Full Length Research Paper

ABSTRACT
Prostate cancer is the leading cause of mortality in men worldwide, and dietary fat influence its incidence. This study investigated the effect of feeding Tetracarpidium conophorum seed oil (TCSO) on 3-methylcholanthrene (MCA) induced prostate cancer in Wistar rats, the expression of cyclooxygenase-2 (COX-2) and peroxisome proliferator activated receptor gamma (PPAR-?) in the prostatic tissues. The TCSO was extracted with n-hexane in a Soxhlet apparatus and characterized by gas chromatography. Forty-eight male wistar rats (4 weeks old) were divided into three groups of 16 rats each and fed for 12 weeks. Group A and B animals were fed with diet containing TCSO extract. The animals in Groups A and C received intraperitoneally a dose of MCA (150 mg/kg) after 30 days of feeding. Groups A and B rats were fed with diet containing 10% of extracted TCSO throughout the period of the experiment. Results showed that COX-2 activity significantly decreased (p < 0.05) in Group A (0.71±0.07) and B (0.60±0.05) when compared with Group C (1.17±0.10) with increased COX-2 expression. PPAR-gamma activity was significantly increased (p < 0.05) in Group A (1.89±0.13) and B (2.30±0.15) in comparison with Group C (1.16±0.10) which has the lowest PPAR-gamma expression. TCSO extract delayed latency period in Group A where lumps were observed after 4 weeks of 3-methylcholanthrene induction in comparison with Group C where lumps were observed in less than 2 weeks of MCA induction. Gamma-linoleic acid, docosahexaenoic acid and myristoleic acid were higher in the liver cell membrane of animals in Group A compared to animals in Group C. This work therefore showed that TCSO contains bioactive components that may oppose prostate carcinogenesis induced by MCA.
Key words: Cyclooxygenase-2, linoleic acid, prostate cancer, Tetracarpidium conophorum, polyunsaturated fatty acid.
INTRODUCTION
The modulation of cancer by nutritional variables has been a subject of interest and controversy. Dietary fat has received considerable attention as a possible risk factor in the aetiology of prostate cancer (Uhunmwangho and Omoregie, 2017). The second report by the World Cancer Research Fund and the American Institute for Cancer Research indicates that food and nutrition may affect the status of hormones that can modify prostate cancer risk. Both the quantity and quality of dietary fat influence the development of spontaneous as well as chemically-induced neoplasm in laboratory animals. Dietary fat is an essential nutrient and important source for the essential fatty acid (FA), α-linolenic acids, linoleic acids, dihomo-dietary-?-linolenic acid (DHLA), docosahexaenoic acid (DHA), and eicosapentaenoic acid (EPA) which contributes to appropriate growth, development and anti-proliferative diseases, with cancers especially domicile in Tetracarpidium conophorum oil.
African walnut (T. conophorum) is a well-known plant in West Africa. The fruits are edible and the bark, leaves, stem and roots are used in ethno medicinal practice against diseases (Ajibesin et al., 2008). Walnut plant is cultivated principally for the nuts which are cooked and consumed as snacks, along with boiled corn (Adebona et al., 1998). Verheij (2002) have reported on the high nutrient potential of conophor nut. Also, the impacts of traditional processing in the nutrient and sensory qualities of the fruit have been reported (Adebona et al., 1988). A biscuit–like snack food from conophor fruit have been developed throwing some light on the functional significance of the oil seed. African walnut comprises such families as Juglandaceae (English walnut), Euphorbiaceae (African walnut) and Olacaceae (African walnut). Each family has its own peculiar characteristics but they have some things in common such as the nuts. Juglandaceae is mostly found in the Southeast Europe to Japan. T. conophorum (family Euphorbiaceae) is found in Nigeria and Cameroon while Coula edulis (family Olacaceae) which is also referred to as African walnut is found in Congo, Gabon and Liberia (Adebona et al., 1998).
Information on the consumption and composition of T. conophorum oil is far from complete, as the oil becomes more popular and is increasingly commercialized, with such information indispensable for proper valorization of the seed oil. Efforts made so far to optimize the economic and to a lesser extent the nutritional values of the fruit have emphasized their ethno medicinal uses and mineral content, but largely ignored how other components especially the oil content, could be utilized to supplement the nutritional and protective needs of the consumer. This means therefore, there is a need to ascertain the role of T. conophorum oil influences on the carcinogen metabolizing enzymes by which it may exert anti-cancer effects. In this study, we shall investigate the effect of feeding T. conophorum oil on 3-methylcholanthrene in male Wistar rats.
MATERIALS AND METHODS
Study location
This study was conducted in the Department of Biochemistry Laboratory, University of Medical Sciences, Ondo City, Ondo State, Nigeria.
Reagents/Chemicals
All reagents used were of analytical grade. Methanol (Sigma Chemicals Co, London), Chloroform (Sigma Chemicals Co., London), Benzene (BDH Chemicals Ltd., Eng.), NaCl (BDH Chemicals Ltd., Eng.), Standard buffer tablets (BDH Chemicals Ltd., Eng.), Ethanol, 3-methylcholanthrene, Sulphuric Acid Aldrich Chemical Company, USA.
Plant material (Sample collection)
Fresh T. conophorum fruits were obtained from farms in Ondo Town, Ondo State, Nigeria. The fruits were authenticated by a Taxonomist of the Botany Department, University of Medical Sciences, Ondo, Nigeria. At each harvest, 40 fruits will be collected randomly from three regions of the plant as follows, apical region - 10 fruits; middle region – 15 fruits; basal region - 15 fruits. The collected fruits were cleaned with a moist soft cotton wool and then the seeds were carefully separated from the fruits and dried at 65°C for 4 h in an oven, crushed with a laboratory mortar and pestle and were kept in a well labeled air tight polythene bags or screw-capped bottles at 4°C for extraction.
Extraction of oil from African walnut
The Soxhlet extraction method according to AOAC (1996) was employed. The sample (5.0 g) was weighed into a weighed filter paper and folded neatly. This was then placed inside the pre-weighed thimble. The thimble with the sample was inserted into the Soxhlet apparatus and extraction under reflux was carried out with the n-hexane (40-60°C boiling range) for 6 h. At the end of extraction, the thimble was dried in the oven for about 30 min at 100°C to evaporate off the solvent, cool in a desiccator and later weighed and kept in the refrigerator.
Feeding the animals with diet containing walnut seed oil
Male Wistar rats (28 day old) were obtained from the Animal house of the University of Medical Science, Ondo, housed in metal cages in a well-ventilated room, and were allowed access to water ad libitum. The experimental diet comprised of chick pea (51.4%), wheat (15.0%), groundnut cake (10.0%), skim milk powder (6.0%), mineral mixture (2.16%), vitamin mix (0.2%) and T. conophorum oil (15.0%). Overall, 52 Wistar male rats were used, of which 4 will be sacrificed to record zero-day observations. The remaining animals were randomly divided into three major groups of 16 animals each. Group 1 animals were fed for 12 weeks with diet containing T. conophorum oil (10%) and the animals injected with 3-methylcholanthrene (150 mg/kg body weight) through intraperitoneal injection after 4 weeks of feeding. Group 2 were fed for 12 weeks with diet containing T. conophorum oil (10%) only. Group 3 animals were fed for 12 weeks with diet containing no T. conophorum oil, and were given 3-methylcholanthrene (150 mg/kg body weight) through intraperitoneal injection after 4 weeks of feeding. The animals were palpated weekly to determine the time of appearance of tumors and body weight.
At necropsy, the prostatic tissues were exposed and tumors excised. Tumor incidence, volume and weight were determined. Animals from each group were sacrificed at 5, 13 and 21 weeks, and the serum and tissues collected for enzymes and biochemical analysis. Portions of the prostatic tissue from no tumor bearing and tumor tissue were preserved in RNA later for gene expression studies. Another portion of tumor tissue was fixed in formalin (10%) for histopathological studies.
Fatty acid determination
Fatty acids were determined as described by Manni and Caron (1995) and Uhunmwangho and Omoregie (2017).
Cyclooxygenase-2 (COX-2) and PPAR-γ gene expression
The liver samples were placed in triazole (a molecular grid RNA isolating reagent). The samples were homogenized and chloroform added for homogenate gradient separation. This was followed by centrifugation at 15,000 rpm for 15 min. After centrifugation, the upper phase (clear supernatant containing RNA) was aspirated into a new sterile Eppendorf tube of 1.5 ml. The clear supernatant was precipitated by adding isopropanol. This was followed by centrifugation at 15,000 rpm for 5 min. RNA pellet was air-dried for 15 min and resuspended in nuclease free water (30 µL). RNA samples were quantified and absorbance was checked using a spectrophotometer. RNA samples were optimized using PCR machine for 1 h at 42°C. The samples were amplified and gel electrophoresis was carried out at 70 volts, 500 milli amperes for 10 min; thereafter the samples were placed in UV documentary for viewing the expression bands.
Statistical analysis
The values were expressed as mean ± SE. One-way analysis of variance (ANOVA) was used for the feed intake, body weight, tumor weight, tumor volume and COX-2 and PPAR-γ gene expression using Systat 7.0 software (SPSS Inc., Chicago, USA). Statistical analysis of tumor incidence was done by Chi-square test using Systat 7.0 software. A difference with P<0.05 was considered statistically significant.
RESULTS
Table 1 summarizes the data on incidence, latency period, weight and volume of prostate tumors in the prostatic tissues.

The incidence of tumors on T. conophorum seed oil fed group (41.8%) was significantly (P<0.05) lower than animals that were fed with no T. conophorum seed oil but treated with MCA (87.4%). The tumor latency period was 4 weeks in MCA treated group without T. conophorum seed oil compared to 7 weeks in the oil treated group. The average size of tumor was generally larger in MCA administered group than in the animals treated with the seed oil. Similarly, average tumor volume was significantly (P<0.05) less in the seed oil treated groups than on MCA only group.
Major fatty acids composition (%) in prostate gland cells of animals fed with and without Tetracarpidium conophorum seed oil
The major unsaturated FAs were gamma linoleic acid and docosahexaenoic acid as shown in Figure 1.

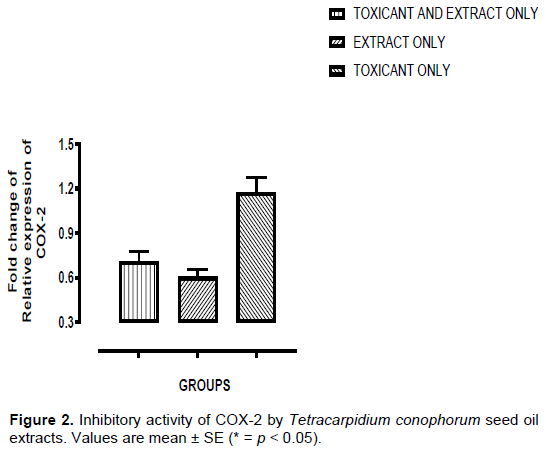
Inhibitory activity of COX-2 by Tetracarpidium conophorum seed oil extracts
Figure 2 showed the effects of T. conophorum seed oil on 3-methylcholanthrene induced prostate carcinogenesis by determining the expression of COX-2 activity. The seed oil extract was found to possess the highest inhibitive activity of COX-2 in rats administered with seed oil extracts only (Group B). There was a significant inhibitory activity of COX-2 in rats that were administered toxicant and oil extract (Group A) when compared to rats that were administered toxicant only (Group C) (p < 0.05). There was no significant inhibitory activity of COX-2 in rats that were administered toxicant and oil extract (Group A) when compared to rats that were given oil extracts only (Group B) (p > 0.05). There was also a significant inhibitory activity of COX-2 in rats given oil extracts only (Group B) in comparison with rats given toxicant only (Group C) (p < 0.05).
Activatory activity of PPAR-gamma by Tetracarpidium conophorum seed oil extracts
The effects of T. conophorum seed oil on 3-methylcholanthrene induced prostate carcinogenesis was evaluated by determining the expression of PPAR-gamma activity. The seed oil extract was found to possess the highest activity of PPAR-gamma in rats that were given oil extracts only (Group B). There was a significant increase in PPAR-gamma activity of rats fed with oil extracts only (Group B) when compared to rats that were administered toxicant only (Group C) (p < 0.05). There was no significant activity of PPAR-gamma in rats that were administered toxicant and oil extract (Group A) when compared to rats that were given oil extracts only (Group B) (p > 0.05). There was also a significant activity of PPAR-gamma in rats given oil extracts only in comparison to rats administered toxicant only (Group C) (p < 0.05).
DISCUSSION
Walnuts are readily available, widely consumed and contain an excellent profile of bioactive components that can exert complex and synergistic effects on carcinogenesis (Sanches et al., 2013). This present study has evaluated the effect of T. conophorum seed oil extract on 3-methylcholanthrene induced prostate carcinogenesis checking for COX-2 and PPAR-gamma protein activity, its anti-carcinogenic activity using a well-established rat cancer model.
COX-2 is a prostaglandin endoperoxidase synthase enzyme (Hung et al., 2004) responsible for generation of prostanoids like prostaglandin E2 that are contributed to the modulation of multiple pro-carcinogenic effects (Li and Zhu, 2015). COX-2 expression is negligible in normal cells (Gurram et al., 2018) in which its basal expression only occurs in the stomach (Su et al., 2016), kidney, central nervous system and in organs of male and female reproduction (Obermoser et al., 2016). COX-2 is a pro-inflammatory enzyme (Pollock et al., 2018) and is overexpressed at the inflammatory site of cancer (Raj et al., 2018). As shown in Figure 2, T. conophorum seed oil extract decrease the expression of COX-2 protein activity in animals treated with the seed oil (0.71±0.07) compared with animals not treated with the seed oil (1.17±0.10) (p < 0.05), and there was no significant difference in the COX-2 protein activity of animals administered with seed oil only and the group of animals treated with seed oil which were earlier administered with MCA. PPAR-gamma is abundant in adipose tissues and appears to be primarily involved in the regulation of lipid metabolism. It also regulates the genes participating in release, transport and storage of fatty acids such as lipoprotein lipase and fatty acid transporter CD36 (Batista et al., 2012). PPAR-gamma also participates in the regulation of cancer development and significantly attenuate tumor progression (Fan et al., 2017). There was an increase (Figure 3) in the activity of PPAR-gamma in animals administered with T. conophorum seed oil only, with result value (1.89±0.13), compared with group of animals that were administered with MCA only with (1.16±0.02). In the group of animals administered with MCA and treated with the T. conophorum seed oil, PPAR-gamma protein activity (2.30±0.15) was also higher than that of animals treated with MCA only, with the result value of (1.16±0.02) (p < 0.05), but there was significant difference between the PPAR-gamma activity of animals administered with MCA and treated with the seed oil and the animals administered MCA only with result values of (2.30±0.15) and C (1.16±0.02) (p > 0.05), respectively. Vanden et al. (2012) reported that PPAR-gamma can be activated by lipid-rich walnut extract and as shown in the result, T. conophorum seed oil extract administered to the rats in resident high amount of gamma-Linolenic acid, docosahexanoic acid and eicosatrienoic acid which are unsaturated fatty acid, hence the increase in PPAR-gamma protein activity.

Research has indicated that T. conophorum oil has anti-cancer activities (Olaniyi et al., 2016; Uhunmwangho and Omoregie, 2017). Anti-carcinogenic activity of T. conophorum is assessed by the antioxidant and anti-inflammatory activities exerted on the 3-methylcholanthrene induced male Wistar rats. Olaniyi et al. (2016) reported that T. conophorum seed oil inhibited inflammation, and Abam et al. (2013) reported that walnut oil acted as an antioxidant in ameliorating the toxic effect of cadmium in the liver, kidney and brain tissues due to the presence of high amounts of bio-flavonoids. Flavonoids intake has been associated with a reduced risk of several chronic diseases with their mechanism of action being attributed to their capacity for anti-oxidation, anti-inflammation, anti-proliferation and modulation of signal transduction pathways. T. conophorum has been suggested to inhibit proliferation of human cancer cell lines via an inhibition of the production of Nitrogen oxide (Casanova et al., 2012). According to Table 1, due to the presence of polyunsaturated fatty acids in T. conophorum seed oil, fewer tumor incidence, smaller tumor size and greater tumor latency period wa observed on T. conophorum seed oil treated group than on the MCA only group, which is suggestive of protection conferred by T. conophorum seed oil in prostate gland carcinogenesis.
Polyunsaturated fatty acids possess diverse bioactivities. Gamma-Linolenic acid is an omega-6 polyunsaturated fatty acid which is associated with anticancer activities both in vitro and in vivo (Uhunmwangho and Omoregie, 2017). Dietary supplement of gamma-linolenic acid reduced tumor growth in an implanted WRC256 rat model (Colquhoun, 2002). More interestingly, gamma-linolenic-induced cytotoxicity has been shown to exhibit high selectivity toward cancer cells with no significant effect on normal cell growth. Series of study also suggested that 3-7 days of incubation with gamma-linolenic acid could selectively induce cell death in various human cancer cell lines including prostate cancer cell PC-3 without affecting normal cell growth (Dan, 2006). Myristoleic acid is a monounsaturated fatty acid that is biosynthesized from myristic acid by the enzyme Stearoyl-CoA desaturase-1. Naoya et al. (2001) reported that myristoleic acid is a cytotoxic component in the extract from Serenoa rapens which induce apoptosis and necrosis in human prostate LNCaP cells.
From Figure 1, percentage of gamma-linolenic acid, eicosadienoic acid, eicosatrienoic acid and docosahexaenoic acid, were high in animals treated with the seed oil in comparison to animals not treated with T. conophorum seed oil. This result showed that the polyunsaturated fatty acids resident in the T. conophorum seed oil has the capacity to exert molecular influence on COX-2 and PPAR-gamma protein expression leading to the prevention of prostate carcinogenesis.
CONCLUSION
T. conophorum seed oil protects against MCA induced prostate carcinogenesis and the effect is mediated through decreased expression of COX-2 and increased expression of PPAR-γ. Further work is needed to understand the apoptotic singling, cell proliferation and prostaglandin synthesis in response to dietary fat.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Abam EO, Oladipo FY, Atasia VN, Obayomi AA (2013). Effect of walnut (Tetracarpidium conophorum) oil on cadmium-induced alterations in lipid metabolism in male albino rats. Food and Public Health 3(4):169-175. |
|
|
Adebona MB, Ogunsuo AO, Ologunde MO (1998). Development of conophor nut based cereal snack food I-biscuits. Journal of Food and Agriculture 2:123-126. |
|
|
Ajibesin KK, Rene N, Bala DN, Essiett UA (2008). Antimicrobial activities of the extracts and fractions of Allanblackia floribunda. Biotechnology 7:129-133. |
|
|
Batista FA, Trivella DB, Bernardes A, Gratieri J, Oliveira PS, Figueira AC (2012). Structural insights into Human Peroxisome Proliferator Activated Receptor Delta (PPAR-DELTA) Selective Ligand Binding. PLoS One 7(5):33643. |
|
|
Casanova F, Quarti J, Costa DC, Ramos CA, Da Silva JL, Fialho E (2012). Resveratrol chemo-sensitizes breast cancer cells to melphalan by cell cycle arrest. Journal of Cellular Biochemistry 113(8):2586-2596. |
|
|
Colquhoun A (2002). Gamma-linolenic acid alters the composition of mitochondrial membrane subfractions, decreases outer mitochondrial membrane binding of hexo-kinase and alters Carnitine Palmitoyltransferase 1 properties in the Walker 256 rat tumor. Biochimica Biophysica 1583(1):74-84. |
|
|
Dan UN (2006). Tumoricidal and antiangiogenic actions of gamma-linolenic acid and its derivatives. Current Pharmaceutical Biotechnology 7(6):457-466 |
|
|
Fan W, Waizenegger W, Lin CS, Sorrentino V, He MX, Wall CE, Li H, Liddle C, Yu RT, Atkins AR (2017). PPAR-δpromotes running endurance by preserving glucose. Cell Metabolism 25(5):1186-1193 |
|
|
Gurram B, Zhang S, Li M, Xie Y, Heng X (2018). Celecoxib conjugated fluorescent probe for identification and discrimination of cyclooxygenase-2 enzyme in cancer cells. Analytical Chemistry 90(8):5187-5193. |
|
|
Hung JH, Su IJ, Lei HY, Wamg HC, Lin WC, Chang WT, Lai MD (2004). Endoplasmic reticulum stress stimulates the expression of cyclooxygenase-2 through activation of NF-kB and pp38 mitogen-activated protein kinase. Journal of Biological Chemistry. 279(45):46384-46392. |
|
|
Li F, Zhu YT (2015). Activated colonic fibroblasts mediates carcinogenesis of colonic epithelial cancer cells via PKC-cMET-ERK1/2-COX-2 signalling. Cellular Signalling 27(4):860-866. |
|
|
Manni G, Caron F (1995). Calibration and Determination of Volatile Fatty Acids in Waste Leachates by Gas Chromatography. Journal of Chromatography 690(2):237-242. |
|
|
Naoya K, Tsutomu S, Tamayo U, Mika M, Mariko K, Sayomi H, Kimitsuna W, Tatsuo T (2001). The human mitochondrial ribosomal protein genes: mapping of 54 genes to the chromosomes and implications for human disorders. Genomics 77(1-2):65-70 |
|
|
Obermoser V, Urban ME, Murgueitio MS, Wolber G, Kintscher U, Gust R (2016). New telmisartan-derived PPAR-gamma agonists: impact of the 3D-binding mode on the pharmacological profile. European Journal of Medicinal Chemistry 124:138-152. |
|
|
Olaniyi FE, Bamidele IO, Omokehinde AO, Ayodeji AA (2016). Anti-inflammatory activities of the chloroform extract of the fruit of Tetracarpidium conophorum (Mull.Arg) (Nigerian walnuts). Journal of Advance in Medical and Pharmaceutical Sciences 6:1-7. |
|
|
Pollock JK, Greene LM, Nathwani SM, Kinsella PO, Boyle NM, Meegan MJ, Zisterer DM (2018). Involvement of NF?κB in mediating the anti?tumour effects of combretastatins in T cells. Investigational New Drugs 36(4):1-13. |
|
|
Raj V, Bhadauria AS, Singh AK, Kumar U, Rai A, Keshari AK, Saha S (2018). Novel 1,3,4?thiadiazoles inhibit colorectal cancer via blockade of IL?6/COX?2 mediated JAK2/STAT3 signals as evidenced through data?based mathematical modeling. Cytokine. |
|
|
Sanches LD, Santos SA, Carvalho JR (2013). Protective effect of gamma-tocopherol enriched diet on N-methyl-N-nitrosourea induced epithelial dysplasia in rat ventral prostate. International Journal of Experimental Patholology 94(6):362-372. |
|
|
Su CW, Zhang Y, Zhu YJ (2016). Stromal COX-2 signalling are correlated with colorectal cancer. A Review Critical Reviews in Oncology/Hematology 107:33-38. |
|
|
Uhunmwangho ES, Omoregie ES (2017). Changes in lipid profile and fatty acid composition during the development of African walnut (Tetracarpidium conophorum) seeds. World Applied Sciences Journal 35 (7):1174-1179. |
|
|
Vanden Heuvel JP, Belda BJ, Hannon DB (2012). Mechanistic examination of walnuts in prevention of breast cancer. Nutrition and Cancer 64(7):1078-1086. |
|
|
Verheij EWT (2002). Dacryodes edulis (G.Don) H.J.Lam. [Internet] Record from Protabase. In: PROTA, Oyen L.P.A. and R.H.M.J.Lemmens [Eds]. Plant Resources of Tropical Africa, Wageningen, Netherlands. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


