Review

ABSTRACT
This review reports and discusses all available information about the fungal carotenoids such general characters, derivatives, common names, chemical structure, molecular formula, color, bioactivity, and industrial applications in medicine, pharmacology, food processing, cosmetics, dyeing and others. It also reviews the classification of the fungal carotenoids, biosynthetic pathway, distribution and function inside the fungal cells. Approximately, 34 fungal carotenoids derivatives are widely distributed in fungal genera species and fungal groups. Fermented carotenoids produce by fungi from agro-industrial wastes have many advantages and solve the production problems. Development of the fungal carotenoids productivity is reported by two main strategies such as metabolic and genetic engineering.
Key words: Fungal carotenoids, natural pigment, applications, bioactivity.
INTRODUCTION
INDUSTRIAL APPLICATIONS AND ECONOMIC IMPORTANCE OF THE FUNGAL CAROTENOIDS
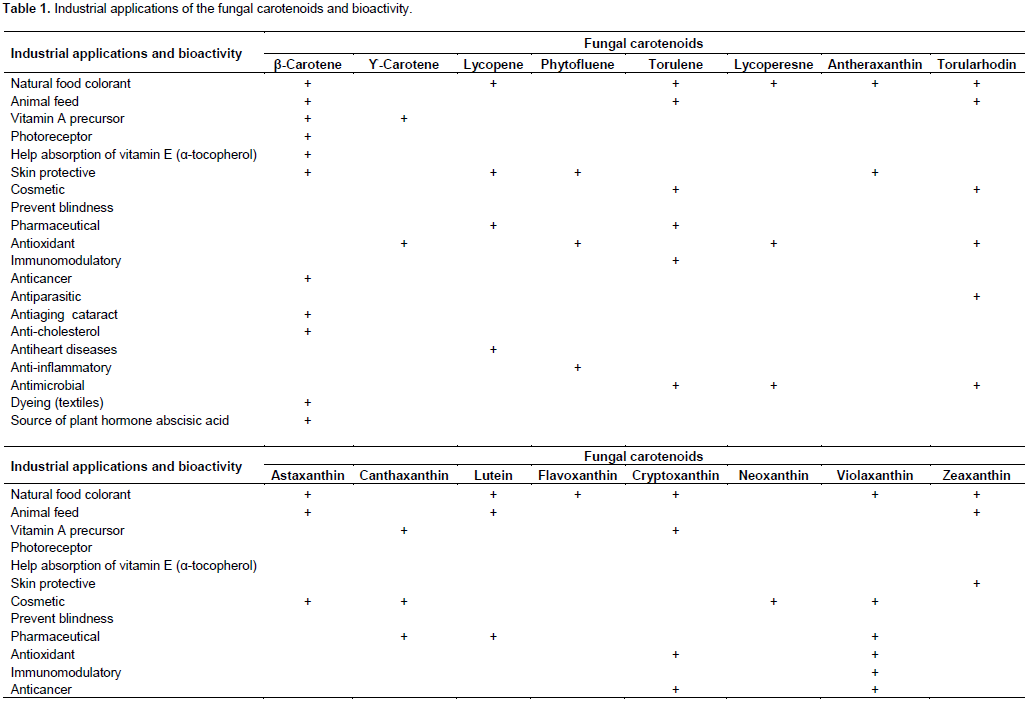
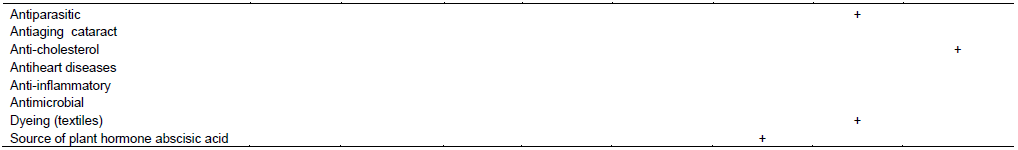
FUNGAL CAROTENOIDS BIOACTIVITY
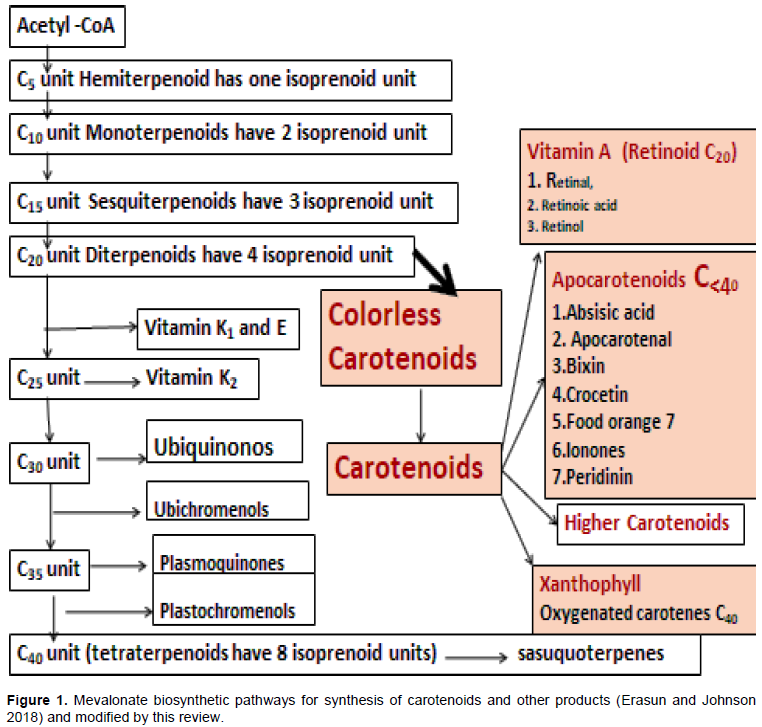
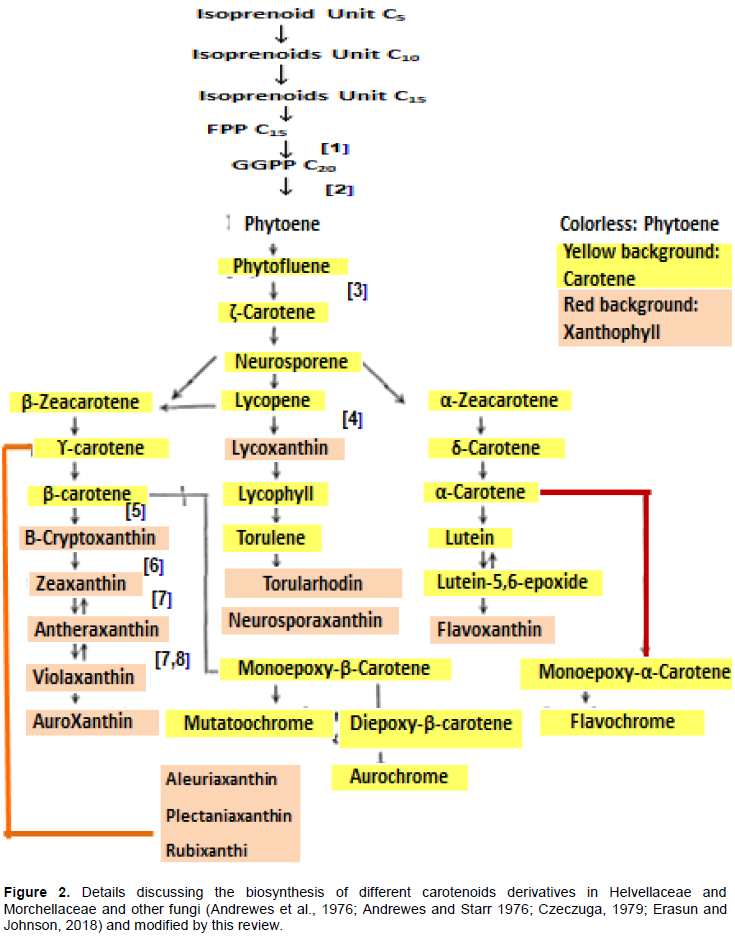
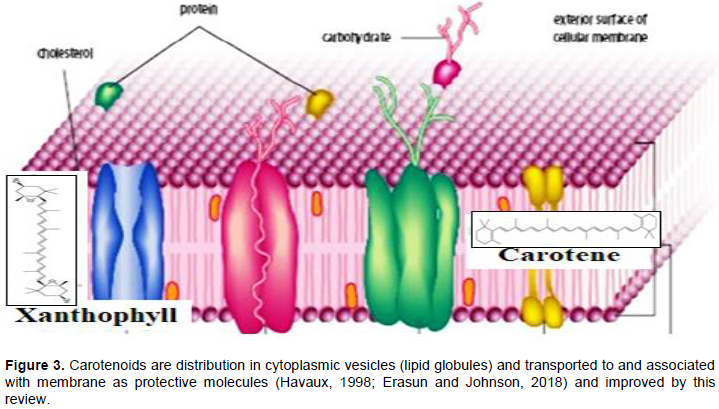
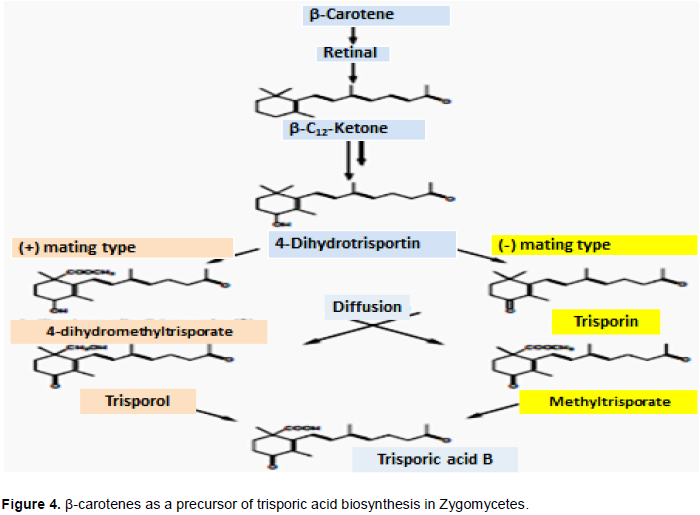
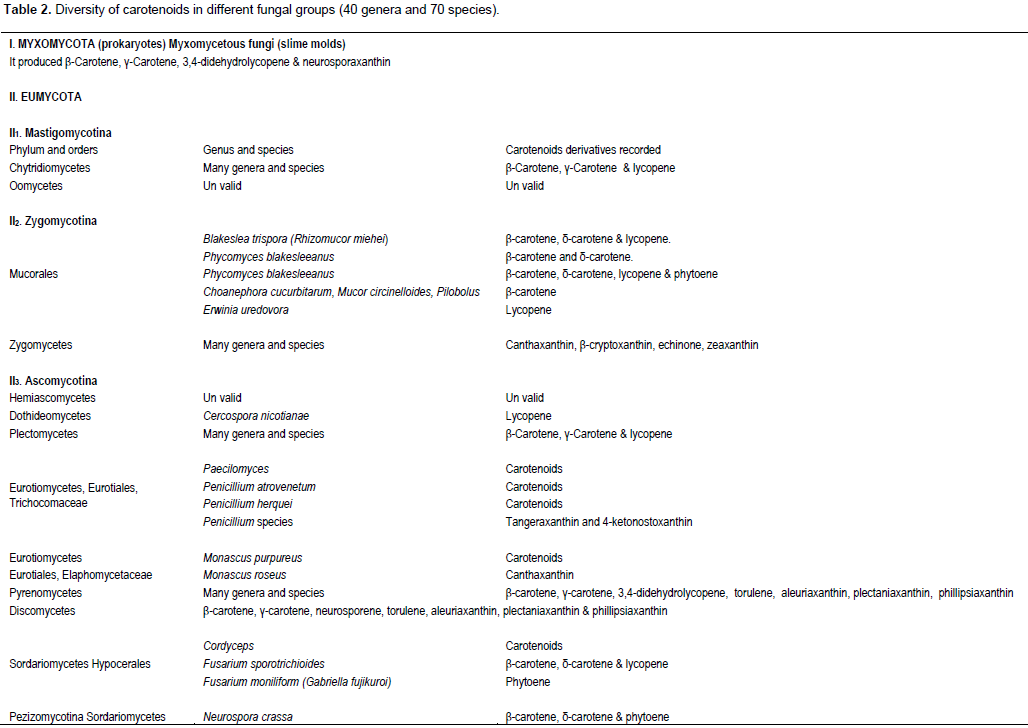
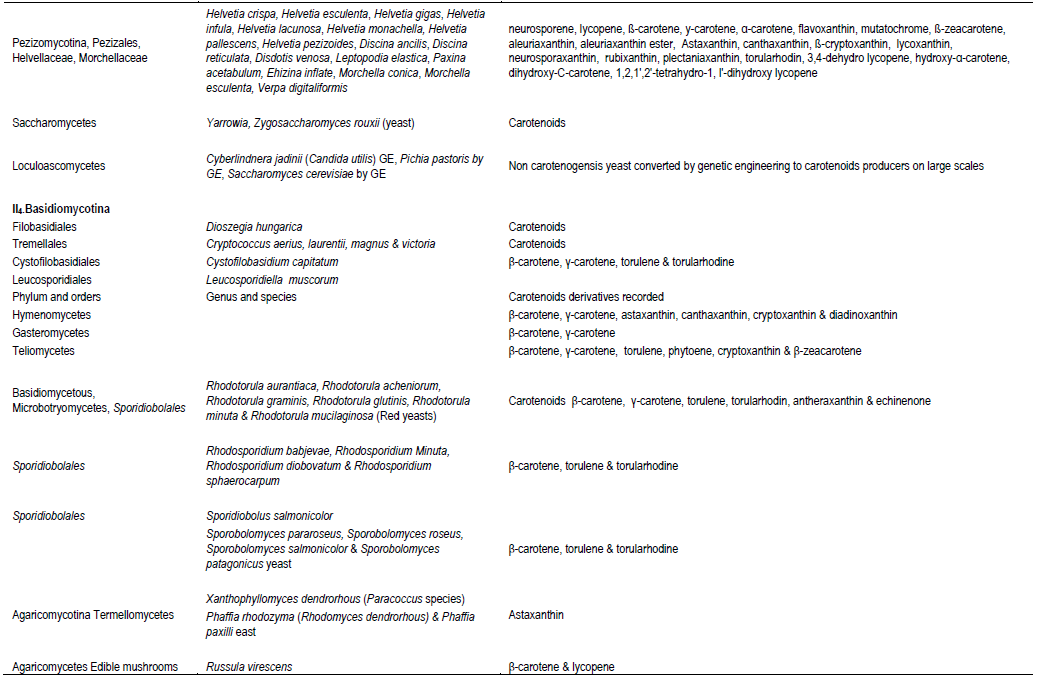

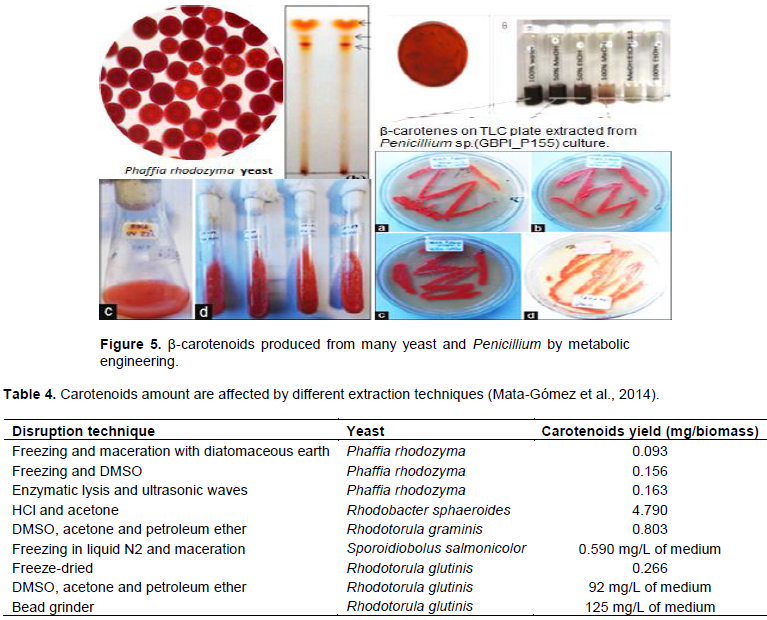
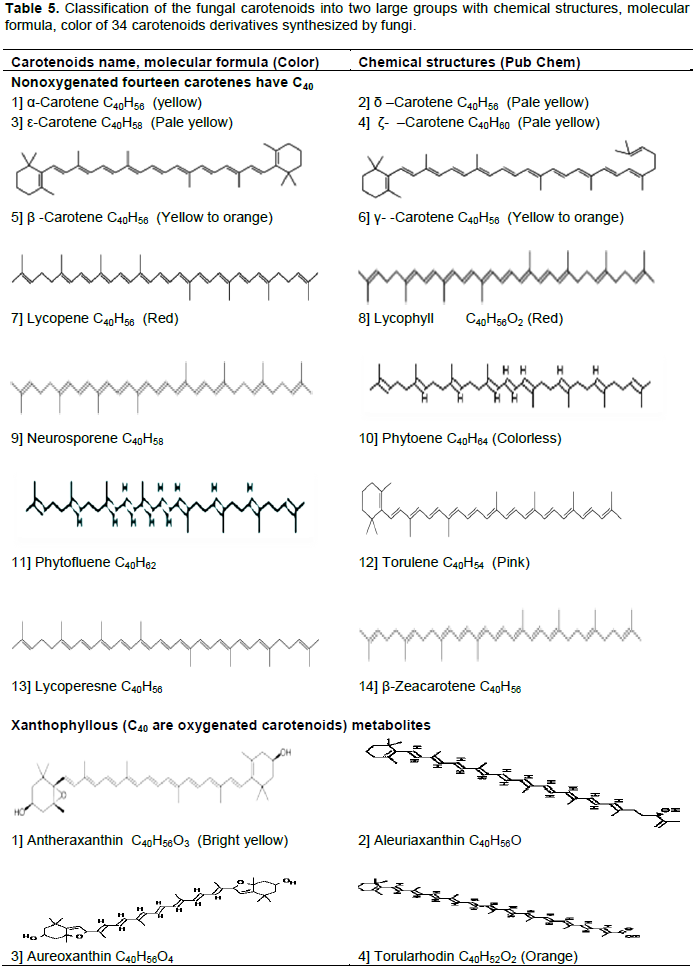
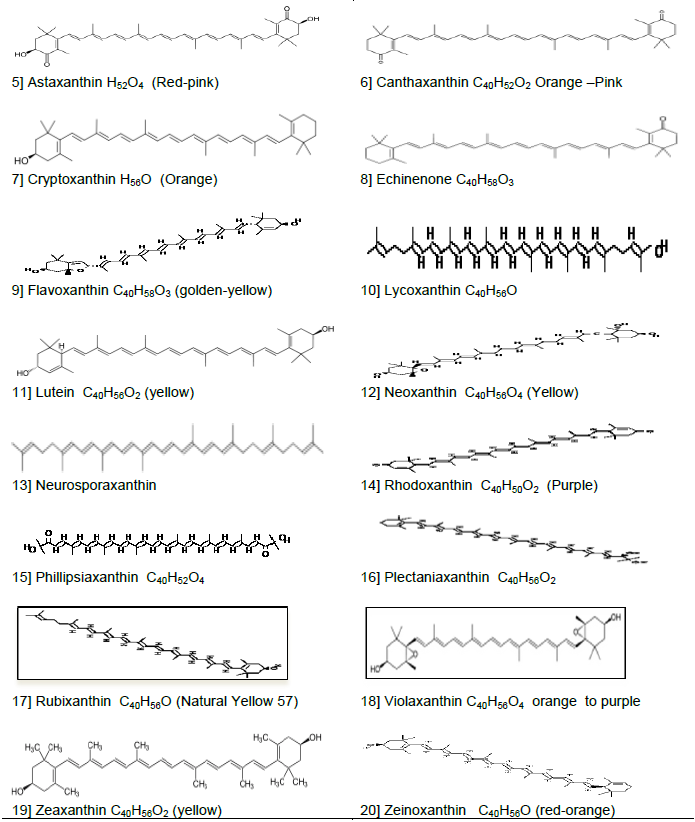
CONCLUSION
CONFLICT OF INTERESTS
The author has not declared any conflict of interests.
REFERENCES
|
Aberoumand A (2011). A Review article on edible pigments properties and sources as natural biocolorants in foodstuff and food industry. World Journal of Dairy and Food Sciences 6(1):71-78. |
|
|
Ahmad WA, Ahmad WY, Zakaria ZA, Yusof NZ (2012). Application of bacterial pigments as colorant. Springer Briefs in Molecular Science, pp. 57-74. |
|
|
Ahmad F, Fanning K, Netzel M, Turner W, Li Y, Schenk PM (2014). Profiling of carotenoids and antioxidant capacity of microalgae from subtropical coastal and brackish waters. Food Chemistry 165:300-306. |
|
|
Andrewes AG, Phaff HJ, Starr MP (1976). Carotenoids of Phaffia rhodozyma, a red pigmented fermenting yeast. Photochemistry15:1003-1007. |
|
|
Andrewes AG, Starr MP (1976). (3R,3'R)-Astaxanthin from the yeast Phaffia rhodozyma. Photochemistry 15:1009-1011. |
|
|
Araya-Garay JM, Ageitos JM, Vallejo JA, Veiga-Crespo P, Sánchez-Pérez A, Tomás G, Villa TG (2012). Construction of a novel Pichia pastoris strain for production of Xanthophylls. Applied Microbiology and Biotechnology 2(24):1-8. |
|
|
Barredo JL (2012). Microbial carotenoids from fungi: Methods and protocols, methods in molecular biology, Springer Science+Business Media New York. |
|
|
Czeczuga B (1979). Investigations on carotenoids in fungi VI. Representatives of the Helvellaceae and Morchellaceae. Phyton (Austria) 19(3-4):225-232. |
|
|
Davies BH (1976). Carotenoids, pp. 38-165. In: Goodwin TW (1976): Chemistry and biochemistry of plant pigments, Academic Press, London, England. |
|
|
Domínguez-Bocanegra AR, Ponce-Noyola T, Torres-Muñoz JA (2007). Astaxanthin production by Phaffia rhodozyma and Haematococcus pluvialis: a comparative study. Applied Microbiology Biotechnology 75:783-791. |
|
|
Eman Mostafa M, Abbady MS (2014). Secondary metabolites and bioactivity of the Monascus pigments. Review Article. Global Journal Biotechnology and Biochemistry 9(1):1-13. |
|
|
Eman Mostafa M, Farghaly FA (2014). Bioactive compounds of fresh and dried Pleurotus ostreatus mushroom. International Journal Biotechnology for Wellness Industries 3:4-14. |
|
|
Eman Mostafa M (2015). Physiological studies on the Monascus ruber red pigment and GC/MS analysis. International Research Journal of Biological Sciences 4(10):1-9. |
|
|
Eman Mostafa M (2016). Flavoring and medicinal values of the yellow pigment produced by Monascus ruber 4066 strain cultivated on static malt agar medium. International Research Journal of Biochemistry and Biotechnology 3(1):37-41. |
|
|
Eman Mostafa M, Khallaf IS, Nassar SM (2018). Antibacterial activities of some yeast strains and GC/MS analysis of Rhodotorula mucilaginosa AUMC13565 bioactive metabolites. Assiut University Journal of Botany and Microbiology 47(2):55-70. |
|
|
Erasun EC, Johnson EA (2018). Fungal carotenoids. Chapter in Applied Mycology and Biotechnology pp1-55. |
|
|
Farghaly FA, Eman Mostafa M (2015). Nutritional value and antioxidants in fruiting bodies of Pleurotus ostreatus mushroom. Journal of Advances in Biology 7(1):1146-1152. |
|
|
Goodwin TW (1976). Chemistry and biochemistry of plant pigments, Academic Press, London, England. |
|
|
Grifin DH (1994). Fungal physiology (2nd Edition). Wiley J & Sons, Inc., New York, ISBN 0471595861. |
|
|
Havaux M (1998). Carotenoids as membrane stabilizers in chloroplasts. Trends Plant Science 3:147-151. |
|
|
Johnson EA, Schroeder W (1995). Microbial carotenoids. Advance Biochemistry Engineering Biotechnology 53:119-178. |
|
|
Joshi VK, Attri D, Bala A, Bhushan (2003). Microbial pigments. Indian Journal Biotechnology 2:362-369. |
|
|
Kiokias S, Proestos C, Varzakas T (2016). A Review of the structure, biosynthesis, absorption of carotenoids analysis and properties of their common natural extracts current research. Nutrition and Food Science 4(1):25-37. |
|
|
Kirti K, Amita S, Priti S, Kumar AM, Jyoti S (2014). Colorful world of microbes: carotenoids and their applications. Review Article. Advances in Biology 13:1-13. |
|
|
Kot AM, Błażejak S, Gientka I, Kieliszek M, Bryś J (2018). Torulene and torularhodin: new fungal carotenoids for industry. Microbial Cell Factories 17(49):3-14. |
|
|
Kuczynska P, Jemiola-Rzeminska M (2017). Isolation and purification of all-trans diadinoxanthin and all-trans diastaxanthin from diatom Phaeodactylum tricornutum. Journal Applied Phycology 29:79-87. |
|
|
Malik K, Tokkas J, Goyal S (2012). Microbial pigments. Review Article. International Journal of Microbial Resource Technology 1(4):361-365. |
|
|
Mata-Gómez LC, Montañez JC, Méndez-Zavala A, Aguilar CN (2014). Biotechnological production of carotenoids by yeasts: An overview Review. Microbial Cell Factories 13(12):1-11. |
|
|
Manimala MR, Murugesan R (2017). Carotenoid pigment production from yeast: Health benefits and their industrial applications. International Journal of Chemical Studies 5(6):392-395. |
|
|
Manik PNR, Xiao M, Li WJ (2017). Fungal and bacterial pigments: Secondary metabolites with wide applications. Chinese Academy of Sciences, Ûrúmqi, China pp. 1-13. |
|
|
Meléndez-Martínez AJ, Stinco CM, Mapelli-Brahm P (2019). Skin carotenoids in public health and nutri-cosmetics: The emerging roles and applications of the UV radiation-absorbing colorless carotenoids phytoene and phytofluene. Review. Nutrients 11:1-41. |
|
|
Miura Y, Kondo K, Saito T, Shimada H, Fraser PD, Misawa N (1998). Production of the carotenoids lycopene, β-carotene and astaxanthin in the food yeast candida utilis. Applied and Environmental Microbiology 64(4):1226-1229. |
|
|
Moharram AM, Eman Mostafa M, Ismail MA (2012). Chemical profile of Monascus ruber strains. Food Technology and Biotechnology 50:490-499. |
|
|
Nagpal N, Munjal N, Chatterjee S (2011). Microbial pigments with health benefits. A mini Review. Trends Bioscience 4:157-160. |
|
|
Narsing-Rao MP, Xiao M, Li WJ (2017). Fungal and bacterial pigments: secondary metabolites with wide applications. Frontiers in Microbiology 8(1113):1-13. |
|
|
Pandey N, Jain R, Pandey A, Tamta S (2018). Optimization and characterization of the orange pigment produced by a cold adapted strain of Penicillium sp. (GBPI_P155) isolated from mountain ecosystem. Mycology 9(2):81-92. |
|
|
Ramesh C, Vinithkumar NV, Kirubagaran R, Venil CK, Dufossé L (2019). Multifaceted applications of microbial pigments: current knowledge, challenges and future directions for public health implications. Review Microorganisms 7(7):186. |
|
|
Rodríguez-Sáiz M, de la Fuente JL, Barredo JL (2010). Xanthophyllomyces dendrorhous for the industrial production of astaxanthin. Applied Microbiology and Biotechnology 88(3):645-658. |
|
|
Sahadevan Y, Richter-Fecken M, Kaerger K, Volgt K, Boland W (2013). Early and late trisporoids differentially regulate β-carotene production and gene transcript levels in the mucoralean fungi Blakeslea trispora and Mucor mucedo. Applied and Environmental Microbiology 29(23):7466-7475. |
|
|
Sardaryan H, Zihlova R, Strnad Z, Cermakoya C (2004). Arpink red- meet a new natural red food colorant of microbial origin. In: Pigments in Food, More than Colours. L. Dufossé (ed) Université de Bretagne Occidentale, Quimper France pp. 207-208. |
|
|
Shimada H, Kondo K, Fraser PD, Yutaka M, Saito T, Misawa N (1998). Increased carotenoid production by the food yeast Candida utilis through metabolic engineering of the isoprenoid pathway. Applied and Environmental Microbiology 64(7):2676-2680. |
|
|
Tan BL, Norhaizan EM (2019). Carotenoids. How effective are they to prevent age-related diseases. Review Molecules 1801:1-23. |
|
|
Tuli HS, Chaudhary P, Beniwal V, Sharma AK (2015). Microbial pigments as natural color sources: current trends and future perspectives. Review Journal Food Science Technology 52(8):4669-4678. |
|
|
Turner WB (1971). Fungal metabolites. Academic Press, London. |
|
|
Wang Q, Liu D, Yang Q, Wang P (2017). Enhancing carotenoid production in Rhodotorula mucilaginosa KC8 by combining mutation and metabolic engineering. Ann Microbiology 67:425-431. |
|
|
Vachali P, Bhosale P, Bernstein PS (2012). Microbial carotenoids. Methods in Molecular Biology 898:41-59. |
|
|
Venil CK, Zakaria ZA, Ahmad WA (2013). Bacterial pigments and their applications. Review. Process Biochemistry 48:1065-1079. |
|
|
Verwaal R, Wang J, Meijnen J, Visser H, Sandmann G, Van Den Berg JA, Albert JJ, van Ooyen AJJ (2007). High-Level production of β-carotene in Saccharomyces cerevisiae by successive transformation with carotenogenic genes from Xanthophyllomyces dendrorhous. Applied and Environmental Microbiology 73(13):4342-4350. |
|
|
Voigt K, Wolf T, Ochsenretter K, Nagy GB, Kaerger K, Sheleste E, Papp TS (2016). Genetic and metabolic aspects of primary and secondary metabolism of the Zygomycetes: Chapter. from book Trehalose metabolism: Enzymatic pathways and physiological functions pp. 361-385. |
|
|
Yamano ST, Ishii M, Nakagawa H, Ikenaga Misawa N (1994). Metabolic engineering for production of β-carotene and lycopene in Saccharomyces cerevisiae. Bioscience Biotechnology Biochemistry 58:1112-1114. |
|
|
Yen H, Zhang Z (2011). Enhancement of cell growth rate by light irradiation in the cultivation of Rhodotorula glutinis. Bioresources Technology 102(19):9279-9281. |
|
|
Yurkov AM, Vustin MM, Tyaglov BV, Maksimova IA, Sineokiy SP (2008). Pigmented Basidiomycetous yeasts are a promising source of carotenoids and ubiquinone Q. Microbiology 77(1):1-6. |
|
|
Zhao Y, Guo L, Xia Y, Zhuang X, Weihua Chu W (2019). Isolation, identification of carotenoid-producing Rhodotorula sp. from marine environment and optimization for carotenoid production Marin Drugs 17(161):1-9. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


