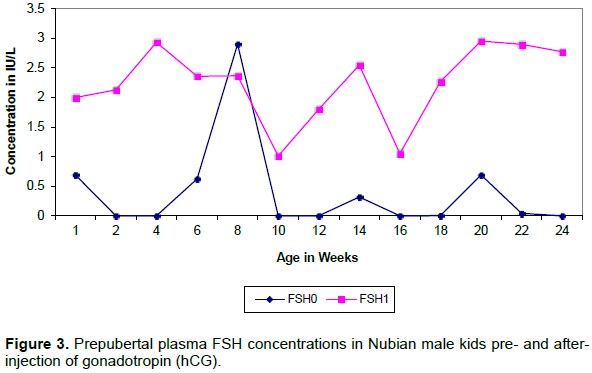
Plasma PRL concentration showed a gradual increase with age reaching the peak at 18 weeks (Figure 4).
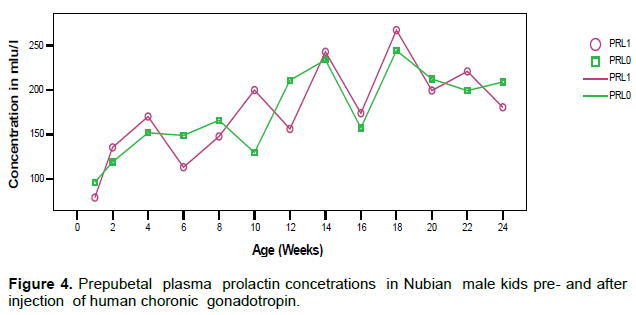
The levels of prolactin before and after hCG injection showed no significant differences (p> 0.05). PRL secretion showed a pulsatile pattern with peaks occurring at almost regular intervals (4-6 weeks) and the frequency and magnitude of peaks decreased after the age of 20 week.
Correlation between the hormones
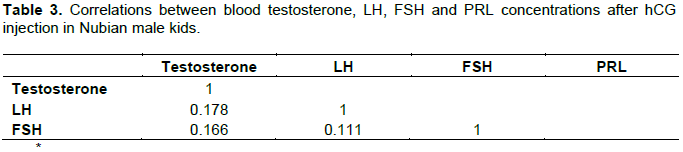
Plasma LH and testosterone concentration after hCG treatment showed biphasic pattern of elevations (Figures 5 and 6). Consistently LH peaks preceded the testosterone elevations by two weeks (Table 3).
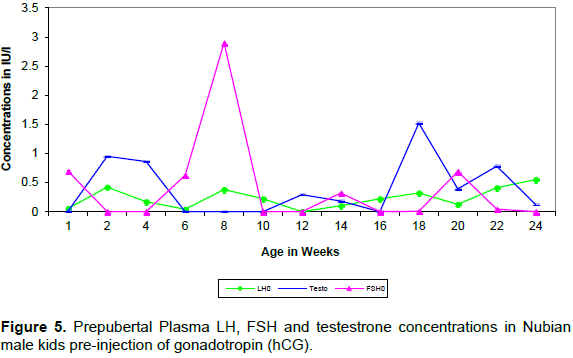
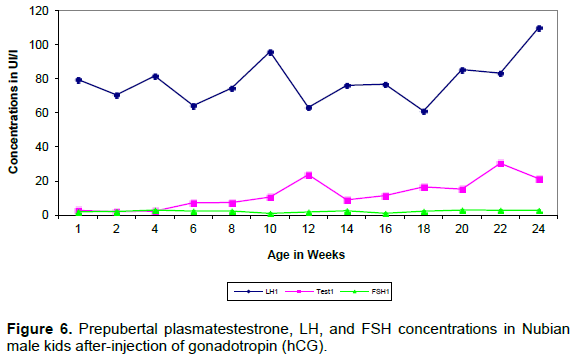
Plasma FSH showed one peak at 8 weeks of age in the sample taken before hCG injection. This was followed in 2 weeks by the first elevation of LH.
Association of hormonal changes and pubertal development of the genital tract
The first significant elevation in FSH, LH and testosterone occurred between week 8 and week 12 (10-14 weeks before puberty). The increase in the levels of these hormones in plasma coincided with an abrupt increase in the diameters of the seminiferous tubules, the epididymis, ductus deferens, and penis. Moreover the first appearance of secondary spermatocytes and initiation of penile separation from the prepuce also occurred during this period.
However, the second surge of testosterone, LH and FSH occurred between weeks 20 and 24. This was associated with the first appearance of spermatids, spermatozoa and completion of the penile separation. Based on the first appearance of spermatozoa in the tubules, puberty was reached in Nubain male kids at the age of 22 weeks (Table 2).
DISCUSSION
The results obtained in this study indicated that testosterone and LH levels before hCG treatment showed a very low concentration compared to that of FSH which showed high levels and reached the peak around 8 weeks of age. The pulsatile (episodic) nature of testosterone and LH secretion necessitates the injection of hCG to monitor their maximum levels with advancing age towards puberty (Renaville et al., 1993; Schams, 1988). Prolactin levels showed progressive increase with age reaching maximum level at 8 weeks of age. This confirms that hormonal levels increase from birth to puberty, and the levels reach higher values around puberty to reach the peak a few weeks after puberty (Renaville et al., 1993; Rawlings, 1972). The results after hCG treatment revealed that the level of testosterone, LH and FSH showed significant increase with age, till they reached the peak around 20 to 24 weeks. This is in accordance with the findings reported in ram (Dufour et al., 1984), Siberain hamster (Meredith et al., 1998) and sheep (Reviers et al., 1980). This result as well revealed the significant effect of hCG injection on testosterone and FSH levels. After one hour of hCG administration, the results showed high levels of these hormones which have significant correlation with age. This is similar to the finding of Meredith et al. (1998). Accordingly, there is a significant effect of age and this is similar to the finding of Walkden-Brown et al. (1994) in Australian Cashmere Goats.
Although this study was conducted in different seasons, the results like those of Omari (2012) were not affected by the seasonal changes. This is probably because the Nubian goat is a tropical breed and has no seasonal breeding pattern (Walkden-Brown et al., 1994; Vera-Avila et al., 1997; Kurohmaru and Nishida, 1987; Mudewli et al., 1997).
The comparison between the levels of hormones before and after hCG injection showed that the highest levels of FSH were followed by LH peaks; the lowest values were those of testosterone. This confirms the findings of Renaville et al. (1993), Rawlings (1972) and Courot (1978) in which it was shown that hormonal levels increased from birth onwards and reached a peak around the age of puberty. This indicates that commencement of reproductive function in bucks has a strong correlation with increase in testosterone concentration in accordance with age (Bercovitch, 1993). The result is suggestive of a highly significant correlation (P≤0.01) between FSH and testosterone levels before and after hCG treatment.
The prolactin has not been demonstrated to have specific effect in male reproduction or response to hCG injection, but in general, it was known to lower blood pressure (Haenlein, 1992).
A good correlation was shown between age, on one hand, and scrotal circumference, heart-girth and body weight, on the other hand. The body measurements reached their maximum at the age of puberty (22 - 24 weeks) (Shaaeldin et al., 2018). Similar findings were also reported in man (Forest et al., 1976), monkeys (Mann et al., 1994; Lunn et al., 1997), ram (Dun, 1955) and sheep (Louw and Joubert, 1964).
The environment for the bucks used in this study was the same with regard to feeding, management and housing. Nutrition however was not considered as a variable. This is in contrast with other findings which focused on nutrition as being the most important factor that can affect body measurements in small domestic animals (Setchel et al., 1965; Martin et al., 1994; Thwaites, 1995a).
For the determination of age of puberty, signs of commencement of spermatogenesis were considered in this study. Thus, puberty in Nubian bucks was reached at 22 weeks of age. Moreover, penile separation was also completed at the same age. Similar findings have been reported on measurement of scrotal circumference, heart girth, and body weight in other breeds of goats and sheep. They indicate that puberty is reached at 18 to 22 weeks (Shaaeldin et al., 2018). However, other breeds of goat and sheep reach puberty between 16 and 21 weeks (Louw and Joubert, 1964). Dyrmundsson (1973) noticed that there were great differences in age of puberty and body weight between ram lambs of various breeds of sheep. Large animals are said to reach puberty between 23 to 26 weeks (Abdel Rahim, 1997).
In the present study, signs of puberty in the Nubian bucks appeared around 22-24 weeks. They reach maturity during the early weeks (Elhammali et al, 2013) and reach one year few weeks later (Renaville et al., 1993).
The two pituitary gonadotropins, LH and FSH, are necessary to maintain the gonadal functions in male animals through the stimulation of spermatogenesis. Full spermatogenesis cannot be accomplished without the effect of LH, known as the interstitial cell stimulating hormone (ICSH) that facilitates the production of testosterone.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
This work has been supported by generous offers from Ahfad University for Women and the Gordan Momeral Fund.
REFERENCES
|
Abdel Rahim SEA (1997). Studies on the age of puberty of male camels (Camelusdromedarius) in Saudi Arabia. The Veterinary Journal 154:79-83. |
|
|
Abdel-Raouf M (1960). The postnatal development of the reproductive organs in bulls with special reference to puberty.ActaEndocrinologica 34 (49):1-109. |
|
|
Al-Daker MB, Salhab SA (2004). Determination of testosterone and LH levels in growing Shami male goats. Journal of Damascus University of Agriculture Science 20:29-44. |
|
|
Al-Sobayil KA, Zeitoun MM, Khalil MH, Abdel-Salam AM (2008).Effect of oral administration of a functionalsynbiotic syrup on libido, semen characteristics, serum testosterone and liver and kidney function of goat's bucks. Asian Journal of Biological Science 1:11-18. |
|
|
Babiker EA, Makawi SA (2005). The Influnce of Season on Sexual Behavior and Semen quality of Nubian Bucks in Sudan. Sudan Journal of Veterinary Science and Animal Husbandry 44(1&2):83-102. |
|
|
Barkawi AH, Elsayed EH, Ashour G, Shehata E (2006).Seasonal changes in semen characteristics, hormonal profiles and testicular activity in Zaraibi goats. Small Ruminant Research 66:209-213. |
|
|
Bercovitch FB (1993). Dominance rank and reproductive maturation in male rhesus macaques (Macacamulatta). Journal of Reproduction and Fertility 99:113-120. |
|
|
Courot M (1978). Prepubertal development and puberty: comparative aspects. International Journal of Andrology 1:11-20. |
|
|
Delgadillo JA, Flores Veliz FG, Hernandez HF, Duarte G, Vielma J, Poindron P, Chemineau P, Malpux B (2002). Induction of sexual activity in lactating anovulatory female goats using male goats treated only with artificially long days. Journal of Animal Science 80:2780-2786. |
|
|
Dufour JJ, Fahmy MH, Minvielle F (1984). Seasonal changes in breeding activity, testicular size, testosterone concentration and seminal characteristics in rams with longer or short breeding season. Journal of Animal Science 58:416-422. |
|
|
Dun RB (1955). Puberty in merino rams. Australian Veterinary Journal 31:104-106. |
|
|
Dyrmundsson OR (1973).Puberty and early reproductive performance in Sheep. II. Ram Lambs. Animal Breeding Abstracts 41:419-430. |
|
|
Elhammali NS, Alqurashi AM, Ibrahim MT, Elsheikh AS (2013). Puberty of crossbred male goat kids Journal of American Science 9(4):95-99. |
|
|
Forest MG, Pereti D, Bertrand J (1976). Hypothalamic - pituitary - gondal relationships in man from birth to puberty.Clinical Endocrinology 5:551-569. |
|
|
Goyal HO, Hutto V, Maloney MA (1994). Effects of androgen deprivation in the goat epididymis. Acta Anatomica 150:127-135. |
|
|
Haenlein GFW (1992). Hormones.Goat handbook.Docn, ooooooo51. No, F-8 . |
|
|
Kinner PR, Gray CD (1999). SPSS for windows (3rd. edn.). Psychology Press Ltd., University of Abrerdeen, UK. |
|
|
Kurohmaru M, Nishida T (1987). Three-dimensional structure of the Sertoli cell in the Shiba goat. Archives of Histolology Japan 50:515-523. |
|
|
Lord APD, Martin AA, Walton PE, Ballard FJ, Read LC (1991). Insulin-like growth factor-binding proteins in tissue fluids from the lamb. Journal of Endocrinology 129:59-68. |
|
|
Louw DFJ, Joubert DM (1964). Puberty in the male Dorper sheep and Boer goat. Journal of Agricultural Science 7:509-520. |
|
|
Lunn SF, Cowen GM, Fraser HM (1997). Blockade of the neonatal increase in testosterone by a GnRH antagonist: the free androgen index, reproductive capacity and postmortem finding in the male marmoset monkey. Journal of Endocrinology 154:125-131. |
|
|
Mann DR, Ansari AA, Akinbami MA, Wallen K, Could KG, Mcclue HM (1994). Neonatal treatment with luteinizing hormone, gonadal and releasing hormone after peripheral lymphocyte subsets and cellular and humorally mediated immuno responses in juvenile and adult male monkeys. Journal of Clinical Endocrinology and Metabolism 78:292-298. |
|
|
Martha PM, Reiter EO (1991). Pubertal growth and growth hormone secretion. Endocrinology and Metabolism Clinics of North America 20:165-182. |
|
|
Martin GB, Tjondronegore S, Blackberry MA (1994). Effects of nutrition on testicular size and concentrations of gonadotrophins, testosterone and inhibin in plasma of mature male sheep. Journal of Reproduction and Fertility 101:121. |
|
|
Meredith JM, Turek FW, Levine JE (1998). Effect of gonadotropin-releasing hormone plusefrequerncy modulation on the reproductive axis of photoinhibited male Siberian hamsters. Biology of Reproduction 59:813-819. |
|
|
Mudewli DS, Sanford LM, Palmer WM, Howland BE (1997). Secretory patterns and circadian and seasonal changes in lutinizing hormone, follicle stimulating hormone, prolactin and testosterone in the male Pygmy goat. Journal of Animal Science 49:543. |
|
|
Omari HAl (2012). Study of Testosterone Concentrations During Breeding Season of two Breeds of Goat Bucks and their Under Exogenous GnRH Treatments. Asian Journal of Animal and Veterinary Advances 7:693-701. |
|
|
Rawlings NC, Hafs, HD, Swason LV (1972).Testicular and plasma androgens in Holsestein bulls from birth through puberty.Journal of Animal Science 34:435-440. |
|
|
Renaville R, Devolder A, Massart S, Sneyers M, Burny A, Portetelle D (1993). Changes in the hypophysial-gonadal axis during the onset of puberty in young bulls. Journal of Reproduction and Fertility 99:443- 449. |
|
|
Reviers PM, Reviers MTH, Blanc MR, Brillard JP, Courot M, Pelletier J (1980).Control of Sertoli and germ cell populations in the cock and sheep testes. Reproduction Nutrition Development 20:241-249. |
|
|
Schams D, Winkler V, Schallenberger E, Karg H (1988). Wachstumshomonud "insulin-like growth factor-1 (somatomedin c)"-Blutspiegelbei Rinderm von der geburtbisnach des Pubertat Deutsches Tierarztliches Wochenschift 95:360-362. |
|
|
Setchel BP, Waites GM, Linder HR (1965). Effect of undernutrition on testosterone in the ram. Journal of Reproduction and Fertility 9:149. |
|
|
Shaaeldin SA, Makawi SA, Tingari MD, Eltayeb AEA (2018). Associatisson of body weight, scrotal circumference, heart girth and penile development with spermatogenesis in the Nubian bucks. Journal of Veterinary Medicine and Animal Health 10(9):217-223. |
|
|
Thompson RG, Rodriguez A, Kowarski A, Migeon CJ, Blizzard RM (1972). Integrated concentrations of GH correlated with plasma testosterone and bone age in pre-and adolescent males. Journal of Clinical Endocrinology and Metabolism 35:334-337. |
|
|
Thwaites CJ (1995a). Effect of undernutrition on the size and tone of the rams testes. Small Ruminant Research 16:283. |
|
|
Vera-Avila HR, Forbes TDA, Berardinelli JG, Randel RD (1997). Effect of dietary phenolic amines on testicular function and luleinizing hormone secretion in male angora goats.Journal of Animal Science 75:1612-1620. |
|
|
Walkden-Brown SW, Restall BJ, Norton BW, Scaramuzzi RJ, Martin GB (1994). Effect of nutrition on seasonal patterns of LH, FSH and testosterone concentration, testicular mass, sebaceous gland volume and odour in Australian Cashmere goats. Journal of Reproductive (on) and Fertility 102:351-360. |
|
|
Williamson G, Payne WJA (1987). An introduction to animal husbandry in the tropics (3rd edit). Longman Group, Hong Kong. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


