Full Length Research Paper

ABSTRACT
This study is aimed at evaluating antibody responses, viral loads, viral clearance and growth rate of Tanzanian free-range local chicken (FRLC) challenged with LaSota strain of Newcastle disease virus (NDV) as indicator traits for selection of chickens for breeding with enhanced resistance to the disease and economic value. Three popular free-range local chicken ecotypes: Kuchi, Ching’wekwe and Morogoro-medium from three ecological zones of Tanzania were used for the experiments. Progenies from the breeder chickens were challenged with 107 titer of 50% egg infectious dose (EID50) of the virus at 28 days of age. The viral loads and viral clearance rates evaluated by qRT-PCR from tear samples collected at 2- and 6-days post infection (dpi) showed that Kuchi could clear NDV better than Morogoro-medium and Ching’wekwe. Anti-NDV antibody levels determined from blood samples collected at 10 dpi using ELISA showed that Kuchi ecotype expressed higher mean anti-NDV antibodies compared to Morogoro-medium and Ching’wekwe. Growth rates determined from body weights collected for 38 days from day of hatch (D0) to 10 dpi showed higher growth rate for Kuchi than Morogoro-medium and Ching’wekwe chickens. Kuchi chickens were potentially more resistant to ND compared to Morogoro-medium and Ching’wekwe.
Key words: Free-range local chickens, Newcastle disease, immune response, innate resistance.
INTRODUCTION
Newcastle disease (ND) is one of the major devastating diseases in poultry worldwide (Ferreira et al., 2019; Miller and Koch, 2013). The disease is endemic in Tanzania and frequently causes outbreaks in free-range local chicken (FRLC) flocks. FRLCs in Tanzania are raised in the extensive management system mainly practiced in rural areas which exposes them to many risk factors like diseases including ND, accidents, mortality at young age, parasitic infestations, predations, poor reproductive performance, and poor growth rates (Mapiye and Sibada, 2005). In addition, extensive management system allows for free movements and interactions of chickens with potential disease reservoirs (Marwa et al., 2018). ND together with the other challenges may have contributed to low productivity in Tanzanian FRLCs. Although Tanzanian FRLCs have not been extensively studied for improved productivity, they are believed to have high genetic potentials that could be exploited through selective breeding to improve their productivity (Mpenda et al., 2019). The demand for chicken and chicken products in Tanzania as sources of animal protein is increasing (TMLF, 2017). Thus, control of ND in Tanzanian FRLC flocks will improve their survivability and productivity, and contribute to availability of animal protein in human diet as well as improved family income. The control of ND in the Tanzanian FRLC flocks has been and is still a big challenge due to the free-range nature of the husbandry system in practice which exposes them to the risk of ND. Vaccination is the only main reliable approach (Alders et al., 2012) although vaccination alone cannot fully/effectively control ND in FRLCs because of the scavenging system of husbandry. There are also many challenges to vaccination like poor infrastructure in rural areas, lack of cold chains for storage and transport of the vaccines and insufficient knowledge of vaccinators where, compared to commercial farms where vaccination complemented with biosecurity measures has significantly reduced ND incidences in poultry worldwide (Dortmans et al., 2012; Dimitrov et al., 2016), it is challenging to control ND in FRLCs using the similar approaches. Vaccination in combination with biosecurity measures has been important in control of many diseases including ND and infectious bursal disease (IBD) in commercial poultry (Alders et al., 2012) but not practicable in FRLC flocks.
Due to all these challenges, this research focused on evaluating the immune response of Tanzanian FRLCs for selection of chickens with improved resistance to ND to complement vaccination. Studies with some FRLCs have demonstrated natural resistance to some poultry diseases, including ND (Hassan et al., 2004; Minga et al., 2004; Padhi, 2016). Thus, genetic selection and breeding of Tanzanian FRLCs based on disease resistance could offer a complementary approach to vaccination in the control of ND (Okeno et al., 2012).
This study used phenotypic traits, including antibody levels, viral loads and viral clearance rate as some of the key indicators for ND resistance (Pitcovski et al., 2001) which have also been used by other researchers elsewhere. For instance, natural antibody levels have been used as indicators for resistance to avian pathogenic Escherichia coli (APEC) in chickens (Berghof et al., 2019). The viral titer at the time of infection and the level of antibodies against a viral agent has impact on the development of disease (Classe, 2014). Elsewhere, using similar approaches, some FRLCs have demonstrated natural resistance to some poultry diseases, including ND (Hassan et al., 2004; Minga et al., 2004; Padhi, 2016). Molecular and serological techniques such as polymerase chain reactions (PCRs) and enzyme linked immunosorbent assays (ELISAs) and also genotyping technologies and genomics have advanced the study of animal genetics and improved animal production, for instance, selection for resistance against diseases in poultry (Jie and Liu, 2011). Using these techniques, Rowland et al. (2018), Saelao et al. (2018) and Walugembe et al. (2019, 2020) have identified many quantitative trait loci (QTLs) that affect response to ND in chickens.
With that in focus, this study aimed to evaluate the natural resistance of Tanzanian FRLCs to ND using antibody response, viral load and viral clearance rate as indicator traits for resistance to the disease. Three selected Tanzanian FRLC ecotypes challenged with LaSota, a lentogenic strain of NDV used as a vaccine against ND were used in the study. It also aimed at assessing the effect of NDV challenge on growth rate as an important economic parameter in FRLCs.
MATERIALS AND METHODS
Preparation and handling of experimental chickens
The experiments were conducted at Sokoine University of Agriculture (SUA) in Morogoro, Tanzania. Experimental chickens were kept in the animal facilities of the Department of Animal, Aquaculture and Range Sciences (DAARS). Experimental chickens were selected as described by Walugembe et al. (2019). Briefly, a parent stock from three Tanzanian FRLC ecotypes, namely Ching’wekwe, Kuchi, and Morogoro-medium from four regions in four zones across the Tanzania mainland, were used to generate chickens for the experiments. The Kuchi were from Mwanza and Singida regions, Morogoro-medium were from Morogoro and Tanga and the Ching’wekwe were also from Morogoro and Tanga regions representing the Lake, Central, Northern and Coastal zones, respectively. The ecotypes were identified using characteristic features as described by Msoffe et al. (2001, 2005) and Guni and Katule (2013). A total of 389 mature chickens (324 females and 65 males) made up the parent stock. All the chickens were uniquely identified using aluminium numbered tags, vaccinated for ND, dewormed and held in collection stations before they were transported to SUA for experiments. Each rooster was kept separately in a labeled pen with 6 to 10 females of corresponding ecotype to make a family and maintained on commercial corn-based layer feeds with ad-libitum drinking water. The parent stock generated a total of 1,399 chicks composed of 477 Ching’wekwe, 315 Kuchi, and 607 Morogoro-medium for the experiments.
Preparation of progeny generation chickens
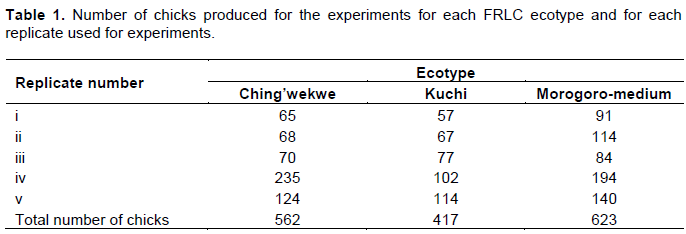
Eggs were collected from the parent flock for periods between 7 and 10 days (less than 10 days old eggs), number-labelled corresponding to the pen numbers of sire to maintain sire identity. For each day, eggs were collected every morning and evening, and subsequently kept at 18°C before being set for incubation. After setting the eggs for incubation, they were candled at day 13 to assess for egg fertility and embryo viability where defective and non-viable eggs were removed. At day 18 of incubation, the eggs were transferred to racks with cubical separations corresponding to sire identity to maintain chick progenies from mixing at hatching. On hatching, day-old chicks were wing-tagged for identity, weighed and transferred to a bio-secured experimental chicken house and maintained on ad-libitum commercial corn-based chick mash and drinking water. The experiment was conducted in five replicates (rounds of laying and hatching) to obtain large number of experimental chickens (Table 1).
Chickens challenging by LaSota NDV Strain
A viral suspension was prepared from the LaSota strain, a commercial vaccine strain of NDV at a titer of 107 of 50% embryos infectious dose (EID50) per bird following the methods described by Ramakrishnan (2016) in specific pathogen free eggs. The stock virus was stored at -80°C before the challenge experiments. The chickens were infected at 28 days of age when the maternal antibodies had waned. A 50 µl of the viral suspension were dropped into each eye and nostril to make a total of 200 µl for each chicken.
Viral load and clearance assays
Tear samples were collected at 2 and 6-days post infection (dpi) into sterile Eppendorf tubes on ice using sterile filtered 200 µl pipette tips through irritation of the ocular mucous membranes with crystalline sodium chloride. The samples were stored in -80°C. Ribonucleic acids (RNAs) were extracted from 50 µl of tear samples using MagMAX-96TM Viral RNA extraction kit (Thermo Fisher Scientific/Life Technologies, USA) that uses the magnetic beads technology. RNA extraction was done in RNAse-free environment decontaminated with RNAse Zap® (Ambion®, USA) and quantified by quantitative real time polymerase chain reaction (qRT-PCRs). The RNA quantification were done using the LSI Vet MAX® NDV qRT-PCR Kit (Thermo Fisher Scientific / Life Technologies, USA) with TaqManTM NDV reagents. The qRT-PCR assays were performed in 7500 fast-real time PCR machine (Applied bio-systems) operated by version 2.3 software. Viral loads were determined using the standard curve method where NDV standards were used to generate a reference standard curve.
Data analyses
Inferential statistics for the chicken populations were determined using the “lsmeans” package in R studio (R Core Team, 2013, R: A language and environment for statistical computing, R Foundation for statistical computing, Vienna, Austria). The quantification of viral loads at the two time points (2 and 6 dpi), antibody titers at 10 dpi, growth rates before challenge and after challenge; data were expressed as LSmean ± SE. The LSmeans for the titers among the chicken groups were analyzed using one-way analysis of variance (ANOVA) and tested for significances of differences using the Tukey honestly significant difference (Tukey HSD) where p-values equal or lesser than 0.05 indicated statistical significant differences among means.
Univariate analyses were performed using linear model to compute the viral loads for the FRLC as least square means (LSmean) with their corresponding standard errors (±SE) considering the chicken ecotypes, the replicate numbers and their interactions. Viral clearance rates were also determined as the difference in viral log copy number from 2 to 6 dpi divided by the viral log copy number at 2 dpi as:

where, VCL is the viral clearance, VL2 is viral loads at 2 dpi, and VL6 is viral loads at 6 dpi.
Antibody response measurement
At 10 dpi, blood samples for the determination of anti-NDV antibody response of the chickens were collected as previously described by Walugembe et al. (2019). Blood samples were aseptically collected from the wing veins into sterile Eppendorf tubes and decanted overnight for sera collection. Enzyme linked immunosorbent assay (ELISAs) were conducted to determine the titers of anti-NDV antibodies (IDEXX NDV ELISA, IDEXX® Laboratories, Inc., Westbrook, ME, United States). Absorbances were read using a spectrophotometer (iMark TM, Micro-plate Reader, USA). Anti-NDV antibody titers were determined from the sample to positive control ratio (S/P) given by the formula:
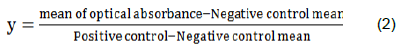
Growth rate before and after NDV challenge with virus
Mean growth rate before challenge and after challenge with LaSota NDV strain were determined. For the growth rate before challenge, body weights were collected from the day of hatch (day 0) to 27 days of age and for growth rate after challenge; the body weights were collected from 28 up to 34 days of age. Individual growth rates (IGRs) were determined according to the formula:
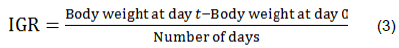
The mean growth rates for the ecotypes were estimated as the least square means (LSmean ± SE) using the LSmeans package in R studio in the following univariate animal model formula.
RESULTS AND DISCUSSION
This study was designed to characterize three Tanzanian FRLC ecotypes in terms of their antibody response, viral load, viral clearance rates and effects on growth rate upon challenge with the LaSota strain of NDV, a non-pathogenic strain of the virus commonly used as a vaccine to protect poultry from ND. Differences observed among the chickens during the challenge experiments in this study may be attributable to mean genetic potentials of the different chicken ecotypes to contain infections through innate protection mechanisms, which according to Chaplin (2010), first recognize the pathogen before the adaptive immune system. Some innate barriers to entry of infectious agents like the NDV and other microbes (Janeway and Medzhitov, 2002) include the physical barriers such as the integrity of the skin, mucus membranes of the mouth, eyes, nose and the mucous itself, anatomical barriers such as the epithelial cell and phagocytic cells enzymes, phagocytic cells, serum proteins related with inflammation such as the complement system proteins and lectins, cells that release cytokines and inflammatory mediators (Aristizába and Gonzalez, 2013), among others. The host immune responses play significant role in determining the severity of disease (Pichyangkul et al., 2003). Differences in abilities of individuals to prevent infectious agents from crossing the barriers could lead to differences in the magnitude of immune response among individuals and populations (Pichyangkul et al., 2003).
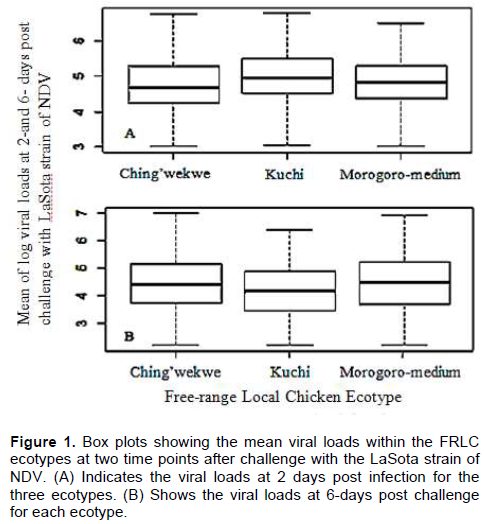
Variations in susceptibility to viral infection in this study could have influenced the observed differences in the viral loads at 2 and 6 dpi and the viral clearance rate by the chickens. In the current study, Kuchi chickens cleared the virus faster than Morogoro-medium and Ching’wekwe, indicating that Kuchi may be more resistant to ND infection compared to the other two. At 2 dpi, the mean log viral load was significantly higher in Kuchi chickens (4.78 ± 0.057) than for Ching’wekwe and Morogoro-medium. Morogoro medium had a mean log of viral load of 4.67± 0.072 while Ching’wekwe had the least mean viral load among the three FRLC ecotypes with mean viral log titer of 4.61 ± 0.077. There was no significant difference in mean log titer between Morogoro-medium and Ching’wekwe (Figure 1A). At 6 dpi, the mean viral loads were significantly lower for Kuchi chickens (4.05 ± 0.088) than for Morogoro-medium (4.21 ± 0.094) and Ching’wekwe (4.32 ± 0.070) but not significantly different between Morogoro-medium and Ching’wekwe (Figure 1B). Kuchi had significantly higher viral clearance rate (12.3%) compared to Morogoro-medium (11.0%) and Ching’wekwe (10.6%) shown in Table 2. However, the viral titers observed in this study in live chickens were high and might be due to the use of less pathogenic strains of NDV and thus might not be seen with more virulent strains. Differences in immune response among local chicken types was demonstrated by Hassan et al. (2004) where using virulent NDV strain and four different Egyptian chicken types, they found that the Mandarah type of chicken had the lowest mortality (20%) compared to other three, in which Gimmizah and Dandarawi types had up to 100% mortality. In a different study using infectious bursal disease virus (IBDv) disease model and the same four Egyptian local chicken types, Hassan et al. (2004) also showed that the Mandarah chickens had the lowest mortality rate (10%) compared to the other three the low viral titers and higher viral clearance observed in Kuchi at 2 dpi compared to Morogoro-medium and Ching’wekwe may indicate that they are less susceptible to the infection better resilient than the others (Table 2). Using a different disease model, Filipovic et al. (2017) also reported differences in susceptibility to infections in different chicken types.
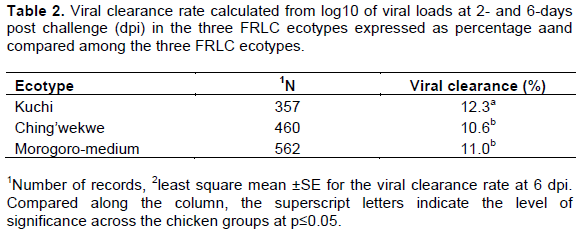
The mean antibody level, one of the immunological responses in the current study, was relatively higher for Kuchi compared to Morogoro-medium, and least for Ching’wekwe ten days after challenging of the chickens with the LaSota strain of NDV. The mean antibody levels were different among the chicken ecotypes where Kuchi had a significantly higher mean anti-NDV antibody level (3.54 ± 0.01) compared to Morogoro-medium (3.50 ± 0.01) and Ching’wekwe (3.39 ± 0.01), respectively (Table 3). The difference was however significant between Morogoro-medium and Ching’wekwe and between Kuchi and Ching’wekwe but not between Kuchi and Morogoro-medium. There were however large variations in the antibody levels among individuals within the ecotypes since the chickens were not pure breeds. The difference in immune response findings observed in this study corroborates findings by Gwakisa et al. (1994) who showed that there were variations in the immune response to NDV infection among Tanzanian ecotypes, however, using different chicken ecotypes from the ones used in the current study. Similarly, Lweramila and Katule (2004) while evaluating the immune response between local and exotic chicken breeds after vaccinating against ND, they found that local chicken types expressed higher levels of anti-NDV antibodies compared to exotic breeds. The variation in antibody response observed in this study could probably indicate that Kuchi and Morogoro-medium ecotype immune systems responded better and more efficiently to infection with NDV by expressing significantly higher level of antibodies against the virus as compared to Ching’wekwe ecotype. Considering the significantly high viral clearance rate in Kuchi compared to Morogoro-medium and Ching’wekwe, Kuchi could be more resistant to ND compared to the other two chicken ecotypes. Neutralizing antibody levels such as for NDV are known to be good indicators of immunity against that infection (Kapczynski et al., 2013). It has also been shown that high levels of antibodies against NDV in commercial layer chickens protect flocks against drop in egg production and deterioration of eggshell quality (shell-less, soft-shell and off-colored eggs) (Bessel et al., 2020; Vanessa and Sembiring, 2020). The high levels of anti-NDV antibodies observed for Kuchi and Morogoro-medium compared to Ching’wekwe might also indicate their higher relative productivity than for the others. The relative differences in immune response to infections among the chicken ecotypes in Tanzania have also been demonstrated using different disease models and chicken ecotypes. The responses of Kuchi immune system from the current study for instance are in support of observations from a similar study but with a different disease model where Msoffe et al. (2006) showed that among five Tanzanian chicken ecotypes they studied, Kuchi ecotype was relatively more resistant to fowl typhoid following infection with virulent strains of Salmonella gallinarum compared to the others. In the current study, a lentogenic strain of NDV was used for the experiments, since use of virulent strains might not have yielded similar results because the chickens would die before sampling and data collection a situation also observed by Msoffe et al. (2005). In a similar study on the immune response against infections in FRLC, Msoffe et al. (2006) found that Morogoro-medium ecotype had higher levels of peripheral leukocytes when infected with S. gallinarum compared to other chicken ecotypes they experimented with. Similar reports have shown variation in infection response where for instance, Okoye and Aba-adulugba (1999) reported that local chickens of Nigeria were more resistant to infectious bursal disease (IBD) compared to exotic chicken types.
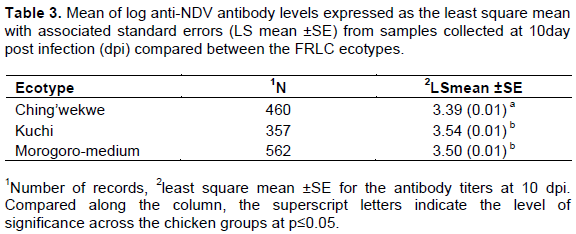
Therefore, this study suggests that Kuchi and Morogoro-medium resist NDV infection better than Ching’wekwe ecotype despite the small differences between the ecotypes. Raising these chickens and complementation with vaccination may reduce loses in flocks and improve their productivity. Additional research may be required to determine the binding affinity of the antibodies to their respective NDV antigens in order to estimate the magnitude of protection. It would also be prudent to perform a similar study with virulent strains of the virus to mimic field conditions. Studies of the immune response to ND in FRLCs have some challenges attributable to the non-commercial nature of the local chickens. The lack of appropriate breeding programs leads to massive diversity among the FRLC ecotypes. This could probably be related to previous findings on researches that did not show significant differences in results among the ecotypes as was shown by Mdegela et al. (2002) while working with Salmonellosis in the FRLCs.
In the current study, growth rate was also evaluated as an important economic trait that could be affected by disease challenges and which is important to FRLCs farmers. It was observed in the study that growth rate was not affected by the viral challenges as indicated by a mean growth rate of 2 g/day before and after challenge with the ND virus. When growth rates were compared among the chicken types, the mean (LSmean ±SE) growth rate before infection (g/day) with NDV was highest for Kuchi (4.30 ± 0.07) followed by Morogoro-medium (4.20 ± 0.06) and least for Ching’wekwe (4.12 ± 0.06). The mean growth rates before the infection were significantly different between the chicken ecotypes. After infection, Kuchi still maintained highest growth rate (6.28 ± 0.21 g/day) compared to Morogoro-medium (6.15 ± 0.17) and Ching’wekwe (6.08 ± 0.18) shown in Table 4. This growth rate was not affected by the infection probably because FRLCs are known to be resistant and resilient to stress (Minga et al., 2004) and also that they were still young and growing, so they were still gaining weight. So, probably because the FRLCs are known to be relatively more resistant and resilient to stress than exotic breeds, challenging them with the avirulent strain of NDV would not affect their growth rate (Minga et al., 2004). This is contrary to the observations by Liu et al. (2015) and Wang et al. (2015) who reported drop in growth rate when broiler chickens were vaccinated with different doses of NDV vaccine and caused immune stress and weight loss compared to observations in the current study where FRLCs were used. The growth rate observed in this study before the challenge was similar to observation by Magonka et al. (2016) who reported a growth rate of 3.96 g/day in Kuchi, whereas in this study, the mean growth of Kuchi was found to be 4.12 g/day before infection. The higher growth rate in the present study might be attributed to the intensive management system in which the chickens were raised which minimizes in free-energy loss during search of feeds and water ranging chickens compared to semi-intensive system used by Magonka et al. (2016). Through genome wide association studies (GWAS) Walugembe et al. (2019) identified five quantitative trait loci (QTL) associated with growth rate and/or immune response to NDV infection in chickens using by single-SNP analyses, with ETS1, TIRAP, and KIRREL3 as related response genes.

CONCLUSION
The current study has revealed that Kuchi chicken ecotype is relatively more innately resistant to Newcastle Disease compared to Ching’wekwe and Morogoro-medium. The high growth rate in Kuchi offers additional economic selection advantage compared to the other ecotypes. Due to the large variations within and between the chicken groups, selection of chickens for breeding with the aid of genomic tools can identify better chicken genotypes within the ecotypes. An extended work is also needed to unravel molecular mechanisms underlying the virus-host interaction in the different FRLC ecotypes that may assist in selecting the right ecotypes and strains of chickens to be raised in the Tanzanian ND endemic areas. It is also important to explore factors other than the ones accessed in the current study to improve productivity of the chickens in the ND endemic stressful environment.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Aristizábal B, González Á (2013). Innate immune system. In: Anaya JM, Shoenfeld Y, RojasVillarraga A, Levy RA, Cervera R, editors. Autoimmunity: from bench to bedside. Bogota DC, El Rosario University Press pp. 31-47. |
|
|
Berghof TVL, Matthijs MGR, Arts JAJ, Bovenhuis H, Dwars RM, van der Poel JJ, Visker MHPW, Parmentier HK (2019). Selective breeding for high natural antibody level increases resistance to avian pathogenic Escherichia coli (APEC) in chickens. Developmental and Comparative Immunology 93:45-57 |
|
|
Bessel PR, Woolley R, Stevenson S, Al-Riyami L, Opondo P, Lai L, Gammon N (2020). An analysis of the impact of Newcastle disease vaccination and husbandry practice on smallholder chicken productivity in Uganda. Preventive Veterinary Medicine 177:104975. |
|
|
Chaplin DD (2010). The immune system. Overview of the immune response. Journal of Allergy and Clinical Immunology 111:S442-59. |
|
|
Classe PJ (2014). Neutralization of Virus Infectivity by Antibodies: Old Problems in New Perspectives. Advances in Biology P 24. |
|
|
Dimitrov KM, Ramey AM, Qiu X, Bahl J, Alfonso CL (2016). Temporal, geographic and host distribution of avian paramyxovirus 1 (Newcastle disease virus). Infection, Genetics and Evolution 39:22-34. |
|
|
Dortmans JC, Peeters BP, Koch G (2012). Newcastle disease virus outbreaks: vaccine mismatch or inadequate application? Veterinary Microbiology 160(1-2):17-22. |
|
|
Ferreira HL, Taylor TL, Dimitrov KM, Sabra Mahmoud, Alfonso CL, Suarez DL, (2019). Virulent Newcastle disease viruses from chicken origin are more pathogenic and transmissible to chickens than viruses normally maintained in wild birds. Veterinary Microbiology 235:25-34. |
|
|
Filipovic A, Ghasemian E, Inic-Kanada A, Lukic I, Stein E, Marinkovic E, Djokic R, Kosanovic D, Schuerer N, Chalabi H, Belij-Rammerstorfer S, Stojanovic M, Barisani-Asenbauer T (2017). The effect of infectious dose on humoral and cellular immune responses in Chlamydophila caviae primary ocular infection. PloS ONE 12(7)e0180551. |
|
|
Guni F, Katule AM (2013). Characterization of local chickens in selected districts of the Southern Highlands of Tanzania: I. Quantitative characters. Livestock Research for Rural Development 25:190. |
|
|
Gwakisa PS, Katule AM, Rugaimukamu EA (1994). Divergent immune responses to NDV vaccination of rural chickens in Tanzania. Tanzania Veterinary Journal 14:198-203. |
|
|
Hassan MK, Afify MA, Aly MM (2004). Genetic resistance of Egyptian chickens to infectious bursal disease and Newcastle disease. Tropical Animal Health and Production 36:1-9. |
|
|
Janeway CA, Medzhitov R (2002). Innate immune recognition. Annual Reviews of Immunology 20:197-216. |
|
|
Jie H, Liu Y (2011). Breeding for disease resistance in poultry: Opportunities with challenges. Worlds Poultry Science Journal 67(4):687-696. |
|
|
Kapczynski DR, Alfonso CL, Miller PJ (2013). Immune responses of poultry to Newcastle disease virus. Development and Comparative Immunology 41(3):447-453. |
|
|
Liu L, Qin D, Wang X, Feng Y, Yang X, Yao J (2015). Effect of immune stress on growth performance and energy metabolism in broiler chickens. Food and Agricultural Immunology 26(2):194-203. |
|
|
Lweramila J, Katule AM (2004). Genetic determination of immune responses to Newcastle disease virus vaccines in chickens. Bulletin of animal Health and Production in Africa 52(3):186-197. |
|
|
Magonka JM, Sendalo DS, Goromela EH, Malingila PB, Daniel E (2016). Production Performance of Indigenous Chicken under Intensive Management Conditions in Central Tanzania. Huria: Journal of Open University of Tanzania 22(1):73-80. |
|
|
Mapiye C, Sibanda S (2005). Constraints and opportunities of village chicken production systems in the smallholder sector of Rushinga District of Zimbabwe. Livestock Research and Rural Development 17:10. |
|
|
Marwa LJ, Mbaga SH, Mutayoba SK, Lukuyu B, (2018). The productivity and management systems of free range local chickens in rural areas of Babati District, Tanzania. Livestock Research for Rural Development 30:34. |
|
|
Mdegela RH, Msoffe PLM, Waihenya RW, Kasanga JC, Mtambo MMA, Minga UM, Olsen JE (2002). Comparative pathogenesis of experimental infections with Salmonella gallinarum in local and commercial chickens. Tropical Animal Health and Production 34(3):195-204. |
|
|
Miller PJ, Koch G (2013). Newcastle disease. In: Swayne DE, Glisson JR, McDougald LR, Nolan LK, Suarez DL, Nair VL, editors. Diseases of Poultry. 13th edition. Ames, IA:Wiley-Blackwell in partnership with the American Association of Avian Pathologists pp. 89-107; p. 120-130. |
|
|
Minga UM, Msoffe PL, Gwakisa PS (2004). Biodiversity (variation) in disease Resistance and in pathogens within rural chicken populations. In: International Health Network for Family Poultry (INFD). World Poultry Congress 8-13 June 2004, Istanbul, Turkey. |
|
|
Mpenda FN, Schilling MA, Campbell Z, Mngumi EB, Buza J (2019). The genetic diversity of local african chickens: A potential for selection of chickens resistant to viral infections. Journal of Applied Poultry Research 28(1):1-12. |
|
|
Msoffe PL, Minga UM, Olsen JE, Yongolo MG, Juul-madsen HR, Gwakisa PS, Mtambo MM (2001). Phenotypes including Immuno-competence in scavenging local chicken ecotypes in Tanzania. Tropical Animal Health and Production 33(4):341-54. |
|
|
Msoffe PLM, Minga UM, Mtambo MMA, Gwakisa PS, Olsen JE, (2006). Differences in resistance to Salmonella enterica serovar Gallinarum infection among indigenous local chicken ecotypes in Tanzania. Avian Pathology 35(4):270-276. |
|
|
Msoffe PLM, Mtambo MMA, Minga UM, Juul-Madsen HR Gwakisa PS (2005). Genetic structure among the local chicken ecotypes of Tanzania based on microsatellite DNA typing. African Journal of Biotechnology 4(8):768-771 |
|
|
Okeno TO, Kahl AK, Peters JK (2012). Characterization of Indigenous Chicken Production Systems in Kenya. Tropical Animal Health and Production 44(3):601-608. |
|
|
Okoye JOA, Aba-adulugba EP (1999). Comparative study of the resistance or susceptibility of local Nigerian and exotic chickens to infectious bursal disease. Avian Pathology 27(2):168-173. |
|
|
Padhi MK (2016). Importance of Indigenous Breeds of Chickens for Rural Economy and their Improvements for Higher Production Performance. Scientifica (2016):9. |
|
|
Pichyangkul S, Endy TP, Kalayanarooj S, Nisalak A, Yongvanitchit K, Green S, Rothman AL, Ennis FA, Libraty DH (2003). A Blunted Blood Plasmacytoid Dendritic Cell Response to an Acute Systemic Viral Infection Is Associated with Increased Disease Severity. Journal of Immunology 171:5571-5578. |
|
|
Pitcovski J, Cahaner A, Heller ED, Zouri T, Gutter B, Gotfried Y, Leitner G (2001). Immune Response and Resistance to Infectious Bursal Disease Virus of Chicken Lines Selected for High or Low Antibody Response to Escherichia coli. Poultry Science 80(7):879-884. |
|
|
Ramakrishnan MA (2016). Determination of 50% endpoint titer using a simple formula. World Journal of Virology 5(2):85-86. |
|
|
Rowland K, Wolc A, Gallardo RA, Kelly T, Zhou H, Dekkers JCM, Lamont SJ (2018). Genetic analysis of a commercial egg laying line challenged with Newcastle disease virus. Frontier Genetics 9:326. |
|
|
Saelao P, Wang Y, Gallardo RA, Lamont SJ, Dekkers JM, Kelly T, Zhou H (2018). Novel insights into the host immune response of chicken Harderian gland tissue during Newcastle disease virus infection and heat treatment. BMC Veterinary Research 14(1):280. |
|
|
Tanzania Ministry of Livestock and Fisheries (TMLF) (2017). Tanzania Livestock master Plan (2017/2018 - 2021/2022) |
|
|
Vanessa V, Sembiring AC (2020). "Quality Improvement of Chicken Eggs Through the Vaccine Schedule Using Database Program System," 2020 3rd International Conference on Mechanical, Electronics, Computer, and Industrial Technology (MECnIT), Medan, Indonesia, pp. 394-398. |
|
|
Walugembe M, Amuzu-Aweh EN, Botchway PK, Naazie A, Aning G, Wang Y, Saelao P, Kelly T, Gallardo RA, Zhou H, Lamont SJ, Kayang BB, Dekkers JCM (2020). Genetic Basis of Response of Ghanaian Local Chickens to Infection with a Lentogenic Newcastle Disease Virus. Frontiers in Genetics 11:739. |
|
|
Walugembe M, Mushi JR, Amuzu-Aweh EN, Chiwanga GH, Msoffe PL, Wang Y, Saelao P, Kelly T, Gallardo RA, Zhou H, Lamont SJ, Muhairwa AP, Dekkers JCM (2019). Genetic Analyses of Tanzanian Local Chicken Ecotypes Challenged with Newcastle Disease Virus. Genes 10(7):546. |
|
|
Wang X, Zhou Q, Shen J, Yao J, Yang X (2015). Effect of difference doses of Newcastle disease vaccine immunization on growth performance, plasma variables and immune response of broilers. Journal Animal of Science and Biotechnology 6(1):20. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


