Full Length Research Paper

ABSTRACT
Iron is an essential element, and its deficiency and excess result in negative effects on the growth and physiology of plants. Heavy metals are the highest anthropic pollutants that affect mangrove ecosystem. This study aimed to analyze the development of Rhizophora mangle seedlings in response to excess Fe under controlled conditions. Seedlings of R. mangle previously established were transferred to the culture substrate containing 45 (Control), 135, 225, 270, and 315 µmol L-1 of Fe. The propagules were cultivated for 270 days and during this period the epicotyl length and leaf production were evaluated. At the end of the experiment, the epicotyl length, growth rate, and gas exchange of the seedlings were calculated in the different treatments and the quantification of Fe in the aerial part and root of the seedlings were performed. The excess of Fe led to lower incorporation of biomass, mainly affecting leaf production in the treatment with 315 µmol L-1. A reduction was observed in the assimilation of carbon, also a reduction in stomatal conductance in the highest concentrations of the metal. The excess of Fe in the cultivation solution increased its concentration in the seedlings biomass, with the roots being principally responsible for the bioaccumulation. Although R. mangle plants showed clear signs of stress to excess Fe, they also have a strong capacity for bioaccumulation and tolerance.
Key words: Trace metal, mangrove, toxicity, bioaccumulation.
INTRODUCTION
Pollution by trace metals in mangroves has been the subject of research around the world. Among the toxic chemical compounds, these elements are considered relevant contaminants since they are not biodegradable. Trace metals can be uptake, stored in the biomass, or adsorbed in the roots and transferred to the leaves. As a result, the biomagnification of these metals can occur along the food chain, generating toxicity to the biota with increased concentration, which amplifies the problems for top predators, including humans (Cardwell et al., 2013).
The functionality of the mangrove in retaining these elements in the sediment, due to the system's ability to act as a sedimentary and then biological filter (Thanh-Nho et al., 2019; Tognella et al., 2022), becomes a point of concern since other tensors imposed on the ecosystem can make these metals available, already inert in the sediment or accumulated in the plant biomass.
In November 2015, more than 50 million m³ of mining tailings were released into the Doce River when the Fundão dam ruptured triggering the highest environmental disaster in Brazilian history (IBAMA, 2015; Neto et al., 2016) and one of the major environmental disasters in the mining industry (Carmo et al., 2017). The mud then flowed over the Doce River and reached the estuarine region 17 days after the collapse of the tailings dam. The transport and release of tailings plumes along the coast impacted marine, estuarine and coastal ecosystems on a still unknown scale (Miranda and Marques, 2016). After the collapse of the Fundão dam, the concentration of trace metals at the mouth of the Doce River and on the adjacent continental shelf was monitored, showing an increase in the concentration of Iron (Fe), Aluminum (Al), Manganese (Mn), and Chromium (Cr). For total Fe, the highest concentration (52,360 mg L-1) was recorded in the bottom water samples collected at the station close to the coast. For the dissolved fraction, the maximum concentration was 226 mg L-1 at a station located near the mouth (Neto et al., 2016).
While in excess in the sediment, Fe can interfere with seedling growth, altering the nutrient storage capacity, and delaying the availability of phosphorus from the sediment, in addition to the precipitation of Fe as Iron sulfides, which may enhance the potential for toxicity of these elements (Alongi, 2010) for both plants and biota in the sediment, as well as causing the release of hydrogen protons making the sediment more acidic (Twilley and Rivera-Monroy, 2009).
The effects of heavy metal compounds on mangrove seedlings have been evaluated based on ecological responses such as biomass production (Mejías et al., 2013), leaf loss, bioaccumulation, and translocation (Mejías et al., 2013; Arrivabene et al., 2016; Ray et al., 2021), survival (MacFarlane and Burchett, 2002), functional mechanisms, such as photosynthesis and concentration of photosynthetic pigments (Huang and Wang, 2010), key enzymatic activities in photochemical processes, as well as an increase in the mutation frequency (Caregnato et al., 2008; Zhou et al., 2021) effect on the anatomical structures (Arrivabene et al., 2016; Raju and Ramakrishna, 2021). Despite being a target of interest to the scientific community, there are no studies that evaluate all sides of both the ecological and physiological responses caused by the excess of metals in mangrove species.
Rhizophora mangle L. popularly known as red mangrove, grows in tropical and subtropical regions. Its distribution ranges from West Africa to the Pacific Coast of Tropical America. In America, it has a wide distribution on the side of the Atlantic Ocean with a limit close to 29° N in Florida (Jiménez, 2000) until the south of Brazil at 27° 30´ S (Soares et al., 2015). On the Pacific coast of the continent, it occurs from Mexico to Peru (Tomlinson, 1986; Jimenez et al., 2006), being the dominant mangrove species along the Brazilian coast (Schaeffer-Novelli et al., 1990).
R. mangle forests play an important role in stabilizing the coastline (Riley and Kent, 1999), constituting natural barriers to coastal erosion caused by tropical storms (Vanegas et al., 2019). Because they function as a habitat for a wide variety of organisms, these forests promote connectivity between terrestrial and coastal systems, act as a reservoir, and as carbon sequestration (Schories et al., 2003), among other numerous ecological functions, that are economically defined as ecosystem services (Lovelock and Ellison, 2007). Across the globe, species of this genus provide goods such as wood for the construction of boats and firewood for the livelihood of traditional people, generating many direct social benefits.
As an essential micronutrient for plants, Fe is directly involved in photosynthesis, respiration, nitrogen fixation, hormonal synthesis, and chloroplast maintenance (Hänsch and Mendel, 2009). Despite being fundamental to good plant development, what are the effects of excess Fe on carbon incorporation and assimilation? This study aims to evaluate the effect of Fe on the development and physiology of R. mangles L seedlings under ex-situ cultivation conditions.
MATERIALS AND METHODS
Plant material
Propagules of R. mangle were collected from different adult individuals in the Guaxindiba mangrove, a region located in the vicinity of the natural mouth of the Itaúnas River, Conceição da Barra, Espírito Santo, Brazil, (collect license 75435/1). This mangrove is inserted in the state conservation unit Parque de Itaúnas (4.967-E / 1991). The propagules collected (n = 497) were washed in running water with neutral detergent to remove residues of mud and sand, as well as microorganisms. After washing and screening, only individuals with no visible morphological damage were selected to implement the experiment. The propagules were then immersed in a 2.5% sodium hypochlorite solution for 10 min, followed by a triple rinse in distilled water. After asepsis, the propagules were randomly numbered with plastic seals for individual monitoring during the different stages of this work. After this identification, its length was individually measured using a calibrated measuring tape (Starrett) as was its fresh weight (g) using an analytical balance with an accuracy of 0.001 g (AL500C, Mars). For the installation of the experiment, propagules of similar sizes were selected.
Experimental designs
The experimental design was in randomized blocks (DBC), distributed in five treatments (control and four different Fe concentrations), with seven replicates per concentration, resulting in 35 experimental plots. Each experimental plot contained four propagules totaling 140 propagules. Initially, the propagules were grown for 30 days in a greenhouse on the UFES campus in São Mateus (CEUNES), in plastic containers (pots) with the following dimensions: 49 cm high, 20 cm wide, with a maximum capacity of 20 L. The cultivation technique employed was hydroponics which was used for each 10 L pot of Hoaglad and Arnon's solution (1950) at half strength, with the pH of the solution adjusted to 6.0. Since R. mangle plants are classified as halophytes (Tomlinson, 1986), 14.61 g of NaCl per liter of the solution was added to the solution (Amorim, 2015).
After all the seedlings emitted, the second leaf pair was about 80 days from the beginning of the cultivation. The cultivation solution was removed, and a new nutrient solution containing different Fe concentration in the form of iron chloride (FeCl3) in which 45 µmol L-1 (Control Treatment), 135, 225, 270 and 315 µmol L-1 of Fe were added, the pH of the solution was 5.5 ± 0.2 to improve the solubility of Fe in the substrate.
Fe is an essential micronutrient for plant development and its absence in the solution would cause the seedlings to show signs of deficiency. Thus, the concentration of the element in the control treatment is the same as recommended by Hoagland and Arnon (1950).
Growth analysis
The length of the seedling epicotyl was evaluated every two weeks, individually, with the aid of a digital caliper (King Tools ± 0.01 mm), which is subsequently converted to units of centimeters. The measurements were made from the transition mark between the hypocotyl and epicotyl, going up to the apex of the largest branch, always keeping the initial point fixed.
Length measurements were used to determine the growth rate (GR), which is calculated using the formula:
TC = ΔC / ΔT
Where ΔC corresponds to the change in height (in cm) and ΔT (time in days) refers to the interval between measurements.
In this once every two weeks monitoring, the number of leaves was also determined by manual and individual counting in each seedling, starting from the epicotyl towards the hypocotyl.
Physiological analysis
The acquisition of gas-exchange parameters was performed after 270 days of cultivation. The readings were taken under natural conditions during the morning (from 8 to 10:30 a.m.), on five leaves (second pair, intact and entirely expanded) of five individuals, per treatment.
The net carbon assimilation rate (A μmol m-2s-1), the intercellular CO2 concentration (Ci vpm), and stomatal conductance (gs mol m-2s-1) were obtained using the portable system ADC, model LCi (ADC, BioScientific Ltd. Hoddesdon, England) - IRGA (Infra-Red Gas Analyser). A and Ci were used to estimate the carboxylation efficiency of ribulose-1,5-bisphosphate-carboxylase/oxygenase (Rubisco) (A/Ci) (Zhang et al., 2001).
Iron acummulation
After 270 days of cultivation, the seedlings of each treatment were separated into two compartments, aerial part, and root, and the samples were packed in paper bags identified according to each compartment and their respective treatment. The samples were dried in a forced circulation oven (Fortinox SL 100) at 65°C until constant weight. Then the samples were groundin a Wiley mill (Tecnal brand; model TE650/1) with 20-micron mesh sieves and sent for chemical analysis to determine the Fe concentration. The technique used was atomic absorption spectrophotometry (Thermo Scientific ICE 3000) by the method of Malavolta (1997).
Statistical analysis
The data referring to the parameters: length of hypocotyl, growth rate and number of leaves, were evaluated during the exposure of iron from 90 to 270 days using a two-way ANOVA in Split plot. The control treatment data were submitted to the Dunnet test at 5% probability. The data were evaluated over time by regression. Statistical analyses were performed using software R with packages ExpDes.pt and asbio.
The data referring to iron accumulation were submitted to ANOVA One-Way analysis of variance followed by the Dunnett test with a significance of 5%. The physiological data were subjected to ANOVA variance test with posterior regression analysis. The statistical tests were applied using the software GENES and BioEstat 5.3.
RESULTS
Growth analysis
The propagules of R. mangle from the mangrove at the mouth of the Itaúnas River (n = 497), used for the installation of the experiment, had an average length of 25.10 ± 4.66 cm and an average weight of 16.41 ± 6.49 g. After seven days of cultivation, the emission of roots for all propagules was registered. At the end of the experiment, it was observed that epicotyl length was not affected by Fe (Figure 1A). After the increase of Fe in the solution, the stabilization of epicotyl along the experiment was observed (Figure 1B).
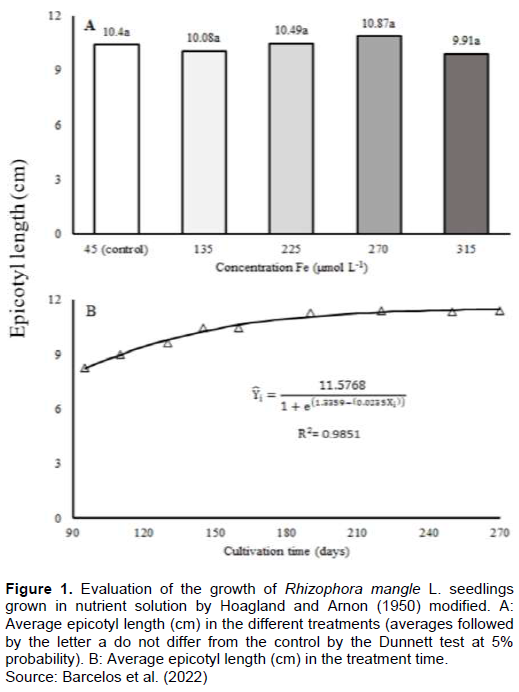
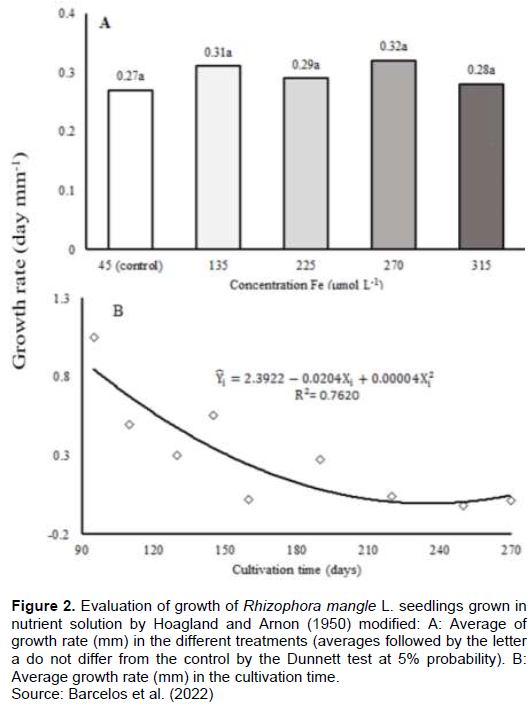
After 190 days of exposure to excess of Fe, there was no significative difference in the final growth rate (Figure 2A). A reduction in the growth rate during the first 90 days after the start of treatments was observed. At 240 days, the growth rate was established (Figure 2B).
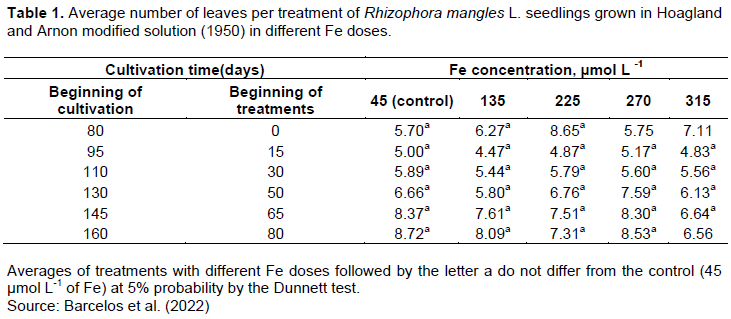
After starting the cultivation under different concentrations of Fe, the seedlings cultivated in the control treatment maintained the number of leaves similar to the cultivation in the initial solution before the Fe increasing; for all other treatments, there was leaf loss throughout the experiment. After 80 days of the beginning of the treatments with Fe, the seedlings of all treatments, except for those cultivated in 315 µmol L-1, maintained the same number of leaves compared to the control (Table 1).
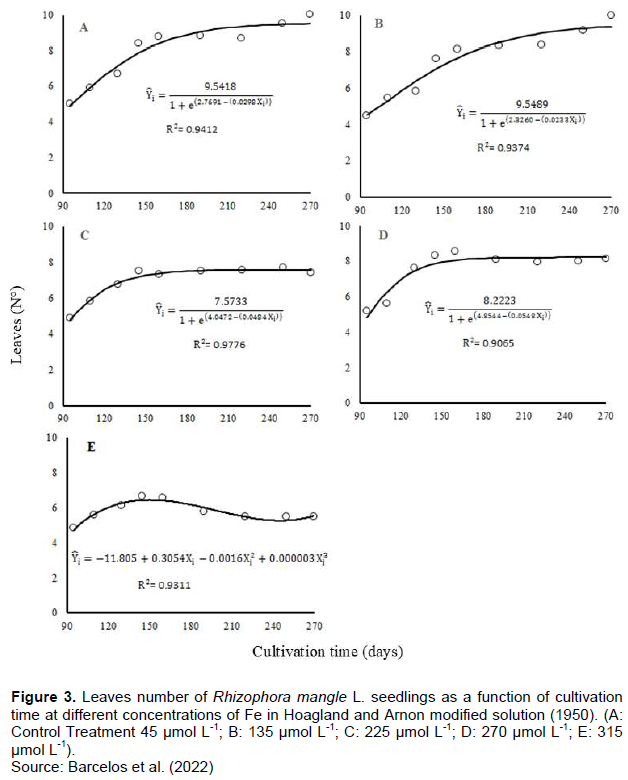
When evaluating the leaf production over time, after the beginning of the treatments, it was observed that the seedlings cultivated in the lowest concentrations (control and 135 μmol L-1 of Fe), maintained the leaf production. Seedlings grown in intermediate concentrations (225 and 270 μmol L-1 Fe) stabilized the number of leaves. For seedlings cultivated in 315 μmol L-1 of Fe, an increase in leaf production followed by loss was observed, for these individuals, and different from seedlings cultivated in intermediate concentrations, no stabilization of leaf production was observed (Figure 3).
Physiological analysis
Leaf CO2 assimilation and gas-exchange variables were analyzed in order to better characterize the effects of Fe on R. mangle photosynthetic performance. The present results identified that the increase in the concentration of Fe in the solution caused different effects on the evaluated parameters (Figure 4A-D). It is possible to observe that above the intermediate concentration of 225 μmol L-1 of Fe, there was an increase in assimilation (A μmol m-2 s-1), however above this concentration, the excess of Fe resulted in a reduction in A (μmol m- 2 s-1). A similar result was observed for the efficiency of carboxylation (A / Ci) and stomatal conductance (gs- mol m-2 s-1). In contrast, the increase in the concentration of Fe did not affect the internal concentration of CO2 (Ci vpm). The maximal concentrations value was determined to estimate the point where the increase of Fe in the solution led to the reduction of the evaluated parameters. The values for A μmol m-2 s-1, A / Ci and gs- mol m-2 s-1 were, respectively 148, 125 and 162, 5 μmol L-1 of Fe.
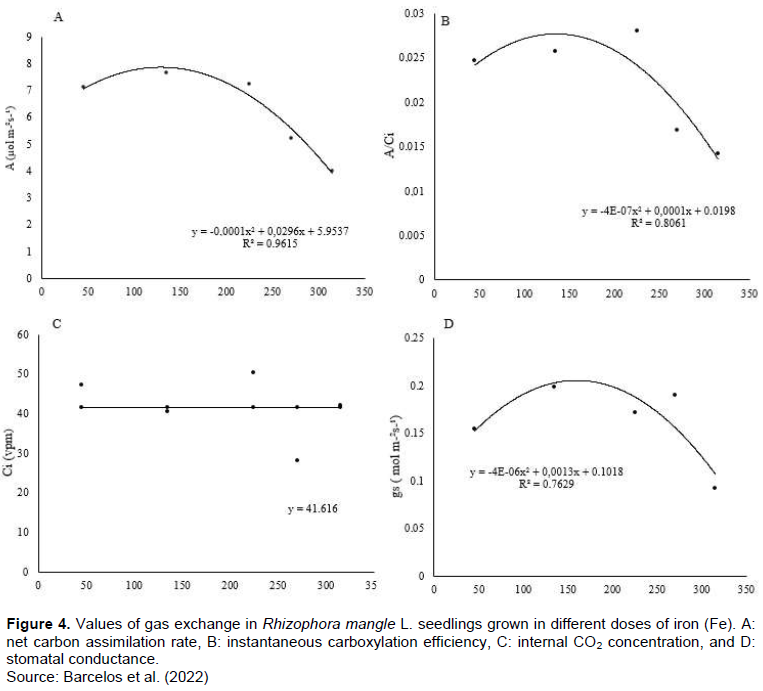
Iron concentration
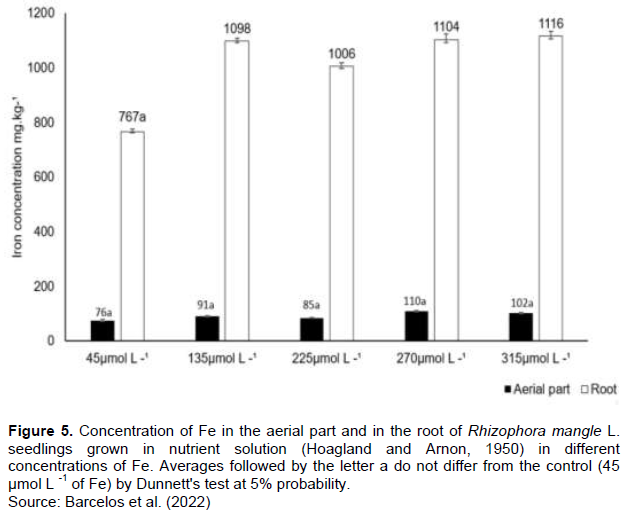
The concentration of Fe in the aerial and root compartments was measured at the end of the experiments, after 270 days of cultivation. A direct relationship was observed between the accumulation of metal in the plant biomass as a result of the increase in the concentration of Fe in the culture solution (Figure 5). For all treatments, the roots had higher Fe bioaccumulation when compared with the concentrations in the aerial parts of the seedling. There was no statistical difference between the control and other treatments considering the aerial part, but a statistical difference was observed for the roots. The highest and the lowest Fe accumulation were observed in the treatment 270 μmol L-1 and in the control (45 μmol L-1), respectively.
DISCUSSION
The fast-initial growth of the seedlings was observed in the first three months. This growth pattern was similar to that found by Ball (2002) in an ex situ experiment and the growth assessment observed by Lima et al. (2018) in situ. The authors also observed a decline and/or stabilization of the growth rate, the same as observed in the present work, which testifies to the expected development for R. mangle seedlings.
Iron is an essential micronutrient for all plants including mangrove species. This metal is directly involved in photosynthesis, respiration, nitrogen fixation, hormonal synthesis, and maintenance of the chloroplast (Hänsch and Mendel, 2009). After 180 days of exposure to Fe, the seedlings showed no significant difference in the growth rate compared to the control. Similar results were observed in seedlings of different Rhizophora species cultivated under other types of stress, as light, nutritional, and salinity (McKee, 1995; Dangremond et al., 2015). The length of the epicotyl also showed no significant difference between treatments. This result differs from Alongi (2010), who evaluated the effect of Fe supply per day in five species of mangrove seedlings. The author observed that the species of the Rhizophoraceae family had the growth of the epicotyl affected only in very high doses of the metal.
Although the presence of Fe did not compromise the initial growth of R. mangle seedlings in high concentrations of this metal (315 µmol L-1), it showed a decrease in the number of leaves after 80 days of treatment. An interesting aspect that demonstrates the adjustment of seedlings to the increased concentration since there was a drop in the leaves immediately after changing the concentrations in the Hoagland solution, only the control continued with progressive leaf growth, as expected for the nutrient conditions of the solution. The over-time concentrations below 315 µmol L-1 recovered the previous number and remained similar to the leaf numbers of conditions.
Under natural conditions, the productivity of mangrove forests is controlled by interactions among regulatory gradients, resource gradients, and hydroperiod (Twilley and Rivera-Monroy, 2009). In the present study, the regulating salinity gradient was kept similar for all treatments, including the control. The luminosity resources and water availability were analogous, only being changed after implantation of Fe with nutrients in different concentrations. The hydroperiod was constant for all treatments, which can be interpreted as similar stress to all experiments.
Dangremond et al. (2015) observed that R. mangle seedlings did not reduce their development with increased salinity. McKee (1995) also observed this pattern with seedlings submitted to luminous and nutritional stress, showing that the species is resilient. Comparing the development of three mangrove species under saline stress, Bompy et al. (2014) observed that R. mangle had no significant reduction in the number of leaves and the production of biomass compared to L. racemosa and A. germinnans. According to the authors, this response is attributable to the energy reserve that the species has in the propagules that makes it manage to invest in the initial development. Thus, our results can be explained through the model proposed by Twilley and Riveira-Monroy (2005), where regulatory gradients, such as salinity, initially increase the acquisition of biomass and then become an energy drain contributing to its decay, whose turning point will depend on the species' strategies concerning salt (Ball, 1988). On the other hand, the stress caused by the gradient of resources (nutrients) has a negative effect on productivity in lower concentrations, gradually increasing with its increase to maintain constant productivity.
Considering that Fe in the mangrove acts as a tensor for the plants, in the same way as salinity, this study shows that R. mangle seedlings may respond in a similar way to metal stress in the environment, as highlighted by the strategy adopted by this species cultivated in excessive concentrations of Fe. As an essential element, the increase in concentrations of Fe initially stimulates the growth and development of the seedling, however, over time, it is possible to observe that in intermediate concentrations there seem to be an adjustment about the tensor and, subsequently, adaptation and development. The results of average leaf production showed similar patterns for all treatments up to the 145th day and reinforce the hypothesis of adjustment for the highest concentrations of Fe. However, the long-term effects of this element accumulation can be observed by the decline in the assimilation of carbon and increasing the concentration of Fe in the roots for the Fe high concentrations treatment (315 µmol L-1).
In contrast to the lower response in terms of seedling growth and development, despite environmental stress, physiological parameters are an essential parameter of observation of the initial tensor data on individuals and collaborate with interpretations that can later explain the decline of a population or community in the face of an acute one-off impact or chronic non-lethal contamination. This analysis makes it possible to evaluate the use of water (transpiration) and gas exchange in plants, indicating processes related to natural tensors (salinity, temperature, and luminosity) or their synergism with other tensors, in this case, the concentration of Fe. Natural tensors are minimized, as the treatments took place under similar conditions, as already discussed. Therefore, they must present low variability among treatments, the differences being explained and interpreted as a result of the loss of net energy from the seedlings since they will adjust in their assimilation processes to compensate for the higher concentrations of Fe (Gurevitch, 2009).
Mangrove species are recognized for their water-conservative behavior despite stress (Ball, 1988; Bompy et al., 2014). The analysis of gas-exchange has been applied mainly in studies related to water and saline stress whose diagnosis of damage is immediate (Lopes et al., 2019).
In this study, the excess of Fe decreased the liquid assimilation rate (A μmol m-2 s-1). This assimilation reduction response was also observed in field evaluations for the same species, comparing situations of different salinities caused by a salt gradient or changes in the precipitation (Lopes et al., 2019). Adamski et al. (2011) obtained similar results, in which the excess of Fe led to a reduction in A μmol m-2 s-1. For the authors, this reduction is associated with a decrease in the reduction capacity of NADH, and the synthesis of ATP.
Seedlings showed a reduction in gs in the highest concentrations of Fe. The decrease of gs is one of the main restrictions to photosynthesis in plants, limiting the influx of CO2 in the leaves (Erickson et al., 2003). However, despite the reduction in these parameters, there was no significant variation in Ci (vpm). According to Alves et al. (2011), a lower A should not only be associated with the lower entry of CO2 in the leaves, but with some biochemical limitation in the fixation of CO2 within the chloroplasts. The increase in A μmol m-2 s-1 and the invariability of Ci were similar to data obtained from seedlings in mangrove areas affected by the tailings plume from the Fundão dam (RRDM, 2020).
The excess of Fe reduced the instant efficiency of carboxylation (A/Ci), which represents the use of Ci for biomass production. Changes in leaf photochemistry and carbon metabolism are often associated with the lower activity of Rubisco and changes in the capacity for regeneration of ribulose-1,5-bisphosphate (Hiscox and Israelstam, 1979). The reduction in Rubisco's activity has also been reported in plants infested with pathogens (D?addazio et al., 2020).
The combination of reduction in A (μmol m-2 s-1) and A / Ci was also observed by Soares et al. (2015) in R. mangle trees that developed in the extreme latitudinal distribution of the species, authors considered to be a physiological restriction caused by environmental temperature.
The concentration of Fe in the plant tissues attended the increase of its availability in the culture solution, where the roots accumulated 75% of the metal when compared to the aerial part. The main source of absorption of trace metals by plants is through the roots, although other tissues may also contribute to this process. A higher concentration of toxic elements in the roots of R. mangle is described in the literature (Mejías et al., 2013; Arrivabene et al., 2016) for mangroves in areas polluted by domestic, agricultural, and industrial effluents. Our results corroborate those of Machado et al. (2005), which report the preferential accumulation of Fe and Zn in the tissues of the roots of R. mangle suppressing the translocation of metals to the leaves. Mangrove roots can immobilize trace metals by forming iron plaque on the root surface and in the rhizosphere (Ye et al., 2001; Thanh-Nho et al., 2019).
Despite the increase in the concentration of iron in the solution, the formation of iron plaque was not observed in this experiment, corroborating the results found by Arrivabene et al. (2016) where seedlings of R. mangle were also cultivated in hydroponical solution with excess of Fe.
R. mangle is considered an exclusive salt species; it prevents the entry of a considerable amount of sodium and chlorine at the root level (Parida and Jha, 2010) through the ultrafiltration process. This is a strategy for plants less tolerant to salinity (Parida and Jha, 2010). Studies assess whether the mechanisms used by species to regulate salt would be the same as those used for trace metals. MacFarlane et al. (2007) evaluated the accumulation and translocation patterns of copper, lead, and metallic zinc cations in the roots and leaves of different mangrove species and observed that in the non-secreting salt species, the translocation of metals to the leaves was lower compared to secretory; the authors then assume that the way of regulating the transport of metals is similar to the way of regulating salt. A similar result was found by Arrivabene et al. (2016), which demonstrates the species' strategy to protect the photosynthetic apparatus from the maximum tensors, preserving the energy-producing structure.
The results of this study indicate that Fe, being an essential element for plants, showed negative effects on carbon assimilation, leaf production, and bioaccumulation in plant organs, corroborating other studies performed with trace elements in mangroves (MacFarlane and Burchett, 2002; MacFarlane et al., 2003; Huang et al., 2010). However, compared to heavy metals, the toxicity is less in the short-term, than long-term, it may compromise mangrove areas with an excess of this element in the sediment. This damage can be intensified in situations of formation of iron sulfides that can alter the pH of the sediment, compromising the biota (Twilley and Riviera-Monroy, 2005).
It was recommended that the knowledge about the role of Fe in other mangrove species should be expanded in higher concentrations and also more assessments should be carried out at the molecular level that will complement the information obtained in this study, supporting the hypotheses raised.
CONCLUSION
Although being an essential element for plants, Fe in high concentrations showed toxic effects on the development of R. mangle seedlings, leading to a reduction in the number of leaves in the highest concentrations of this study. It was observed that the time of exposure to Fe and its concentration are significant factors for the seedlings to begin to morphologically outline the effects. Initially, leaf production was the most affected morphological parameter by the excess of this metal. The integrated analysis of ecological and physiological factors is complementary and could be considered a good indicator for the analyses of Fe toxicity, with the greatest inhibitions observed in the highest concentrations. Seedlings cultivated in the highest concentration (315 µmol L-1) showed a reduction in leaf production and all physiological parameters, except Ci (μmol m-2 s-1). The species showed resilience in dealing with the metal, accumulating most of it in the roots, thus protecting the photosynthetic apparatus.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
This research was developed under the Aquatic Biodiversity Monitoring Program, Environmental Area I, established by the Technical-Scientific Cooperation Agreement nº 30/2018 between Espírito Santense Technology Foundation (FEST) and Renova Foundation, published in Brazil's Official Gazette (Diário Oficial da União). Financial support as scholarship awarded by Coordination for the Improvement of Higher Education Personnel (CAPES) is appreciated.
REFERENCES
|
Adamski JM, Peters JA, Danieloski R, Bacarin MA (2011). Excess iron-induced changes in the photosynthetic characteristics of sweet potato. Journal of Plant Physiology 168(17):2056-2062. |
|
|
Alongi DM (2010). Dissolved iron supply limits early growth of estuarine mangroves. Ecology 91(11):3229-3241. |
|
|
Alves AA, Guimarães LMS, Chaves ARM, DaMatta FM, Alfenas AC (2011). Leaf gas exchange and chlorophyll a fluorescence of Eucalyptus urophylla in response to Puccinia psidii infection. Acta Physiologiae Plantarum 33(5):1831-1839. |
|
|
Amorim KDSP (2015) Ex situ cultivation of Rhizophora mangle L. propagules in different saline concentrations.Master dissertation . Federal University of Espírito Santo. |
|
|
Arrivabene HP, Campos CQ, Costa SI, Wunderlin DA, Milanez CRD, Machado SR (2016). Differential bioaccumulation and translocation patterns in three mangrove plants experimentally exposed to iron. Consequences for environmental sensing. Environmental Pollution 215:302-313. |
|
|
Ball M, Cowan IR, Farquhar, GD (1988). Maintenance of leaf temperature and the optimisation of carbon gain in relation to water loss in a tropical mangrove forest. Functional Plant Biology 15(2):263-276. |
|
|
Ball MC (2002). Interactive effects of salinity and irradiance on growth: Implications for mangrove forest structure along salinity gradients. Trees - Structure and Function 16(2-3):126-139. |
|
|
Bompy F, Lequeue G, Imbert D, Dulormne M (2014). Increasing fluctuations of soil salinity affect seedling growth performances and physiology in three Neotropical mangrove species. Plant and Soil 380(1):399-413. |
|
|
Cardwell RD, Deforest DK, Brix, KV, Adams WJ (2013). Do Cd, Cu, Ni, Pb, and Zn biomagnify in aquatic ecosystems ? Reviews of Environmental Contamination and Toxicology 226:101-122. |
|
|
Caregnato, FF, Koller CE, MacFarlane GR, Moreira JCF (2008). The glutathione antioxidant system as a biomarker suite for the assessment of heavy metal exposure and effect in the grey mangrove, Avicennia marina (Forsk.) Vierh. Marine Pollution Bulletin 56(6):1119-1127. |
|
|
Carmo FF, do Kamino, LHY, Junior RT, Campos IC, de Carmo FF, Silvino G, Castro KJSX, de Mauro ML, Rodrigues NUA, Miranda MPS, Pinto CEF (2017). Fundão tailings dam failures: the environment tragedy of the largest technological disaster of Brazilian mining in global context. Perspectives in Ecology and Conservation 15(3):145-151. |
|
|
Dangremond EM, Feller IC, Sousa WP (2015). Environmental tolerances of rare and common mangroves along light and salinity gradients. Oecologia 179(4):1187-1198. |
|
|
D?addazio V, Silva JVG, Jardim AS, Longue LL, Santos RAA, Fernandes AA, Silva MB, Silva DM, Santos TA, Schmildt ER, Pfenning LH, Falqueto AR (2020). Silicon improves the photosynthetic performance of black pepper plants inoculated with Fusarium solani f. sp. piperis. Photosynthetica 58(3):692-701. |
|
|
Erickson JE, Stanosz GR, Kruger EL (2003). Photosynthetic consequences of Marssonina leaf spot differ between two poplar hybrids. New Phytologist 161(2):577-583. |
|
|
Gurevitch J. Scheiner SM, Fox GA. (2009). Ecologia Vegetal Artmed publish. |
|
|
Hänsch R, Mendel RR (2009). Physiological functions of mineral micronutrients (Cu, Zn, Mn, Fe, Ni, Mo, B, and Cl).Current Opinion in Plant Biology, 12(3):259-266. |
|
|
Hiscox JD, Israelstam GF (1979). A method for the extraction of chlorophyll from leaf tissue without maceration. Canadian Journal of Botany 57 (12):1332-1334. |
|
|
Hoagland DR, Arnon DI (1950). The water culture method for growing plants without soils. Berkeley: California Agricultural Experimental Station. |
|
|
Huang GY, Wang YS (2010). Physiological and biochemical responses in the leaves of two mangrove plant seedlings (Kandelia candel and Bruguiera gymnorrhiza) exposed to multiple heavy metals. Journal of Hazardous Materials 182(1-3):848-854. |
|
|
IBAMA (2015). Brazilian Institute of Environment: Preliminary Technical Report: 313 Environmental Impacts from the Disruption of the Fundão Dam, in Mariana, Minas Gerais. |
|
|
Jiménez JA (2000). Rhizophora mangle L. Mangle colorado. Bioecología de arboles nativos y exóticos de Puerto Rico y las Indias Occidentales: silvics of native and exotic tree species of Puerto Rico and the Caribbean Islands. Puerto Rico: United States Department of Agriculture 450-455. |
|
|
Lima KOO, Tognella MMP, Cunha SR, Andrade HA (2018). Growth models of Rhizophora mangle L. seedlings in tropical southwestern Atlantic. Estuarine, Coastal and Shelf Science 207:154-163. |
|
|
Lopes DMS, Tognella MMP, Falqueto AR, Soares MLG (2019). Salinity variation effects on photosynthetic responses of the mangrove species rhizophora mangle L. Growing in natural habitats. Photosynthetica 57(4):1142-1155. |
|
|
Lovelock CE, Ellison J (2007). Vulnerability of mangroves and tidal wetlands of the Great Barrier Reef to climate change. ISBN 9781876945619. |
|
|
MacFarlane GR, Burchett MD (2002). Toxicity, growth and accumulation relationships of copper lead and zinc in the grey mangrove Avicennia marina (Forsk.) Vierh. Marine Environmental Research 54(1):65-84. |
|
|
MacFarlane GR, Koller CE, Blomberg SP (2007). Accumulation and partitioning of heavy metals in mangroves: A synthesis of field-based studies. Chemosphere 69(9):1454-1464. |
|
|
MacFarlane GR, Pulkownik A, Burchett MD (2003). Accumulation and distribution of heavy metals in the grey mangrove, Avicennia marina (Forsk.)Vierh. Biological indication potential. Environmental Pollution 123(1):139-151. |
|
|
Machado W, Gueiros BB, Lisboa-Filho SD, Lacerda LD (2005). Trace metals in mangrove seedlings: role of iron plaque formation. Wetlands Ecology and Management 13(2):199-206. |
|
|
Malavolta E (1997). Avaliação do estado nutricional das plantas: princípios e aplicações/Eurípedes Malavolta, Godofredo Cesar Vitti, Sebastião Alberto de Oliveira.2. ed. Piracicaba: Potafos. |
|
|
McKee KL (1995). Interspecific Variation in Growth, Biomass Partitioning, and Defensive Characteristics of Neotropical Mangrove Seedlings: Response to Light and Nutrient Availability. American Journal of Botany 82(3):299-307. |
|
|
Mejías LC, Musa CJ, Otero J (2013). Exploratory Evaluation of Retranslocation and Bioconcentration of Heavy Metals in Three Species of Mangrove at Las Cucharillas Marsh, Puerto Rico. Journal of Tropical Life Science 3(1):14-22. |
|
|
Miranda LS, Marques AC (2016). Hidden impacts of the Samarco mining waste dam collapse to Brazilian marine fauna - an example from the staurozoans (Cnidaria). Biota Neotropica 16(2). |
|
|
Neto RR, Sá F, Carneiro MTWD, Costa ES, Rodrigues DRP (2016). The Worst Brazilian Environmental Disaster Altered the Distribution of Metals in Water At the Doce River Estuary and Marine Region. 18th International Conference on Heavy Metals in the Environment, September 15-16. |
|
|
Parida AK, Jha B (2010). Salt tolerance mechanisms in mangroves: A review. Trees - Structure and Function 24(2):199-217. |
|
|
Raju KA, Ramakrishna C (2021). The effects of heavy metals on the anatomical structures of Avicennia marina (Forssk.) Vierh. Brazilian Journal of Botany 44(2):439-447. |
|
|
Riley J, Kent CPS (1999). Riley encased methodology: Principles and processes of mangrove habitat creation and restoration. Mangroves and Salt Marshes 3(4):207-213. |
|
|
Rede Rio Doce Mar (RRDM) (2020). Annual Report of PMBA/Fest-RRDM - Coastal Environment. RT-36C, Aquatic Biodiversity Monitoring Program, Espírito Santo Technology Foundation. 422 page 2020. |
|
|
Schaeffer-Novelli Y, Cintrón-Molero G, Adaime RR, de Camargo TM (1990). Variability of mangrove ecosystems along the Brazilian coast. Estuaries 13(2):204-218. |
|
|
Schories D, Barletta BA, Barletta M, Krumme U, Mehlig U, Rademaker V. (2003). The keystone role of leaf-removing crabs in mangrove forests of North Brazil. Wetlands Ecology and Management 11(4):243-255. |
|
|
Soares MLG, Estrada GCD, Fernandez V, Tognella MMP (2015). Southern limit of the Western South Atlantic mangroves: Assessment of the potential effects of global warming from a biogeographical perspective. Estuarine, Coastal and Shelf Science 101:44-53. |
|
|
Thanh-Nho N, Marchand C, Trady ES, Huu-Phat N, Nhu-Trang TT (2019). Bioaccumulation of some trace elements in tropical mangrove plants and snails (Can Gio, Vietnam). Environmental Pollution 248:635-645. |
|
|
Tognella MMP, Falqueto AR, Espinoza HCF, Gontijo I, Gontijo ABPL, Adriano Alves Fernande, Schmildt ER, Soares MLG, Chaves FO, Schmidt AJ, Lopes DMS, Barcelos UD, D'Addazio V, Lima KOO, Pascoalini SS, Paris JO, Júnior NVB, Porto LA, Filho EA, Oliveira CP, Leopoldo RVS, Leite S, Berribilli MP, Vieiras SFR, da Rosa MB, Sá F, Neto RR, Ghisolfi RD, Castro MSM, Rigo D , Tosta VC, Albino J (2022). Mangroves as traps for environmental damage to metals: The case study of the Fundão Dam, Science of The Total Environment 806(4):150452. |
|
|
Tomlinson PB (1986). The Botany of Mangroves. Cambridge University Presss, Cambridge 1:419. |
|
|
Twilley RR, Rivera-Monroy VH (2005). Developing performance measures of mangrove wetlands using simulation models of hydrology, nutrient biogeochemistry, and community dynamics. Journal of Coastal Research pp. 79-93. |
|
|
Twilley RR, Rivera-Monroy VH (2009). Ecogeomorphic Models of Nutrient Biogeochemistry for Mangrove Wetlands. In Coastal Wetlands: An Integrated Ecosystem Approach 1:641-684. |
|
|
Vanegas G, Catalina A, Osorio AF, Urrego LE(2019). Wave dissipation across a Rhizophora mangrove patch on a Colombian Caribbean Island: An experimental approach. Ecological Engineering 130:271-281. |
|
|
Ye ZH, Cheung KC, Wong MH (2001). Copper uptake in Typha latifolia as affected by iron and manganese plaque on root surface. Canadian Journal of Botany 79:314-320. |
|
|
Zhang S, Li Q, MA K, Chen L (2001). Temperature-dependent gas exchange and stomatal/non-stomatal limitation to CO2 assimilation of Quercus liaotungensis under midday higher irradiance. Photosynthetica 39 (3):383-388. |
|
|
Zhou YY, Wang YS, Inyang AI (2021). Ecophysiological differences between five mangrove seedlings under heavy metal stress. Marine Pollution Bulletin 172:112900. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


