Full Length Research Paper

ABSTRACT
The present study was conducted to determine the molecular characterization of nucleoprotein (N) gene segment of the rabies virus (RV) in Sultanate of Oman. A portion of nucleotide sequence of N gene was amplified from the brains of 4 rabid animals (two goats, one sheep, and one camel) collected during 2011 to 2012, through reverse transcription polymerase chain reaction (RT-PCR). The nucleotide sequences of the gene were subjected to molecular and phylogenic analysis. The phylogenic analysis was performed in relation to Omani isolates and other sequences from the neighboring countries available in the GenBank. Results indicated that one genetic group of virus appeared to be circulating in the Sultanate of Oman with 99% of homology. The study also indicated that the Omani rabies virus is closely related (> 97% homology) to viruses isolated from Saudi Arabia, Jordan and Israel. It is clear from the results that since its first diagnosis in 1990 the rabies virus has not gone through significant variations from the one isolated from first outbreaks. Since, there is no evidence of any other rabies virus variant in Oman, it seem that rabies is maintained endemically in Oman through wild foxes.
Key words: Rabies, Oman, molecular characterization, phylogenetic analysis, Oman.
INTRODUCTION
Rabies is an infectious viral disease of the central nervous system characterized by severe encephalo-myelitis, change in behavior, paralysis and death. It is endemic in almost all parts of the world, with the excep-tion of few countries including New Zealand Australia, Hawaii, the United Kingdom and Japan (Seimenis, 2008).
Globally, rabies is categorized as either urban (where dogs and cats are the major reservoir hosts), and sylvatic (major reservoirs are foxes, wolves, bats and other wildlife) rabies (World Health Organization, 2013). Based upon genetic similarity, there are 7 recognized genotypes of the virus circulating in the world and genotype 1 is res-ponsible for the classical disease throughout world (David et al., 2007). The principal route of rabies transmission is through animal bites however, other routes such as aerosol (Beran and Steele, 1994) ingestion and trans-plantation have also been documented (Kaplan et al., 1986).
Rabies was first recorded in the Sultanate of Oman in August 1990, when a school boy was bitten by a fox (Anonymous, 1991; Novelli and Malankar, 1991). Within a year, rabies spread was recorded among red foxes (Vulpes vulpes) countrywide and human cases occurred in 1991 (wolf bite), 1992 (fox bite) and 1997 (fox bite) (Anonymous, 1998). Unlike other developing countries, dogs are not central to rabies transmission in Oman; foxes are considered the main reservoir (sylvatic transmission) of rabies (Novelli and Malankar, 1991; Ata et al., 1993; Anonymous, 1998; Scrimgeour and Mehta, 2001; Hussain et al., 2013). All the recorded human cases (n = 8) of rabies in Oman have the history of animal bites (mostly fox bite) and the last human case was through a fox bite recorded in 2003 (Anonymous, 2004). The record of Animal Health Research Center (AHRC), responsible for the surveillance and monitoring of rabies in Oman indicated the disease is still prevalent among different species of livestock.
Rabies virus belong to the genus Lyssavirus in the Family Rhabdoviridae and consist of non-segmented negative single-stranded RNA genome that encodes five structural proteins, nucleoprotein (N), phosphoprotein, matrix protein, glycoprotein (G) and RNA-dependent RNA polymerase (Wunnwr et al., 1988; David et al., 2001). Recent advances in technology have contributed to better understanding of molecular epidemiology and geographic relationships of rabies virus isolates (Susetya et al., 2008; Zhang et al., 2009). The nucleoprotein (N) gene has been extensively used for genetic typing and evolutionary studies because of its relatively conserved variation among reservoir-associated variants and geographic lineages (Bourhy et al., 1993; Wiktore tal., 1980). Moreover, the nucleotide sequence of the N gene used extensively as a molecular marker to explain the patterns of the geographic distribution of rabies virus at the regional and global level (David et al., 2000). There is no information available on the relationship of rabies virus variants to each other or to the host species in Oman, as also, limited information is available on link between these variants in Oman and the submissions from neighboring countries (David et al., 2000, 2007). Therefore, determining the genotype of the circulating rabies virus was important to elucidate the dynamic of disease transmission. Keeping in view the afore mentioned scenario, current study was conducted to understand the molecular characterization of rabies in the sultanate of Oman by sequencing the N gene fragment of virus obtained from the samples submitted for the routine diagnosis to AHRC.
MATERIALS AND METHODS
Virus detection
Either carcasses from different rabies suspected animals species or their chilled/frozen heads (n = 257) were submitted to the virology section of AHRC from all over Oman during 2011 and 2012. Rabies was confirmed in 135 (52.5%) of these animals by a commercial direct immunofluorescent test (FAT) using Anti-Rabies Monoclonal Globulin (Fujirebio Diagnostics Inc. 201 Great Valley Parkway Malvern, PA 19355, USA) (Wiktor and Koprowski, 1978; Wiktor et al., 1980).
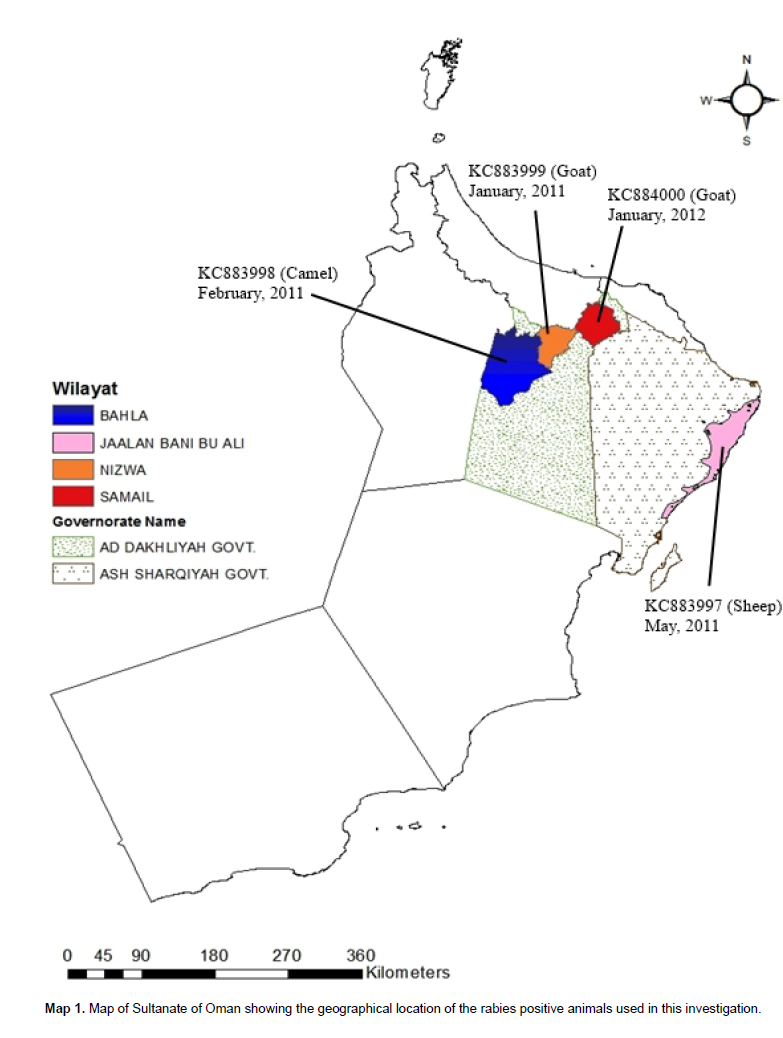
Ten of the FAT positive samples were used for the RT-PCR ana-lysis in this study and 4 samples out of these were further subjected to sequencing. The species of origin location of sequenced samples and the year are shown in Map 1.
Viral RNA extraction and reverse transcription-polymerase chain reaction (RT-PCR)
Total RNA was extracted from 0.2 to 0.5 g of brain tissue using QIA amp Viral RNA kit (QIAGEN, Germany) following the manufacturer’s protocol. The RNA was dissolved in the RNase free buffer provided with kit. PCR was performed using one step RT-PCR kit following the manufacturer’s protocol. For the amplification, the N gene specific primers were designed to target 355 bp conserved regions based upon the previous submissions in the GenBank (Singh et al., 2010). Briefly, 25 ul reaction mix was prepared by using 5 ul of purified RNA template, 5ul of 5× QIAGEN one-step RT-PCR Buffer, 3ul of 5× QIAGEN Q solution, 1.5 of dNTP Mix (10 mM), 3 ul (30 pmol) each of N gene specific forward primer RabN-533F;5-CATTGCAGATAGGATAGAGC-3 and Reverse primer; Rab N-888R;5-GAGGAACGGCGGTCTCCTG-3, 1 ul of QIAGEN one-step RT-PCR enzyme mix, 0.5 ul of RNase inhibitor (20 U/ul), and 3 ul of H2O. The reaction was incubated at 60°C for 1 min, 42°C for 10 min, and 50°C for 30 min for cDNA synthesis (step 1) followed by denaturation at 95°C for 15 min (step 2). After initial denaturation, the amplification was carried out for 30 cycles of three steps, at 95°C for 30 s, 55°C for 30 s, 75°C for 45 s (step 3), with final extension at 72°C for 5 min.
Sequencing of N gene nucleotide
PCR fragments were sequenced using the ABI PRISM BigDye terminator cycle sequencing kit (Perkin Elmer, USA) with primers RabN-533F and RabN-888R, and run on an ABI 310 genetic analyzer. The phylogenetic tree was constructed with neighbor-joining method (Saitou and Nei, 1987) in MEGA (Molecular Evolutionary Genetic Analysis) software (Kumar et al., 2001). Phylogenetic analysis was carried out for 355 nucleotides of each sequence. The sequences of 100 rabies viral isolates submitted from neighboring countries and world were retrieved from GenBank and used for phylogenetic survey. The dendrogram branch lengths represented the predicted number of substitutions. The nucleotide sequences described in this report were submitted to GenBank and assigned the following accession numbers: KC883997, KC883998, KC883999 and KC884000.
RESULTS
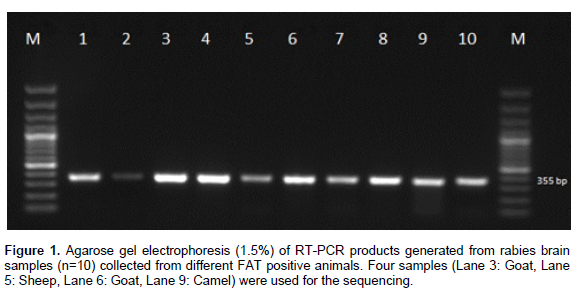
During the study period, 257 suspected samples were submitted to AHRC during 2011 to 2012. Off these, 135 samples were found positive for rabies through FAT. Ten of FAT positive cases were subjected to RT-PCR using N gene specific primer test. A specific product of 355 pb was detected in brain samples of different species on agarose gel electrophoresis (Figure 1).
Similarity and phylogenetic analysis
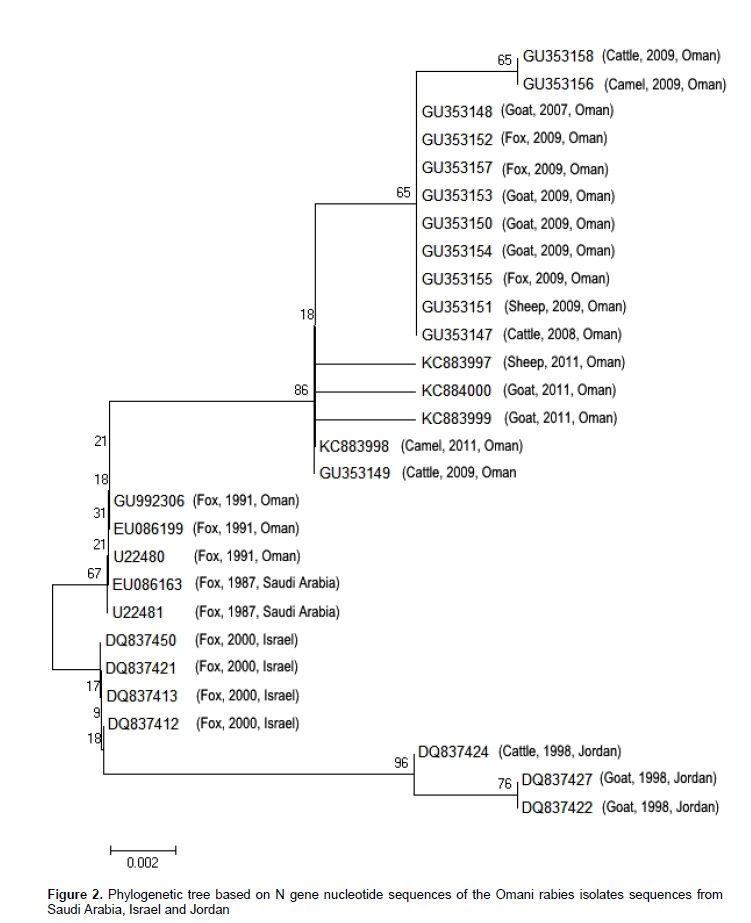
Out of the 10 RT-PCR positive samples, four samples (sheep (1), goat (2) and camel (1) origin submitted from Al Dakhiliyah and Ash Sharqiyah governorates of Oman were sequenced and submitted to GenBank. The details of samples are given in map 1. A total of 355 bp nucleotide sequence encoding the nucleoprotein from 4 FAT and RT-PCR positive samples was determined. The N gene sequences of other rabies isolates obtained from GenBank database were compared with those of 4 Omani isolates to study the phylogenetic relationship. Phylogenetic characterization of the sequenced fragments revealed that the four submissions grouped into one cluster, together with other Omani. The nucleotide similarity among 4 Omani isolates was 99% and with other Omani isolates was found to range from 99- to 98%. Moreover, the Omani isolates collected from foxes during first outbreak (1990s and 1991) were submitted to GenBank in 2000 (EU086166) and 2010 (GU992306) by Pasture Institute also had 98% homology with analyzed samples. Based on these findings red foxes were found to play an important role in the epidemiology of rabies in Sultanate of Oman since 1990. The Omani samples were found genetically indistinguishable from those obtained from foxes from other countries in Middle East and the submissions in this study has 98% homology with the viruses isolated from foxes in Saudi Arabia (U22480 in 1987, foxes from Israel (DQ37450; EU08613) in 2000 and 98 to 97% homology with the viruses isolated during 1998 and 1999 from Jordon (Figure 2), suggesting that only sylvatic rabies virus is circulating in Oman and foxes acted as an important reservoir host in transmitting rabies to domesticated animals and humans in this region.
DISCUSSION
The current study was conducted to characterize Omani rabies virus N gene segments by using molecular and phylogenetic analysis. PCR and sequencing assay des-cribed in this study greatly enhanced our ability to make a sensitive and specific diagnosis as well as typing of animal viruses, including rabies virus, which is an important tool that provides a better understanding of epidemiological relationships (Kissi et al., 1995; Haas, 1997). The nucleotide sequence analysis of the 355 bp fragment of the N gene of isolates represented the first molecular epidemiological study of rabies virus from Oman.
The highly conserved nucleoprotein (N) gene of rabies virus is the most popular target for the diagnosis by using reverse transcription polymerase reaction (PCR) (Ito et al., 1999; Arai et al., 2001). However, it also allows viral strains to be differentiated accurately by analyzing genetic differences present within the gene (Johnson et al., 2002). The molecular sequence and phylogenetic analysis of Omani rabies virus isolates described here, has demonstrated a high degree of similarity between isolates originating from different species which is in agreement with previous finding that rabies virus isolates from different species usually clustered according to geography and not according to the species (Nagarajan et al., 2006; David et al., 2007).
Identification of host population critical to the maintenance of rabies in a region, as opposed to populations in which outbreaks are the result of 'spillover' infections is of uttermost importance in devising the appropriately directed control measures. Our finding further strengthened the claim that rabies virus has established a sustained transmission network in foxes (reservoir host) in Oman (Novelli and Malankar, 1991; Hussain et al., 2013). Foxes were previously indicated as important wildlife reservoir host in Middle East and world (Seimenis, 2008). The close relationship among these isolates and evolutionary clustering of rabies virus isolates from 3 species (sheep, goat and camel) with samples submitted from foxes affected back in 1991 in Oman (GU992306, EU086199 and U22480) and their different geographical regions of collection (Map 1) in this study suggested that virus clustering was not due to geographical origin or host species in Oman.
The phylogenetic analysis indicated that samples originating from Oman were closely related (> 97% homology) to viruses isolated from Saudi Arabia, Jordan and Israel. The isolates from these countries were found closely related in previous studies as well (David et al., 2000, 2007) and well supported by the established paradigm of rabies epidemiology that throughout the world rabies virus had evolved into distinct variants that tend to establish in a region with relatively low species density (Lembo et al., 2007).
CONCLUSION
Our results indicated that based on phylogenetic analysis Omani rabies virus submissions are closely related and the foxes play an important role as a reservoir host of rabies in Oman. Their population and movement could be responsible for the distribution of virus in the country. Application of bait vaccine for wild animals could be used around the place of outbreaks to control the disease in affected areas. The genetic surveillance of circulating rabies virus could become an important tool in monitoring and further understanding the epidemiology of rabies in the Sultanate of Oman.
ACKNOWLEDGEMENTS
The Project was funded by the Directorate General of Agriculture and Livestock Research, Ministry of Agriculture and Fisheries, Oman. We are extremely thankful to the field veterinarians for the collection and submission of the samples.
CONFLICT OF INTEREST
All authors declare that they have no conflict of interest.
REFERENCES
| Anonymous (1991). Wildlife rabies in Oman and the United Arab Emirates. Wkly. Epidemiol. Rec. 67:65-72. | ||||
| Anonymous (1998). Rabies in Oman. Comm. Health Dis. Surv. Newslett. 7(3): 1–9. | ||||
| Anonymous (2004). Human Rabies: Epidemiology & Control. Comm. Health Dis. Surv. Newslett. 13(2):1–9. | ||||
|
Arai YT, Takahashi H, Kameoka Y, Shiino T, Wimalaratne O, Lodmell DL (2001). Characterization of Sri Lanka rabies virus isolates using nucleotide sequence analysis of nucleoprotein gene. Acta Virol. 45: 327-333. Pubmed |
||||
|
Ata FA, Tageldin MH, Al-Sumry HS, Al-Ismaily SI (1993). Rabies in Sultanate of Oman. Vet. Rec. 132:69–69. Crossref |
||||
| Beran GW, Steele JH (1994). Concepts in bacterial, ricketsial, chlamydial and mycotic zoonoses. In: George WB (Ed.), Handbook of Zoonoses, Section A: Bacterial, Rickettsial, Chlamydial, and Mycotic. CRC Press, Boca Raton, Florida, USA. pp. 1-4. | ||||
|
Bourhy H, Kissi B, Tordo N (1993). Molecular diversity of the Lyssavirus genus. Virol. 194:70–81. Crossref |
||||
|
David D, Hughes GJ, Yakobson BA, Davidson I, Un H, Aylan O, Kuzmin IV, Rupprecht CE (2007). Identification of novel canine rabies virus clades in the Middle East and North Africa. J. Gen. Virol. 88:967–980. Crossref |
||||
|
David D, Yakobson B, Smith JS, Stram Y (2000). Molecular epidemiology of rabies virus isolates from Israel and other middle- and Near-Eastern countries. J. Clin. Microbiol. 38:755–762. Pubmed |
||||
| David MK, Peter MH, Diane EG (2001). Fields virology. Lippincott Williams & Wilkins. P 3159. | ||||
|
Haas L (1997). Molecular epidemiology of animal virus diseases. J. Vet. Med. Series B. 44:257–272. Crossref |
||||
|
Hussain MH, Ward MP, Body M, Al-Rawahi A, Wadir AA, Al-Habsi S, Saqib M, Ahmed MS, Almaawali MG (2013). Spatio-temporal pattern of sylvatic rabies in the Sultanate of Oman, 2006-2010. Prev. Vet. Med. 110:281-9. Crossref |
||||
|
Ito N, Sugiyama M, Oraveerakul K, Piyaviriyakul P, Lumlertdacha B, Arai YT, Tamura Y, Mori Y, Minamoto N (1999). Molecular epidemiology of rabies in Thailand.Microbiol.Immunol. 43:551–559. Crossref |
||||
|
Johnson N, McElhinney LM, Smith J, Lowings P, Fooks AR (2002). Phylogenetic comparison of the genus Lyssavirus using distal coding sequences of the glycoprotein and nucleoprotein genes. Arch. Virol. 147:2111–2123. doi:10.1007/s00705-002-0877-4 Crossref |
||||
| Kaplan C, Turner GS, Warrell DA (1986). Rabies: The Facts. Oxford University Press, London, UK. | ||||
|
Kissi B, Tordo N, Bourhy H (1995). Genetic polymorphism in the rabies virus nucleoprotein gene. Virol. 209(2):526–37. Crossref |
||||
|
Kumar S, Tamura K, Jakobsen I B, Nei M (2001). MEGA2: molecular evolutionary genetics analysis software. Bioinformatics 17:1244–1245. Crossref |
||||
|
Lembo T, Haydon DT, Velasco-Villa A, Rupprecht CE, Packer C, Brandão PE, Kuzmin IV, Fooks AR, Barrat J, Cleaveland S (2007). Molecular epidemiology identifies only a single rabies virus variant circulating in complex carnivore communities of the Serengeti. Proc. Biol. Sci. 274:2123–2130. Crossref |
||||
|
Nagarajan T, Mohanasubramanian B, Seshagiri EV, Nagendrakumar SB, Saseendranath MR, Satyanarayana ML, Thiagarajan D, Rangarajan PN, Srinivasan VA (2006). Molecular epidemiology of rabies virus isolates in India. J. Clin. Microbiol. 44:3218–3224. Crossref |
||||
|
Novelli VM, Malankar P (1991). Epizootic of fox rabies in the Sultanate of Oman. R. Soc. Trop. Med. Hyg. 85:543. Crossref |
||||
|
Saitou N, Nei M (1987). The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4: 406–425. Pubmed |
||||
|
Scrimgeour EM, Mehta FR (2001). Rabies in Oman: Failed postexposure vaccination in a lactating woman bitten by a fox. Int. J. Inf. Dis. 5:160-162. Crossref |
||||
| Seimenis A (2008). The rabies situation in the Middle East. Dev. Biol. 131:43–53. | ||||
| Singh KP, Body M, AL-Lamki K, AL-Habsy S, AL-Rawahi Q, AL-Rawahi A, AL-Maawali M (2010). Detection of rabies virus in brain tissue by one-step reverse transcription-polymerase chain reaction. Ind. J. Vet. Pathol. 34:1–4. | ||||
|
Susetya H, Sugiyama M, Inagaki A, Ito N, Mudiarto G, Minamoto N (2008). Molecular epidemiology of rabies in Indonesia. Virus Res. 135: 144–149. Crossref |
||||
|
Wiktor TJ, Flamand A, Koprowski H (1980). Use of monoclonal antibodies in diagnosis of rabies virus infection and differentiation of rabies and rabies-related viruses. J. Virol. Method 1(1):33-46. Crossref |
||||
|
Wiktor TJ, Koprowski H (1978). Monoclonal antibodies against rabies virus produced by somatic cell hybridization: detection of antigenic variants. Proc. Natl. Acad. Sci. 75:3938–3942. Crossref |
||||
|
World Health Organization (2013). WHO Expert Consultation on rabies: Second report. World Health Organization Technical Report Series, 982:1–139. Pubmed |
||||
|
Wunnwr WH, Larson JK, Dietzchold B, Smithm CL (1988). The molecular biology of rabies viruses. Rev. Infect. Dis. 10:771–784. Crossref |
||||
|
Zhang YZ, Xiong CL, Lin XD, Zhou DJ, Jiang RJ, Xiao QY, Xie XY, Yu XX, Tan YJ, Li MH, Ai QS, Zhang LJ, Zou Y, Huang C, Fu ZF (2009). Genetic diversity of Chinese rabies viruses: Evidence for the presence of two distinct clades in China. Infect. Genet. Evol. 9:87–96. Crossref |
||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


