Review

ABSTRACT
Tick-borne diseases (TBDs) are a major economic constraint to livestock production affecting the productivity of livestock worldwide. Identification of these haemoprotozan and rickettsial infections is essential in understanding the epidemiology and it is important to distinguish between species and subspecies involved. Conventional techniques including serological and microscopic examinations do not always meet these requirements. Clinical diagnostic and surveillance tools, such as the complement fixation test (CFT), the indirect fluorescent antibody test (IFAT) and the enzyme linked immunosorbent assay (ELISA) have been successfully used over decades. In addition, DNA-based tests for diagnosis, differentiation and characterisation of different haemoparasites have been developed. Molecular diagnostic techniques, such as DNA hybridization and polymerase chain reaction (PCR), allow detection of parasites in blood, tissue or ticks with high levels of sensitivity, specificity and reliability. In addition, some techniques can identify multiple pathogens in the same samples. Furthermore, these techniques can also be exploited to identify unambiguous species and subspecies. Under the precondition that these tests are correctly designed and validated, they provide a powerful tool for epidemiology, with greater advantages of affordability and amenability to standardization. The implementation of these techniques for studying TBDs worldwide will be invaluable. Thus, the aim of this study is to put together the details of the techniques in the form of small review consultation of the practitioners and researchers.
Key words: Diagnosis, livestock, molecular, haemoparasites, tick-borne diseases.
INTRODUCTION
‘Diagnosis’ is an art of knowing about the cause of a particular disease (Dia = through, gnosis = knowledge). Diagnosis of parasitic infections has changed remarkably as technical skills and knowledge have expanded. Ticks rank first as arthropod vectors of protozoa, rickettsiae, bacteria and viruses in nonhuman vertebrates and rank second only to mosquitoes as vectors of pathogens to humans (Zhou et al., 2009). Though tick borne diseases (TBDs) differ among ecological regions, their impact on animal production is more important in tropical and subtropical regions of the world as they pose major threat to the health and management of livestock in these regions (Jongejan and Uilenberg, 1994). These diseases are ranked very high in terms of their impact on the livelihood of resource poor farming communities in developing countries (Perry et al., 2002; Minjaw and McLeod, 2003; Bishop et al., 2008). The most important tick-borne haemoparasitic disease in veterinary medicine include the protozoan parasite of Theileria species (e.g. Theileria annulata, Theileria parva, Theileria lestoquardi), Babesia species (e.g. Babesia bovis, Babesia bigemina, Babesia ovis) and Rickettsial species (e.g. Cowdria ruminantium, Anaplasma marginale, Anaplasma centrale) (Uilenberg, 2006). In addition, several novel Babesia and Theileria (Theileria uilenbergi and Theileria luwenshuni) species were isolated from naturally infected sheep in China, where it cause severe and often lethal disease (Yin et al., 2007; Guan et al., 2010).
The direct impacts of these tick-borne pathogens are reflected in reduction of production, loss of weight and death of substantial proportion of the affected animals. By reducing the losses due to tick-borne diseases there would be a better chance to increase livestock production. The effective management of tick-borne haemoparasitic diseases requires rapid, reliable and highly sensitive diagnostic tests, which can also serve to monitor the effectiveness of the therapeutic and prophy-lactic measures. The common conventional method (history, blood or tissue smears examination, clinical signs, postmortem lesions and conventional serology) provide the basic needs for diagnosis, but have the disadvantage of sensitivity, specificity, involvement of expertise, labour intensiveness, etc., so do not meet complete requirements. The serodiagnosis is not different between current and past infection as the animal may already have cleared the pathogen, but remain seropositive. Nucleic acid based diagnostics, particularly, a wide range of DNA based techniques have been developed and validated for identification, characteri-zation and pathogenic studies for various pathogens. The polymerase chain reaction (PCR) based assays permit identification of parasite at levels far below the detection limit of the commonly used parasitological techniques. Several techniques have been developed separately for detection for each species tick-borne haemoprotozoa (Figueroa and Buening, 1995) (Table 1). In this review, a detailed discussion on diagnostic procedures from classical to molecular approaches including clinical, parasitological, serological and molecular techniques is made.
CONVENTIONAL DIAGNOSTIC TECHNIQUES
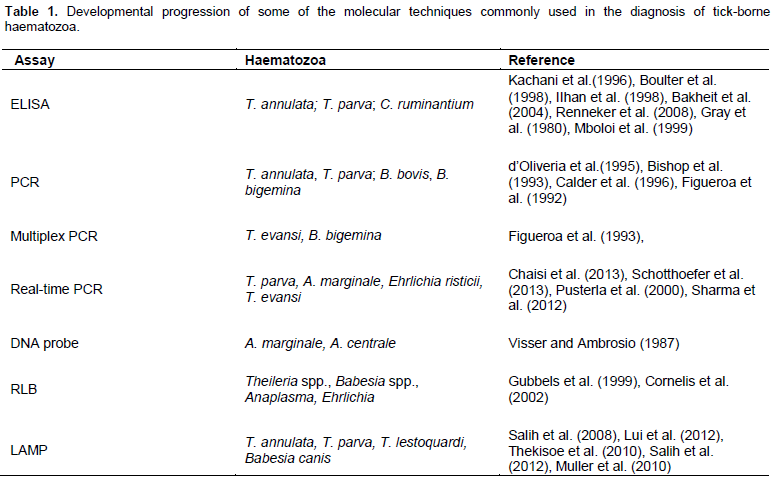
The microscopic techniques for diagnosis of tick-borne diseases are still considered as the “gold standard” technique. Microscopic examination shows Theileria schizonts in the lymph node smears and piroplasms alone or along with schizonts in blood smears (Figure 1).
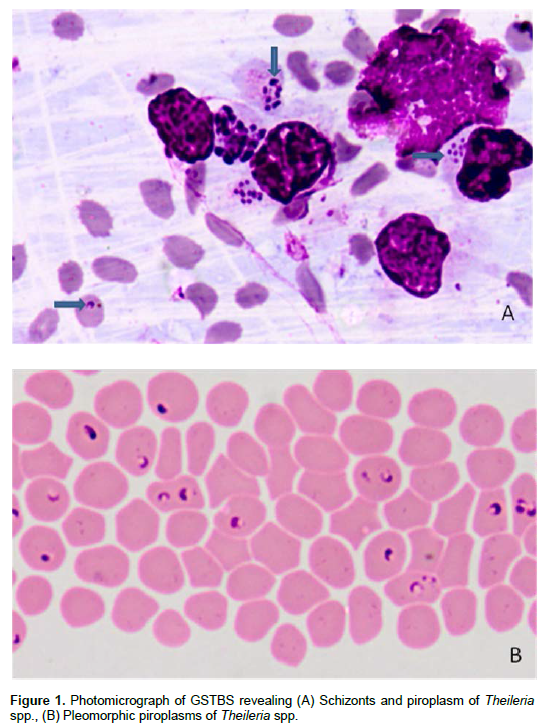
However, the detection of piroplasms in blood smears in the absence of clinical assessment and lymph node biopsy are difficult to interpret, since piroplasms of T. annulata, T. parva, Theileria mutans and T. lestoquardi can be found in the blood smears (Norval et al., 1992) which are difficult to differentiate based on the morpho-logy. It is also quite difficult, if not impossible, to demon-strate parasites in carrier animals as the numbers of pa-rasites in such animals fall below detectable levels soon after the acute stages of the disease (de Waal, 2012).
Babesiosis is diagnosed by examination of blood or organ smears stained with Romanowsky stain (Callow et al., 1993; Bose et al., 1995). The direct method involves identifying the parasite in the stained blood smears; however, this technique shows a low sensitivity in subclinical and chronic phase of the infection (Terkawi et al., 2011). Blood film examination requires very much expertise to differentiate between Babesia species from one or more animal species which look similar under stained preparation (Figure 2). Quantitative buffy coat (QBC), an acridine orange based improved technique, which concentrates and stains parasitized blood in one step (Levine et al., 1989) which may prove more valuable in low parasitaemic cases (Figure 3).

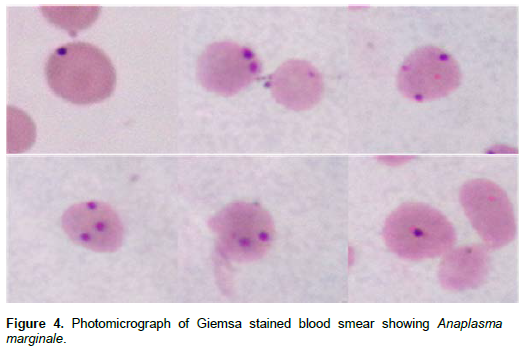
The most commonly used method for diagnosis of Anaplasma infection is the microscopic examination of Giemsa stained thin blood smears especially in clinical acute form of disease (Figure 4). The sensitivity of this method is 106 infected erythrocytes per milliliter of blood (Gale et al., 1996), but due to the low parasitemia in carrier cattle and difficulty to differentiate Anaplasma from other structures even by an experienced hand, this method is not recommended for the characterization of persistently infected cattle (Carelli et al., 2007). Sub-inoculation of A. marginale infected erythrocytes into susceptible splenectomized calves has been considered as the ’gold standard’ for detection of such cattle, but it is not practical for routine testing (Luther et al., 1980).
Owing to the rapid development of heartwater disease (Cowdriosis), the frequent lack of characteristic clinical signs, and sometimes even a total absence of lesions, the diagnosis of the disease in live animal is difficult (Camus et al., 1996). Tentative diagnosis relies on some clinical, epidemiological and macroscopic appearance of some organs. Differential diagnosis should consider diseases that cause central nervous system “CNS” manifestations. In dead animals, tentative diagnosis could be made by the presence of transudates in the pericardium and thorax at postmortem, but definitive diagnosis requires the demonstration of C. ruminantium in brain crush smears. This has been established much earlier and by the application of specific serological and molecular techniques in live animals.
The conventional microscopy for diagnosis is simple and does not require the purchase and maintenance of expensive equipments. However, limitations of con-ventional microscopy method include: (i) it is extremely labour intensive particularly when a large number of samples are to be examined in a short period; (ii) trained technicians are required for accurate diagnosis; (iii) not of much use in cases, when haemoparasites are morphologically similar or too small to be missed or infection is very low; (iv) in some cases culture of the parasite is needed, which may require specialized media, which is again time consuming.
INDIRECT DIAGNOSTIC METHODS
When parasites occur at densities below the sensitivity of direct method employed or cannot be directly demonstrated in a biological sample due to the life cycle in the host, in those cases indirect methods of diagnosis are used, which include serological tests either used for detection of antibodies or antigens. Among the various serological tests, most important once include comple-ment fixation test (CFT), indirect fluorescent antibody technique (IFAT) and enzyme-linked immunosorbent assay (ELISA).
CFT
CFT has been used for diagnosis of Babesia, Theileria,Toxoplasma, Trypanosoma, etc (Herr et al., 1985; Bose et al., 1995). Based on this test, commercial kit (COFEB Kit) has been developed for diagnosis of equine piroplasmosis (Sengupta, 2001).
IFAT
Indirect fluorescent antibody technique has been used since long for the diagnosis of parasites like Babesia spp. (Morzaria et al., 1977; Anderson et al., 1980), Theileria (Morzaria et al., 1977; Darghouth et al., 2004). IFA test has also been recommended by OIE as one of the diagnostic test for theileriosis.
ELISA
ELISA is increasingly being used for detection of parasite-specific antibodies, antigens and immune complexes (Kachani et al., 1992). ELISA based on schizont antigen functioned well in the case that cellular fraction was enriched from the soluble fraction (Manuja et al., 2000). On the other hand, ELISA for the diagnosis of Theileria spp. infection in sheep using piroplasm antigen obtained from experimentally infected sheep with para-sitemia reaching 30% was developed (Gao et al., 2002).
However, cross-reaction was found with B. ovis. This result may be questionable, because it could not be excluded that these sheep were already infected with both Theileria spp. and B. ovis.
In spite of relatively good performances of crude anti-gens, the disadvantages of this approach include the requirement for experimental animals for piroplasm antigen production and the batch-to-batch variation, beside the need to standardize protocols to obtain antigen from crude parasite material (Gubbels, 2000). These problems have been circumvented by use of several recombinant parasite antigens in ELISA. Two ELISAs based on recombinant proteins have been deve-loped. Firstly, the sporozoite antigen (SPAG-1) has been demonstrated to detect exposure to T. annulata, but sensitivity and specificity of this ELISA has not been evaluated (Boulter et al., 1998). Secondly, a merozoite surface antigen, Tams-1 has been tested as a candidate antigen for a diagnostic ELISA (Ilhan et al., 1998). More-over, an ELISA for detection of T. annulata infection was established and validated and applied for epidemiological studies in the field (Schnittger et al., 2002; Bakheit et al., 2004; Salih et al., 2005, 2007; Seitzer et al., 2007). An advance in serological diagnosis was achieved with the development of a competitive ELISA applying the TaSP antigen and using a monoclonal antibody (1C7) that was found to bind to TaSP antigen (Renneker et al., 2008). ELISA is widely used as the basis for epidemiological surveys (Passos et al., 1998) and for evaluation of vaccination programme (Guglielmone et al., 1997). Serological tests have the disadvantage of relying on the presence of specific antibodies, which may take days or weeks to develop in an infected animal or may persist for months after the infection has been cleared (Mosqueda et al., 2012).
Mboloi et al. (1999) investigated the major antigenic protein 1 fragment B (MAP1-B) in ELISA for the diagnosis of C. ruminantium infection. They concluded that the MAP1-B ELISA is a useful test for the diagnosis of C. ruminantium infection in small ruminants. However, this test is known to be less sensitive in cattle than in small ruminant (Knopf et al., 2002).
NUCLEIC ACID-BASED TECHNIQUES
Molecular nucleic acid-based diagnostic techniques have been developed for a number of haemoparasite. They have been proved to be sensitive, easy to use, can analyze large number of samples and can detect the parasites directly in clinical and environmental samples without culture (Weiss, 1995; Dey and Singh, 2009). The first introduced diagnostics was nucleic acid hybridization technique, however, in the recent years, PCR and allied techniques along with genomic sequencing have over taken it, and have become a driving force for the development of rapid, sensitive and specific assays capable of genomic detection. They are widely used because of being highly sensitive and can be performed rapidly in a cost effective manner. Various molecular diagnostic techniques that have been developed for diagnosis of parasites include conventional PCR, RAPD-PCR, RFLP-PCR, multiplex-PCR, real-time PCR, reverse transcriptase PCR, PCR-ELISA, micro-arrays, loop-mediated isothermal amplification (LAMP), etc.
PCR
Many authors discussed the use of molecular tools for the study of Theileria parasites, which affect ruminants, mostly bovines. Tanaka et al. (1993) utilized a probe derived from a gene encoding a 32 kDa intra-erythrocytic piroplasm surface protein of Theileria sergenti (Theileria orientalis). Their method was sufficiently sensitive to detect four parasites per microlitre of blood with a 10 µl sample. Detection of T. annulata in blood samples of carrier cattle using polymerase chain reaction (PCR) was reported (d’Oliveria et al., 1995). The assay employed primers specific for the gene encoding the 30 kDa major merozoite surface antigen of this species. This PCR assay was found to be highly specific and sensitive (three parasites per microlitre of blood). Allsop et al. (1993) developed another technique based on PCR using oligonucleotide probes, which detected small subunit ribosomal RNA sequences (srRNA). These probes were efficient in discriminating between six different tick-borne parasites harboured by cattle. Bishop et al. (1993) were able to further increase the accuracy of the identification of isolates or strains by using random amplified polymorphic deoxyribonucleic acid ‘DNA’ (RAPD). Moreover, several real-time PCR assay has been developed for diagnosis and quantitation of many tick-bone parasites (Dong et al., 2013; Schotthoefer et al., 2013; Bloch et al., 2013).
The sensitivity and specificity of molecular methods is very high and over the years a number of different approaches have been developed to detect Babesia spp. in the hosts and vectors. Deoxyribonucleic acid (DNA) probing was the first developed method, which was used to detect babesial DNA from parasitized blood (Buening et al., 1990). Figueroa et al. (1994) set up a PCR-based diagnostic assay to detect B. bovis in chronically infected cattle. The target sequence was a gene encoding a 60 kDa merozoite surface protein. The level of sensitivity was high as the PCR product was detected in blood samples containing approximately 20 µl of packed cell with a parasitemia of 0.000001%. Nested PCR (nPCR) has been effective for the detection of carrier animals infected with B. bigemina; the sensitivity was reported to be as low as one infected erythrocyte in 108 cells (Figueroa et al., 1992). Carson et al. (1994) used RAPD to reveal markers of species and isolates for B. bovis and B. bigemina. As mentioned earlier, RLB technique has proved highly sensitive and specific for the detection of these parasites (Gubbels et al., 1999). A hot-start PCR (semi-nested) to detect B. bovis, B. bigemina based
on aspartic proteinase babesipsin gene was developed and successfully applied (Martin et al., 2008; Awad et al., 2011). Multiplex PCR (Figueroa et al., 1993), real-time PCR (Buling et al., 2007) and multiplex LAMP (Iseki et al., 2007) are in process of development and validation for identification and characterization of B. bigemina.
Probes specific for A. marginale and A. centrale have been isolated (Visser and Ambrosio, 1987). These probes could detect 127 and 8 ng DNA of A. centrale and A. marginale, respectively. This probe could also be used on blood from field samples to detect parasiteamia in cattle. On the other hand, the A. marginale probes described by Goff et al. (1988) could detect 0.01 ng DNA, which is equivalent to a parasiteamia of 0.000025% (Eriks et al., 1989). This probe could also detect the presence of the parasite in the infected tick vector. Presently, knowledge about carrier state of anaplasmosis is so difficult to know the epidemiological status in enzootic regions. PCR based on msp4 gene for A. marginale and Anaplasma ovis (de la Fuente et al., 2001, 2003) have also been developed. Sequencing of gltA and ompA genes, identification of Rickettsia species based on the sizes of highly variable intergenic spacers, namely, dksA-xerC, mppA-purC, and rpmE-tRNAfMet was carried out. Application of multiplex PCR for simultaneous amplification of 3 spacers combined with capillary electrophoresis separation technique is simple, accurate, and high-throughput fragment sizing with considerable time and cost savings (Nakaoa et al., 2013). PCR base tests including PCR ELISA and duplex PCR have been developed and applied successfully with high sensitivity and specificity to differentiate tick borne haematozoan diseases (Galle et al., 1996; Ala and Wayne, 2005; Torina et al., 2008; Ashuma et al., 2013; Sharma et al 2013). Further, nested PCR devised for detection of A. marginale in cattle shows increased specificity and sensitivity (Ybanez et al., 2013). Recently, semi-quantitative multiplexed-tandem PCR for the detection and differentiation of four T. orientalis genotypes in cattle has been applied (Pereraa et al., 2014).
There are certain reservations to the routine use of such technique. First of all, contamination of the laboratory environment has to be rigorously controlled and this implies numerous controls for quality diagnosis. Suggestions for avoiding contamination have been presented by Altwegg (1995) and Carino and Lee (1995). The first suggestion relates to the organization of the laboratory space so as different stages of diagnosis are carried out in separate areas. The second suggestion relates to the use of Uracil DNA Glycosylase (UDG) for preamplification sterilization of the PCR product (Longo et al., 1990). For the detection of contamination, negative control must be realized at each step of the PCR preparation (Comes et al., 1995). PCR may also fail due to the inhibition of specific amplification (false negative). Thus, the use of positive control is necessary to increase confidence in negative PCR results. The cost of diagnosis is the second drawback of PCR. Although PCR identification costs are being progressively reduced, they remain higher than those of parasitological and immuno-logical techniques. The third drawback is the absence of quantification. Only limited attempt of using PCR has been conducted to date to estimate the intensity of parasitemia due tick-borne parasitic diseases.
RAPD-PCR
Random Amplification of Polymorphic DNA-PCR also known as AP-PCR (arbitrary primed PCR), in which primers of arbitrary sequences are used to amplify fragments of the genome. This technique is very simple, fast and does not require either prior knowledge of the DNA sequence or DNA hybridization. This technique has been employed to differentiate species of Leishmania and also to study polymorphisms of Plasmodium, Trypanosoma, etc (Hajjaran et al., 2004).
PCR-RFLP
PCR-Restriction Fragment Length Polymorphism is used for diagnosis of species and genotypes of parasites. It is performed by digesting the PCR products obtained from parasitic gene amplification, by restriction enzymes or endonucleases. These enzymes cut DNA into fragments of certain sizes, whose analysis on agarose or poly-acrylamide gel results in different patterns of fragment sizes, enabling the identification. Zaeemi et al. (2011) were able to differentiate among T. lestoquardi, Theileria ovis, and T. annulata in case of sheep. Recently, semi nested PCR-RFLP was used for detection of persistent anaplasmosis in tick infested cattle (Jaswal et al., 2014).
Multiplex PCR
Multiplex polymerase chain reaction is a modification of conventional polymerase chain reaction in order to rapidly detect deletions or duplications in a large gene in a single reaction. It is a variant of PCR which offers a significant advantage over single-plex PCR as this two or more target loci from one or more organisms are amplified using mixture of locus-specific primer pairs in a single reaction (Edwards and Gibbs, 1994; Markoulatos et al., 2002). Multiplex PCR had been employed in detection of concurrent infections in field for the detection of haemoprotozoans (Figueroa et al., 1998). Various duplex PCR for the detection of concurrent infections of economically important haemoprotozoans have also already been standardized in our laboratory with con-venient large scale field application (Sharma et al, 2013; Kaur et al., 2012).
Real-time PCR (RT-PCR)
This technique involves the analysis of genome using fluorogenic probes that release fluorescent signals during amplification. The advantages of real time PCR assay over conventional PCR are that it is relatively rapid and convenient because there is no need to perform gel electrophoresis to visualize the PCR products. Real-time PCR is a simple, fast, closed and automatized amplification system responsible for decreasing the risk of cross contamination. This technique has been used for the detection of a number of parasites including Leishmania, Plasmodium, Trypanosoma (Bell and Ranford-Cartwright, 2002; Gasser, 2006; Sharma et al., 2012) in various regions of the world. Real-time PCR has engendered wider acceptance of PCR due to its improved rapidity, sensitivity, reproducibility and the reduced risk of carryover contamination (Mackay, 2004). Jeong et al. (2003) applied real-time PCR for diagnosis and quantification of T. sergenti using specific primer for 33 kDa gene. A pan-Theileria FRET-qPCR that can detect all recognized Theileria spp. of ruminants in a single reaction has also been developed (Yang et al., 2014).
PCR-ELISA
By PCR ELISA, the sensitivity of ELISA and specificity of PCR are combined and used for the detection of parasitic genome. The PCR products are hybridized to an immobilized capture probe. The assay thus measures sequences internal to the PCR product and is a less expensive assay and can be an alternative to real time PCR. This ELISA is useful for detecting and differen-tiating between multiple targets. This technique has been used in detection and quantification of Trypanosoma evansi in animals and vectors (Chansiri et al., 2002). The sensitivity limit of PCR-ELISA was 0.01 pg, which corresponded to one parasite/ml of blood. No cross-reactivity of the assay was observed against B. bovis, B. bigemina, A. marginale, Theileria spp. and host DNA (Chansiri et al., 2002).
RLB
Two integrated approaches were developed to detect several Theileria or Babesia spp. in one assay (Figueroa et al., 1993; Allsop et al., 1993). Using these approaches, multiple species can be detected in one assay without performing independent PCR reactions for each parasite (Gubbels et al., 1999). One of such techniques, reverse line blot (RLB) hybridization, combines a genus specific PCR with hybridization to membrane bound type/species-
specific oligonucleotide for differential detection. This technique can differentiate all known Theileria and Babesia spp. of importance in cattle in the sub-tropics on the basis of their differences in 18S subunit rRNA gene sequences (Gubbels et al., 1999). The specificity of the techniques result from the fact that amplified conserved domains of the 18 srRNA genes of the parasites are hybridized to species specific oligonucleotide immobilized on a solid membrane.
LAMP
Recently, a rapid, simple, and sensitive technique, loop mediated isothermal amplification (LAMP), was deve-loped (Notomi et al., 2000). This is a novel strategy for gene amplification which relies on the auto-cycling strand displacement synthesis of target deoxyribonucleic acid (DNA) by Bst DNA polymerase under isothermal con-ditions. Further improvement of the technique has been achieved by the use of additional loop primers, which increased its efficiency and rapidity (Nagamine et al., 2002). The LAMP technique allows visual detection of amplified products through the addition of fluorescent dyes such as SYBR Green (Poon et al., 2006) and measurement of turbidity (Mori et al., 2001). Unlike PCR, LAMP is carried out at a temperature range of 60 to 65°C eliminating the need of a thermal cycler. In addition, the reaction can be carried out without the need of DNA extraction. The method has been successfully developed for the detection of several TBDs (Salih et al., 2008; Liu et al., 2008; Muller et al., 2010; Thekisoe et al., 2010;Salih et al., 2012).
DNA Microarrays
The technology commonly known as gene chip, DNA chip, or biochip was originally developed for mapping of genes being used to detect a wide variety of pathogens through multi-gene detection. The microarrays consist of solid supports like glass slide or silicon chip or nylon membrane, onto which the nucleic acid sequences from thousands of different genes are attached at fixed locations. The main advantage of this technique is that it combines DNA amplification strategies with subsequent hybridization to oligonucleotide probes specific for multiple target sequences. Secondly, it allows for the simultaneous analysis of a larger number of genetic features in a single trial. However, the technique is costly. This technique has been used in detection and genotyping of vector-borne parasites of medical impor-tance like Leishmania, Plasmodium, Toxoplasma, and Trypanosoma (Duncan, 2004).
CONCLUSION
Diagnosis of haemo-parasitic infection has been largely based on clinical symptoms and identifying/ demonstrating the causative agent by parasitological techniques. Although, microscopy is still considered as a gold standard in the diagnosis of many parasitic diseases, it cannot be applied to all situations particularly where the diagnostic requirements demand defining the carrier status. Although, the use of various serological methods provide definite clues about the parasitic infection in general, but these tests have some limitations. In this direction, the recently introduced molecular biological techniques will certainly be of help, though presently their use on large scale is mainly limited to large parasitology laboratories. The use of DNA hybridization probes, although developed several years ago, has never been developed to suite regular diagnostic laboratories and has now been super ceded by polymerase chain reaction. The ability of PCR to detect very small quantities of a target material and the absence of the need to use radioactive elements are two of the advantages of PCR compared with hybridization techniques. However, more accurate identification of a PCR product may require the use of specific nucleic acid probes. But, it is not evident, with exception of RLB which is now being commercially produced, that the use of the technique will spread as a routine diagnostic tool in the laboratories. The use of molecular biology tools based on nucleic acid for tick-borne diseases will therefore, for sometime continue to be used in research activities rather than for day-to-day diagnosis in the laboratories. However, recombinant antigens based ELISAs may be available for routine diagnosis in the field.
CONFLICT OF INTEREST
Authors have no conflict of interest.
REFERENCES
| Ala L, Wayne J (2005). Molecular approaches to detect and study the organisms causing bovine tick borne diseases: babesiosis and anaplasmosis. Afr. J. Biotechnol. 4:292-302. | ||||
|
Allsop BA, Baylis HA, Allsop MT, Cavaher-smith T, Bishop RP, Camngton DM, Sohanpal B, Spooner P (1993). Discrimination between six species of Theileria using oligonucleotide probes which detected small subunit ribosomal RNA sequences. Parasitology 107:157-165. Crossref |
||||
|
Altwegg M (1995). General problems associated with diagnostic appli-cations of amplification methods. J. Microbiol. Methods 23:21-30. Crossref |
||||
|
Ambrosio RE, Du Plessis JL, Bezuidenout JD (1987). The construction of genomic libraries of Cowdria ruminantium in an expression vector λgt11. Onderstepoort J. Vet. Res. 54:255-256. Pubmed |
||||
|
Anderson JF, Magnarelli LA and Sulzer AJ. (1980). Canine babesiosis: Indirect fluorescent antibodies test for a North American isolate of Babesia gibsoni. Am. J. Vet. Res. 41:2102-2105. Pubmed |
||||
|
Ashuma, Sharma A, Singla LD, Kaur P, Bal MS, Batth BK, Juyal PD (2013). Prevalence and haemato-biochemical profile of Anaplasma marginale infection in dairy animals of Punjab (India). Asian Pac. J. Trop. Med. 6:139-144. Crossref |
||||
|
Awad H, Sandra Antunes S, Galindo RC, do Rosário VE, de la Fuente J, Domingos A, El Hussein AM (2011). Prevalence and genetic diversity of Babesia and Anaplasma species in cattle in Sudan. Vet. Parasitol. 181:146-152. Crossref |
||||
|
Bakheit MA, Schnittger L, Salih DA, Boguslawski K, Beyer D, Fadl M, Ahmed JS (2004). Application of the recombinant Theileria annulata surface protein in an indirect ELISA for the diagnosis of tropical theileriosis. Parasitol. Res. 92:299-302. Crossref |
||||
|
Bell AS, Ranford-Cartwright LC (2002). Real-time quantitative PCR in parasitology. Trends Parasitol. 18:338-342. Crossref |
||||
|
Bishop R, Musoke A, Skilton R, Morzaria S, Garder M, Nene V (2008). Theileria: life cycle stages associated with the ixodid tick vector. In: Alan S. Bowman, Patricia A. Nuttall (eds.), Ticks: Biology, Disease and Control. Cambridge University Press. pp. 308-324 Crossref |
||||
|
Bishop R, Sohanpal B, Morzaria S (1993). Theileria parva: Detection of genomic polymorphisms by polymerase chain reaction amplification of DNA using arbitrary primers. Exp. Parasitol. 77:53-61. Crossref |
||||
|
Bloch EM, Lee TH, Krause PJ, Telford SR, Montalvo L, Chafets D, Usmani-Brown S, Lepore TJ, Busch MP (2013). Development of a real-time polymerase chain reaction assay for sensitive detection and quantitation of Babesia microti infection. Transfusion 53: 2299-2306. Pubmed |
||||
|
Bose R, Jorgensen WK, Dalgliesh RJ, Friedhoff KT, de Vos AJ (1995). Current state and future trends in the diagnosis of babesiosis. Vet. Parasitol. 57:61-74. Crossref |
||||
|
Boulter NR, Brown CGD, Kirvar E, Glass E, Campbell J, Morzaria S, Nene V, Musoke A, d'Oliveria C, Gubbels MJ, Jongejan F, Hall R (1998). Different vaccine strategies used to protect against Theileria annulata. Ann. N. Y. Acad. Sci. 849:234-246. Crossref |
||||
|
Buening G M, Barbet A, Myler P, Mahan S, Nene V, McGuire TC (1990). Characterisation of a repetitive DNA probe for Babesia bigemina. Vet. Parasitol. 36:11-20. Crossref |
||||
|
Buling A, Criado-Fornelio A, Asenzo G, Benitez D, Barba-Carretero JC, Florin CM (2007). A quantitative PCR assay for the detection and quantification of Babesia bovis and Babesia bigemina. Vet. Parasitol. 147:16-25. Crossref |
||||
|
Calder JA, Reddy GR, Chieves L, Courtney CH, Littell R, Livengood JR, Norval RA, Smith C, Dame JB (1996). Monitoring Babesia bovis infections in cattle by using PCR-based tests. J. Clin. Microbiol. 34:2748-2755. Pubmed |
||||
| Callow LL, Rogers RJ, de Vos AJ (1993). Tick-borne diseases: cattle pathology and serology. In: Corner LA, Bagust TJ (eds.), Australian Standard Diagnostic Techniques for Animal Diseases. CSIRO Information Services, East Melbourne. pp. 1-16. | ||||
| Camus E, Barre N, Martinez D, Uilenberg G (1996). Heartwater (Cowdriosis). A review, 2nd ed. Office International des Epizooties, Paris, France. | ||||
|
Carelli G, Decaro N, Lorusso A, Elia G, Lorusso E, Mari V, Ceci L, Buonavoglia C (2007). Detection and quantification of Anaplasma marginale DNA in blood samples of cattle by real-time PCR. Vet. Microbiol. 124:104-107. Crossref |
||||
|
Carino JJ, Lee HH (1995). Nucleic acid amplification methods. J. Microbiol. Methods 23:3-20. Crossref |
||||
|
Carson CA, Brandt HM, Jensen JB, Bailey CW, Allen GK (1994). Use of the random amplified polymorphic DNA analysis to compare Babesia bovis and B. bigemina. Parasitol. Res. 80:312-315. Crossref |
||||
|
Chaisi ME, Janssens ME, Vermeiren L, Oosthuizen MC, Collins NE, Geysen D (2013). Evaluation of a real-time PCR Test for the detection and discrimination of Theileria Species in the African Buffalo (Syncerus caffer). PLoS One 17:e75827. Crossref |
||||
|
Chansiri K, Khuchareontaworn S, Sarataphan N (2002). PCR-ELISA for diagnosis of Trypanosoma evansi in animals and vector. Mol. Cell. Probes 16:173-177. Crossref |
||||
| Comes AM, Humbert JF, Carbaret J, Elard L (1995). Using molecular tools for diagnosis in veterinary parasitology. Vet. Parasitol. 27:333-342. | ||||
|
Cornelis PJB, Vos de S, Taoufik A, Olivier AES, Jongejan F (2002). Simultaneous detection of Anaplasma and Ehrlichia species in ruminants and detection of Ehrlichia ruminantium in Amblyomma variegatum ticks by reverse line blot hybridization. Vet. Microbiol. 89:223-238. Crossref |
||||
| d'Oliveria C, Weide M, Habela MA, Jacquiet P, Jongejan F (1995). Detection of Theileria annulata in blood samples of carrier cattle by PCR. J. Clin. Microbiol. 33:2665-2669. | ||||
|
de la Fuente J, Van Den Bussche RA, Kocan KM (2001). Molecular phylogeny and biogeography of North American strains of Anaplasma marginale (Rickettsiaceae: Ehrlichieae). Vet. Parasitol. 97:65-76. Crossref |
||||
| de la Fuente J, Van Den Bussche RA, Prado T, Kocan KM (2003). | ||||
|
Anaplasma marginale major surface protein 1a genotypes evolved under positive selection pressure but are not markers for geographic strains. J. Clin. Microbiol. 41:1609-1616. Crossref |
||||
|
de Waal T (2012). Advances in diagnosis of protozoan diseases. Vet. Parasitol. 189:65-74. Crossref |
||||
| Dey A, Singh S (2009). Progress of science from microscopy to microarrays (Part 1): Diagnosis of parasitic diseases. J. Lab. Phys. 1: 2-6. | ||||
|
Dong T, Qu Z, Zhang L (2013). Detection of A. phagocytophilum and E. chaffeensis in patient and mouse blood and ticks by a duplex real-time PCR assay. PLoS One 8: e74796. Crossref |
||||
|
Duncan R (2004). DNA microarray analysis of protozoan parasite gene expression: outcomes correlate with mechanisms of regulation. Trends Parasitol. 20:211-216. Crossref |
||||
|
Edwards MC, Gibbs RA (1994). Multiplex PCR: advantages, development, and applications. PCR Methods Appl. 3:S65-S75. Crossref |
||||
| Eriks I, Palmer G, Mc Guire T, Allred D, Barbet T (1989). Detection and quantitation of Anaplasma marginale in carrier cattle using a nucleic acid probe. J. Clin. Microbiol. 27:206-212. | ||||
|
Figueroa JV, Chieves LP, Johnson GS, Buening GM (1992). Detection of Babesia bigemina infected carriers by polymerase chain reaction amplification. J. Clin. Microbiol. 30:2576-2582. Pubmed |
||||
|
Figueroa JV, Buening GM (1995). Nucleic acid probes as diagnostic method for tick-borne hemoparasites of veterinary importance. Vet. Parasitol. 57:75-92. Crossref |
||||
|
Figueroa JV, Chieves LP, Johnson GS (1994). Polymerase chain reaction-based diagnostic assay to detect cattle chronically infected with Babesia bovis. Rev. Lantinoam Microbiol. 36:47-55. Pubmed |
||||
|
Figueroa JV, Chieves LP, Johnson GS, Buening GM (1993). Multiplex polymerase chain reaction based assay for detection of Babesia bigemina, B. bovis and Anaplasma marginale DNA in bovine blood. Vet. Parasitol. 50:69-81. Crossref |
||||
|
Figueroa JV, Alvarez JA, Ramos JA, Rojas EE, Santiago C, Mosqueda JJ, Vega CA, Buening GM (1998). Bovine babesiosis and anaplasmosis follow-up on cattle relocated in an endemic area for hemoparasitic diseases. Ann. N. Y. Acad. Sci. 849:1-10. Crossref |
||||
|
Gale KR, Dimmock CM, Gartside M, Leatch G. (1996). Anaplasma marginale: detection of carrier cattle by PCR ELISA. Int. J. Parasitol. 26:1103-1109. Crossref |
||||
|
Gao YL, Yin H, Luo JX, Ouyang WQ, Boa HM, Guan GQ, Zhang QC, Lu WS, Ma LM (2002). Development of enzyme-linked immunosorbent assay for the diagnosis of Theileria sp. infection in sheep. Parasitol. Res. 88:s8-s10. Crossref |
||||
|
Gasser RB. (2006). Molecular tools - advances, opportunities and prospects. Vet. Parasitol. 136(2):69-89. Crossref |
||||
|
Goff W, Barbet AF, Stiller D, Palmer G, Knowles D, Kocan KM, Gorham J, Mc Guire TC (1988). Detection of Anaplasma marginale infected tick vector by using cloned DNA probes. Proc. Natl Acad. Sci. USA 85:919-923. Crossref |
||||
|
Gray MA, Luckins AG, Rae PF, Brown CGD (1980). Evaluation of an enzyme immunoassay for serodiagnosis of infections with Theileria parva and Theileria annulata. Res. Vet. Sci. 29:360-366. Pubmed |
||||
|
Guan G, Moreau E, Liu J, Hao X, Ma M, Luo J, Chauvin A, Yin H (2010). Babesia sp. BQ1 (Lintan): molecular evidence of experimental transmission to sheep by Haemaphysalis qinghaiensis and Haemaphysalis longicornis. Parasitol. Int. 59:265–267. Crossref |
||||
| Gubbels JM (2000). Molecular characterization and diagnosis of Theileria annulata and Theileria buffeli. PhD. Thesis, Utrecht University, The Netherlands. | ||||
|
Gubbels JM, de Vos AP, van der Weide M, Viseras J, Schouls LM, de Vries E, Jongejan F (1999). Simultaneous detection of bovine Theileria and Babesia species by reverse line blot hybridization. J. Clin. Microbiol. 37:1782-1789. Pubmed |
||||
|
Guglielmone AA, Lugaresi CI, Volpogni MM, Anziani OS, Vanzini VR (1997). Babesia antibody dynamics after cattle immunization with live vaccines, measured with an indirect immunofluorescence test. Vet. Parasitol. 70:33-39. Crossref |
||||
| Hajjaran H, Mohebali M, Razavi MR, Rezaei S, Kazemi B and Edrissian GH. (2004). Identification of Leishmania species isolated from human cutaneous leishmaniasis using Random Amplified Polymorphic DNA (RAPD-PCR). Iranian J. Public Health 33:8-15. | ||||
|
Herr S, Huchzermeyer HF, Te Brugge LA, Williamson CC, Roos JA, Schiele GJ. (1985). The use of a single complement fixation test technique in bovine brucellosis, Johne's disease, dourine, equine piroplasmosis and Q-fever serology. Onderstepoort J. Vet. Res. 52:279-282. Pubmed |
||||
|
Ilhan T, Williamson S, Kirvar E, Shiels B, Brown CGD (1998). Theileria annulata: Carrier state and immunity. Ann. N. Y. Acad. Sci. 849:109-125. Crossref |
||||
|
Iseki H, Alhassan A, Ohta N, Thekisoe OM, Yokoyama N, Inoue N, Nambota A, Yasuda J, Igarashi I (2007). Development of a multiplex loopmediated isothermal amplification (mLAMP) method for the simultaneous detection of bovine Babesia parasites. J. Microbiol. Methods 71:281-287. Crossref |
||||
| Jaswal H, Bal MS, Singla LD, Amrita, Kaur P, Mukhopadhyay, Juyal PD (2014). Application of msp1β PCR and 16S rRNA semi nested PCR-RFLP for detection of persistent anaplasmosis in tick infested cattle. Int. J. Adv. Res. 2:188-196. | ||||
|
Jeong W, Hweon CH, Kang SW, Paik SG (2003). Diagnosis and quantification of Theileria sergenti using TaqMan PCR. Vet. Parasitol. 111:287-295. Crossref |
||||
|
Jongejan F, Uilenberg G (1994). Ticks and control methods. Rev. Sci. Tech. 13: 1201-1226. Pubmed |
||||
|
Kachani M, Flach EJ, Williamson S, Ouhelli H, El Hasnaoui M, Spooner RL (1996). The use of an enzyme-linked immunosorbent assay for tropical theileriosis research in Morocco. Prev. Vet. Med. 26:329-339. Crossref |
||||
|
Kachani M, Oliver RA, Brown CGD, Ouhelli H, Spooner RL (1992). Common and stage-specific antigens of Theileria annulata. Vet. Immunol. Immunopathol. 34:221-234. Crossref |
||||
| Kaur P, Sharma A, Singla LD, Juyal PD (2012). Molecular detection of anaplasmosis and babesiosis by duplex PCR in cattle. Crop Improv. 1395-1396. | ||||
|
Knopf L, Komoin-Oka C, Betschart B, Jongejan F, Gotstein B, Zinsstag J (2002). Seasonal epidemiology of ticks and aspects of Cowdriosis in N′dama village cattle in Central Guinea Savannah of Cote ÄIvoire. Prev. Vet. Med. 53:21-30. Crossref |
||||
|
Levine RA, Wardlaw SC, Patton CL (1989). Detection of haematoparasites using quantitative buffy coat analysis tubes. Parasitol. Today 5:132-33. Crossref |
||||
|
Liu Z, Hou J, Bakheit MA, Salih DA, Luo J, Yin H, Ahmed JS, Seitzer U (2008). Development of loop-mediated isothermal amplification (LAMP) assay for rapid diagnosis of ovine theileriosis in China. Parasitol. Res. 103:1407-1412. Crossref |
||||
|
Liu A, Guan G, Du P, Liu Z, Gou H, Liu J, Yang J, Li Y, Ma, M, Niu Q, Ren Q, Bai Q, Yin H, Luo J (2012). Loop-mediated isothermal amplification (LAMP) assays for the detection of Theileria annulata infection in China targeting the 18S rRNA and ITS sequences. Exp. Parasitol.131:125-129. Crossref |
||||
|
Longo MC, Berninger MS, Hartley JL (1990). Use of the uracil DNA glycosylase to control carry-over contamination in polymerase chain reaction. Gene 93:125-128. Crossref |
||||
|
Luther DG, Cox HU, Nelson WO (1980). Comparisons of serotests with calf inoculations for detection of carriers in anaplasmosis-vaccinated cattle. Am. J. Vet. Res. 41:2085-2086. Pubmed |
||||
|
Mackay IM (2004). Real time PCR in the microbiology laboratory. Clin. Microbiol. Infect. Dis. 10:190-212. Crossref |
||||
|
Manuja A, Nichani AK, Kumar R, Rakha NK, Kumar B, Sharma KD (2000). Comparison of cellular schizont, soluble schizont and soluble piroplasm antigens in ELISA for detecting antibodies against Theileria annulata. Vet. Parasitol. 87:93-101. Crossref |
||||
|
Markoulatos P, Siafakas N, Moncany M (2002). Multiplex polymerase chain reaction: a practical approach. J. Clin. Lab. Anim. 16:47-51. Crossref |
||||
|
Martin TM, Pedro OC, Caldeira RA, do Rosário VE, Neves L, Domingos A (2008). Detection of bovine babesiosis in Mozambique by a novel seminested hot-start PCR method. Vet. Parasitol. 153:225-230. Crossref |
||||
|
Mboloi MM, Bekker CPJ, Kruitwager C, Griener M, Jongejan F (1999). Validation of indirect MAP-1B enzyme-linked immunosorbent assay for diagnosis of experimental C. ruminantium infection in small ruminants. Clin. Diagn. Lab. Immunol. 6:66-72. Pubmed |
||||
| Minjaw B, McLeod A (2003). Tick-borne diseases and poverty: the impact of ticks and tick-borne disease on the livelihoods of small-scale and marginal livestock owners in India and eastern and southern Africa. Research report, Department for International Development Animal Health Programme. Centre of Tropical Veterinary Medicine, University of Edinburgh, Scotland. P 116. | ||||
|
Mori Y, Nagamine K, Tomita N, Notomi T (2001). Detection of loop mediated isothermal amplification reaction by turbidity derived from magnesium pyrophosphate formation. Biochem. Biophys. Res. Commun. 289:150-154. Crossref |
||||
|
Morzaria SP, Brockelsby DW, Harradine DL (1977). Evaluation of the indirect fluorescent antibody test for Babesia major and Theileria mutans in Britain. Vet. Rec. 100:484-487. Crossref |
||||
|
Mosqueda J, Olvera-Ramírez A, Aguilar-Tipacamú G, Cantó GJ (2012). Current Advances in Detection and Treatment of Babesiosis. Curr. Med. Chem. 19:1504-1518. Crossref |
||||
|
Muller H, Aysu N, Liu Z, Salih DA, Karagenc T, Beyer D, Kullmann B, Ahmed JS, Seitzer U (2010). Development of a Loop-mediated Isothermal Amplification (LAMP) Assay for Rapid Diagnosis of Babesia canis infections. Transbound. Emerg. Dis. 57:63-65. Crossref |
||||
|
Muramatsu Y, Ukegaua S, El Hussein AM, (2005). Ehrlichia ruminantium in Sudan. Emerg. Inf. Dis.11:1792-1793 Crossref |
||||
|
Nagamine K, Hase T, Notomi T (2002). Accelerated reaction by loop mediated isothermal amplification using loop primers. Mol. Cell. Probes 16:223-229 Crossref |
||||
|
Nakaoa R, Qiu Y, Igarashi M, Magona JW, Zhou L, Ito K, Sugimoto C (2013). High prevalence of spotted fever group rickettsiae in Amblyomma variegatum from Uganda and their identification using sizes of intergenic spacers. Ticks Tick-borne Dis. 4:506-512. Crossref |
||||
|
Nantulya VM (1994). Suratex: a simple latex agglutination antigen test for diagnosis of Trypanosoma evansi infections (surra). Trop. Med. Parasitolol. 45:9-12. Pubmed |
||||
| Norval RAI, Perry BD, Young AS (1992). The epidemiology of theileriosis in Africa. Academic Press. pp. 749-4, 481. | ||||
|
Notomi T, Okayama H, Masubuchi H, Yonekawa T, Watanabe K, Amino N, Hase T (2000) Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 28:E63. Crossref |
||||
| OIE (2008). Bovine Anaplasmosis. In: "Terrestrial Manual". Office International des Epizooties. World Health Organization for Animal Health, Paris. Vol. 1 pp. 599-10. | ||||
|
Passos LM, Bell-Sakyi L, Brown CG (1998). Immunochemical characterization of in vitro culture derived antigens of Babesia bovis and Babesia bigemina. Vet. Parasitol. 76:239-249. Crossref |
||||
| Perry BD, Randolph TF, Mcdermott JJ, Sones KR, Thornton PK (2002). Investing in animal health research to alleviate poverty. International Livestock Research Institute, Nairobi, Kenya. P 138. | ||||
|
Poon LL, Wong BW, Ma EH, Chan KH, Chow LM, Abeyewickreme W, Tangpukdee N, Yuen KY, Guan Y, Looareesuwan S, Peiris JS (2006). Sensitive and inexpensive molecular test for falciparum malaria: detecting Plasmodium falciparum DNA directly from heat-treated blood by loop-mediated isothermal amplification. Clin. Chem. 52:303-306. Crossref |
||||
| Pereraa PK, Gassera RB, Firestone SM, Smith L, Roeber F, Abdul J (2014). Semi-quantitative Multiplexed-Tandem PCR for the Detection and Differentiation of Four Theileria orientalis Genotypes in Cattle. J. Clin. Microbiol. JCM.02536-14. | ||||
|
Pusterla N, Leutenegger CM, Sigrist B, Lutz H, Madigan JE. (2000). Detection and quantitation of Ehrlichia risticii genomic DNA in infected horses and snails by real-time PCR. Vet. Parasitol. 90:129-35. Crossref |
||||
|
Renneker S, Kullmann B, Gerber S, Dobschanski J, Bakheit MA, Geysen D, Shiels B, Tait A, Ahmed JS, Seitzer U (2008). Development of a Competitive ELISA for Detection of Theileria annulata Infection. Transbound. Emerg. Dis. 55:249-256. Crossref |
||||
|
Salih DA, Ahmed JS, Bakheit M, Ali EB, El Hussein AM, Hassan SM, Shariff OE, Fadl M, Jongejan F (2005). Validation of the indirect TaSP enzyme-linked immunosorbent assay for diagnosis of natural Theileria annulata infection in cattle. Parasitol. Res. 97:303-308. Crossref |
||||
|
Salih DA, Ali, AM, Liu Z, Bakheit MA, Taha KM, EL Imam AH, Kullmann B, El Hussein AM, Ahmed JS, Seitzer U (2012). Development of a loop-mediated isothermal amplification method for detection of Theileria lestoquardi. Parasitol. Res. 110:533-538. Crossref |
||||
|
Salih DA, El Hussein AM, Kyule MN, Zessin KH, Ahmed JS, Seitzer U, (2007). Determination of potential risk factors associated with Theileria annulata and Theileria parva infections of cattle in the Sudan. Parasitol. Res. 101:1285-1288. Crossref |
||||
|
Salih DA, Liu Z, Bakheit MA, Ali, AM, El Hussein AM, Unger H, Viljoen G, Seitzer U, Ahmed JS (2008). Development and evaluation of a loop-mediated isothermal amplification method for diagnosis of tropical theileriosis. Transbound. Emerg. Dis. 55:238–243. Crossref |
||||
|
Schnittger L, Katzer F, Biermann R, Shayan P, Boguslawski K, Mckellar S, Beyer D, Shiels BR, Ahmed JS (2002). Characterization of a polymorphic Theileria annulata surface protein (TaSP) closely related to PIM of Theileria parva: implications for use in diagnostic tests and subunit vaccines. Mol. Biochem. Parasitol. 120:247-256. Crossref |
||||
|
Schotthoefer AM, Meece JK, Ivacic LC, Bertz PD, Zhang K, Weiler T, Uphoff TS, Fritsche TR (2013). Comparison of a real-time PCR method with serology and blood smear analysis for diagnosis of human anaplasmosis: importance of infection time course for optimal test utilization. J. Clin. Microbiol. 51:2147-2153. Crossref |
||||
|
Seitzer U, Bakheit MA, Salih DA, Ali A, Haller D, Yin H, Schnittger L, Ahmed JS (2007). From molecule to diagnostic tool: Theileria annulata surface protein TaSP. Parasitol. Res. 101:S217–S223. Crossref |
||||
| Sengupta PP. (2001). Complement fixation test based COFEB- kit for the diagnosis of Babesia equi infection in equines. Patent application No. 36/DEL/2001, dt. 19.01.2001). Patent related to COFEB kit (56/DEL/2004) | ||||
|
Sharma A, Singla LD, Tuli A, Kaur P, Batth BK, Javed M, Juyal PD (2013). Molecular prevalence of Babesia bigemina and Trypanosoma evansi in dairy animals from Punjab, India by duplex PCR: A step forward to detection and management of concurrent latent infections. Biomed Res. Int. Article ID 893862:8 p Crossref |
||||
|
Sharma P, Juyal PD, Singla LD, Chachra D, Pawar H (2012). Diagnosis of Trypanosoma evansi in cattle and buffaloes by employing real time PCR using TaqMan assay. Vet. Parasitol. 190:375-382. Crossref |
||||
|
Tanaka M, Onoe S, Matsuba T, Katayama S, Yamanaka M, Yonemichi H, Hiramatsu K, Baeck B, Sugimoto C, Onuma M (1993). Detection of Theileria sergenti infection in cattle by polymerase chain reaction amplification of parasite-specific DNA. J. Clin. Microbiol. 31:2565-2569. Pubmed |
||||
|
Terkawi MA, Thekisoe OM, Katsande C, Latiff AA, Mans BJ, Matthee O, Mkize N, Mabogoane N, Marais F, Yokoyama N, Xuan X, Igarashi I (2011). Serological survey of Babesia bovis and Babesia bigemina in cattle in South Africa. Vet. Parasitol. 182:337-342. Crossref |
||||
|
Thekisoe OM, Rambritch NE, Nakao R, Bazie RS, Mbati P, Namangala B, Malele I, Skilton RA, Jongejan F, Sugimoto C, Kawazu S, Inoue N (2010). Loop-mediated isothermal amplification (LAMP) assays for detection of Theileria parva infections targeting the PIM and p150 genes. Int. J. Parasitol. 40:55-61 Crossref |
||||
|
Torina A, Alongi A, Naranjo V, Estrada-Pe-a A, Vicente J, Scimeca S, (2008). Prevalence and genotypes of Anaplasma species and habitat suitability for ticks in a Mediterranean ecosystem. Appl. Environ. Microbiol.74:7578-7584. Crossref |
||||
|
Uilenberg G (2006). Babesia—a historical overview. Vet. Parasitol. 138:3-10. Crossref |
||||
|
Visser ES, Ambrosio RE (1987). DNA probes for the detection of Anaplasma centrale and Anaplasma marginale. Onderstepoort J. Vet. Res. 54:623-627. Pubmed |
||||
|
Weiss JB (1995). DNA probes and PCR for diagnosis of parasitic infections. Clin. Microbiol. Rev. 8:113-130. Pubmed |
||||
|
Yang Y, Mao Y, Kelly P, Yang Z, Luan L, Zhang J, Li J, El-Mahallawy HS, Wang C (2014). A pan-Theileria FRET-qPCR survey for Theileria spp. in ruminants from nine provinces of China. Parasit. Vectors 7:413. Crossref |
||||
|
Ybanez AP, Sivakumar T, Ybanez RD, Ratilla JC, Perez ZO, Gabotero SR, Hakimi H, Kawazu S, Matsumoto K, Yokoyama N, Inokuma H (2013). First Molecular Characterization of Anaplasma marginale in Cattle and Rhipicephalus (Boophilus) microplus Ticks in Cebu Philippines. J. Vet. Med. Sci. 75:27-36. Crossref |
||||
|
Yin H, Schnittger L, Luo J, Seitzer U, Ahmed JS (2007). Ovine theileriosis in China: a new look at an old story. Parasitol. Res. 101: S191-S195. Crossref |
||||
|
Zaeemi M, Haddadzadeh H, Khazraiinia P, Kazemi B, Bandehpour M (2011). Identification of different Theileria species (Theileria lestoquardi, Theileria ovis, and Theileria annulata) in naturally infected sheep using nested PCR-RFLP. Parasitol. Res. 108:837-843. Crossref |
||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


