The aim of this paper is to evaluate gas exchanges and yield of Prata-type banana plants subjected to fertilizer sources for organic management in soil with improved fertility. Two cultivars (‘Dwarf-Prata’ and ‘BRS Platina’), 5 K2O rates (0, 200, 400, 600, and 800 kg ha-1 year-1) supplied by cattle manure and Naturalplus® rock powder, and 2 evaluation times (8:00 am and 2:00 pm) were arranged in a randomized complete block design with 15 replicates. As for production, a randomized block design with 4 cycles and 3 replicates was used. In fertile soils, gas exchanges are little influenced by fertilization. Fertilization with increasing application rates determines a quadratic variation in stomatal conductance and internal CO2 concentration in ‘Dwarf-Prata’ and a linear variation for leaf temperature and instantaneous water-use efficiency in ‘BRS Platina’. Quantum efficiency of photosynthesis is higher at 8:00 am, whereas at 2:00 pm, for leaf temperature. Photosynthesis and leaf transpiration directly correlate with stomatal conductance, instantaneous water-use efficiency, and leaf temperature, while instantaneous water-use efficiency and photosynthesis inversely associate with transpiration and leaf temperature. Cattle manure and rock powder fertilizations do not increase yield in banana plants grown in soils with improved fertility.
Bananas are grown in a variety of climatic zones, such as the semiarid tropics and under different abiotic stresses, which limit their production (Donato et al., 2016). Improving production under these conditions requires, besides plant breeding, studies on management practices that enable higher resilience across the soil-water-plant-atmosphere interactions. Among these practices, natural and organic fertilizations increase diversity and biological activity and promote suppression of pathogens (Geense et al., 2015).
‘BRS Platina’, which is derived from ‘Dwarf Prata’ through hybridization, exhibits comparative advantages, such as resistance to Yellow Sigatoka Leaf Spot and Panama Disease, as well as high-quality fruits that are very similar to those of its genitor. According to Silva et al. (2011), a safe recommendation of a new cultivar requires its evaluation in different environments, regions, and repetitions of production cycles, aiming at the identification of cultivars, as it allows establishing a genotypic variation of physiologic responses under local conditions (Turner et al., 2007). These variations allow extrapolation of these results to be used in the management of specific production systems and to properly manage cultivars under similar conditions (Arantes et al., 2016).
Rock powder or rock dust, a low-cost fertilizer in which there are K, P, Ca, and other nutrients, is used to replenish or to fertilize several types of soils (Harley and Gilkes, 2000). It slowly releases nutrients into the soil, particularly in soils with high pH; furthermore, its use in combination with cattle manure, and other organic materials, enhances its efficiency (Osterroht, 2003).
Various studies on organic fertilization in banana plants (Damatto Júnior et al., 2011; Moniem et al., 2008; Ribeiro et al., 2013) verified the possibility of using these sources; however, studies on physiological characteristics in fertilizer-related trials are needed as the aforementioned studies are predominantly on Cavendish cultivars (Robinson and Gálan Saúco, 2012) even though there are some available results for Prata-type cultivars (Arantes et al., 2016), where cultivars and irrigation systems were studied.
Fertilizations that are based on organic materials might enable nutritional improvements in banana plants. Therefore, this paper aimed to evaluate gas exchanges and yield of ‘Dwarf-Prata’ and ‘BRS Platina’ banana plants subjected to fertilizer sources for organic management in soils with improved fertility.
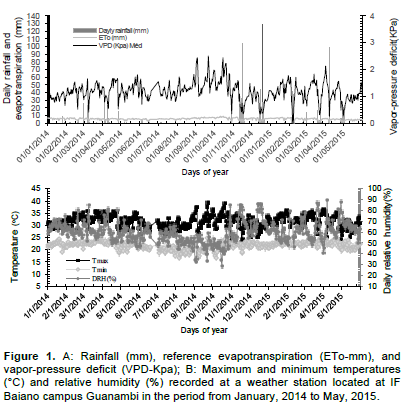
The experiment was carried out in area at Federal Institute of Bahia (IF Baiano), campus Guanambi, BA. Originally, the soil was classified as a typical dystrophic yellow-red Latosol (Oxysol), weak A horizon, medium texture, located at latitude 14°17’27’’S, longitude 42°46’53’’W, altitude of 537 m, average annual rainfall and temperature of 680 mm and 26°C, respectively (Aw climate-Köppen), whose weather variables recorded over the experimental period are as shown in Figure 1.
The experiment was established with micro-propagated plantlets in a spacing of 2.5 × 2.0 m with crop practices consonant with the recommendations of Rodrigues et al. (2015). The area was sub-soiled, plowed, harrowed, furrowed, and the fertilizers were incorporated into the planting hole, according to the treatment.
The irrigation method was micro-sprinkler with NetafimTM emitters (pressure compensating model, Netafim Israel, KibutzHatzerim, Israel), 130 L h-1 flow rate, wetted diameter of 7.4 m, with both laterals and emitters spaced out in 5 m apart. The irrigations were performed based on the crop evapotranspiration, through to the product of the reference evapotranspiration (ETo), calculated by the modified Penman-Monteith method, and the crop coefficient, which varied with the phenological stage in the first cycle and became a fixed value of 1.4 from the flowering, in accordance with Coelho et al. (2012).
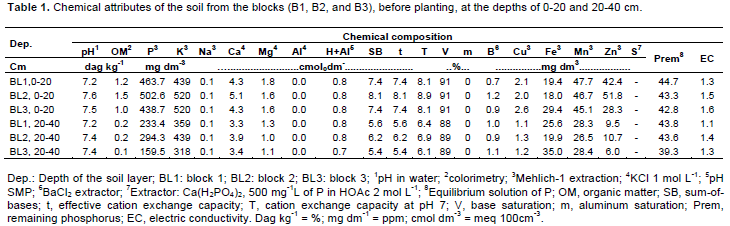
The treatments were 2 cultivars (‘Dwarf-Prata’ and ‘BRS Platina’), 5 K2O rates (0, 200, 400, 600, and 800 kg ha-1 year-1) supplied by cattle manure and Naturalplus®rock powder, and 2 times for gas exchanges evaluation (8:00 am and 2:00 pm) were arranged in a randomized complete block design, in a 2×5×2 factorial experiment, with 15 replicates. As for production, a randomized block design was used, with 4 production cycles, in a 2×5×4 factorial experiment and 3 replicates. The experimental plots consisted of 20 plants, from which the 6 plants in the middle of the plot were the measurement plants. Before the onset of the experiment, soil samples were collected from each experimental block, whose chemical attributes exhibit high fertility (Table 1) due to man-made modifications.
In average, the manure used, on a dry basis (65°C), contained 16.72% of moisture, 63.73 g kg-1 of organic matter, and the following macronutrients (g kg-1): Ca = 1.7; Mg = 0.2; K = 2.5; N = 5.2; S = 2.3 (EPA 3051 / APHA 3120B) and P (APHA 4500-PC); and micronutrients (mg kg-1): B = 2.1; Cu = 45.2; Zn = 200.5; Mn = 391.8; and Fe = 1.932.4 (EPA 3051 / APHA 3120B). pH was 7.42 (Official Method, OM), and the density, 0.38 g cm-3. The Naturalplus®rock powder (natural fertilizer), which was from a soil in Ipirá-Bahia State, Brazil, produced by Terra Produtiva Mineradora Ltd., contains 30.0 g kg-1 of K2O (total), 10.0 g kg-1 of P2O5, 52.0 g kg-1 of CaO, 30.0 g kg-1 of MgO, 63.0 g kg-1 of Fe2O3, 1.5 g kg-1 of MnO, 630 g kg-1 of SiO2, 69 mg kg-1 of Zn (ICP95A – Lithium metaborate fusion – ICP OES), 127 mg kg-1 of Cu and 5 mg kg-1 of OM (IMS95A – Lithium metaborate fusion – ICP MS).
The rates were set on the basis of the maximum recommendation for N found in the literature (Souto et al., 1997), 700 kg ha-1 year-1 of N. From this rate, 5 rates were defined, with an interval of 175 kg ha-1 year-1 from one rate to another, down to a fertilizer rate of 0 (700, 525, 350, 175, and 0 kg ha-1 year-1 of N). The maximum K2O rate was fixed at 800 kg ha-1 year-1 with intervals of 200 kg ha-1 year-1 from one rate to another (800, 600, 400, 200, and 0 kg ha-1 year-1 of K2O). the N/K2O ratio was fixed at 1.7/1. The manure rate (160 Mg ha-1year-1) was defined to meet the demand of 700 kg ha-1 year-1 of N. The amount of K2O (405 kg ha-1 year-1) was calculated on the basis of the manure rate. Based on the K2O content in the rock powder, the rock powder rate was calculated (13 Mg ha-1 year-1) to supply 395 kg ha-1 year-1 of K2O to complement the requirement of 800 kg ha-1 year-1 of K2O. All fertilizer rates were split into 6 applications, every 60 days, distributed to 2,000 plants ha-1. Additionally, 10 g of zinc sulfate and 10 g of boric acid were applied to each mother plant/sucker via rhizome (Rodrigues et al., 2015). Concerning the second cycle, copper sulfate (3 g/mother plant-sucker) split into 3 rates, and 30 g of magnesium sulfate.
Always, 3rd or 4th leaf was measured, in case the 3rd leaf is of difficult access or damaged by the wind, from tip to base (Arantes et al., 2016), the incident radiation on leaf (Oleaf) expressed in µmol; leaf temperature (Tleaf), °C; internal CO2 concentration (Ci), µmol CO2 mol-1, stomatal conductance (gs), mol H2O m-2s-1, transpiration (E), mmol H2O m-2s-1, net photosynthesis (A), µmol CO2 m-2s-1, instantaneous water-use efficiency (A/E), µmol CO2 m-2s-1/mmol H2O m-2s-1, carboxylation efficiency (A/Ci), quantum efficiency or photochemistry of photosynthesis (A/Qleaf), µmol CO2 m-2s-1/µmol photons m-2s-1. These measurements were performed by anLcpro+® Portable Photosynthesis System (ADC BioScientific Limited, UK) infrared gas analyzer (IRGA), always with the radiation shield facing the sun, with ambient temperature and irradiance, and airflow of 200 ml min-1.
For the statistical analysis, the following procedure was adopted; for physiological characteristics, a 2×5×2 factorial design was used; 2 cultivars, 5 cattle manure rates + rock powder, and 2 times for gas exchange readings, with 15 replicates (evaluation season was: from January, 2014-flowering of 2nd cycle, 510 days after transplanting to May, 2015-flowering of the 4th cycle, 900 days after transplanting: periods of higher physiologic maturity and production); as for yield of the bunches and hands, a 2×5×4 factorial design, with 4 production cycles. The data were subjected to analysis of variance and proceeded to split the interactions according to their significance. For the interactions and independent effect of K2O rates, regression analysis was performed. In the absence of interactions, the means for cultivars and times were compared to one another by the omnibus F test. As for cycles, Tukey’s test was done at 5% of significance level. Moreover, correlation studies were done across the variables of gas exchanges.
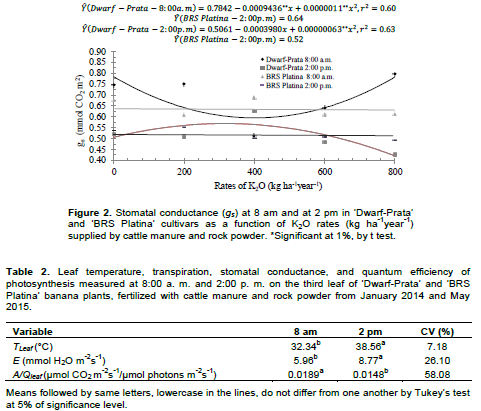
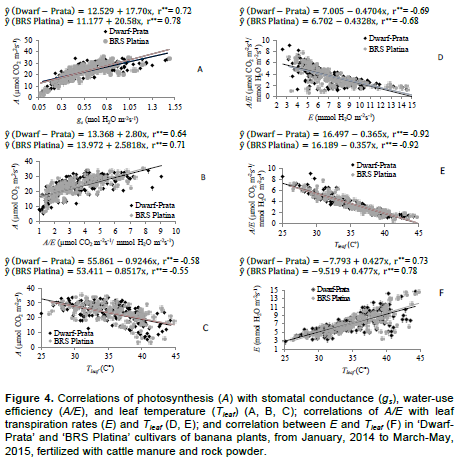
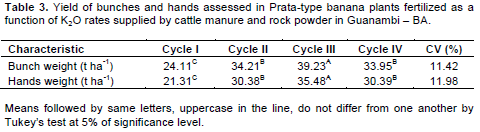
Gas exchanges of the 3rd leaf of Prata-type banana plants had three-way interactions, considering cultivar, fertilizer rate, and time with stomatal conductance (Figure 2), and two-way interactions, considering cultivar and fertilizer rate with leaf temperature, instantaneous water-use efficiency, and transpiration (Figure 2). Leaf temperature, transpiration, and quantum efficiency of the photosynthesis in Prata-type banana plants varied with evaluation time, regardless of the cultivar or fertilizer rate (Table 2). The associations between gas exchanges in Prata-type banana plants were moderated and positive for A×gs and A×A/Ee, and moderated and negative for A×Tleaf (Figure 4). The correlations A/E×Tleaf, A/E×E, and E×Tleaf were of high magnitude and negative, medium magnitude and negative, and medium magnitude and positive, respectively; and are as shown in Figure 4. The bunch and hand weights interacted with cultivar and fertilizer rate and varied with evaluation season in an independent manner.
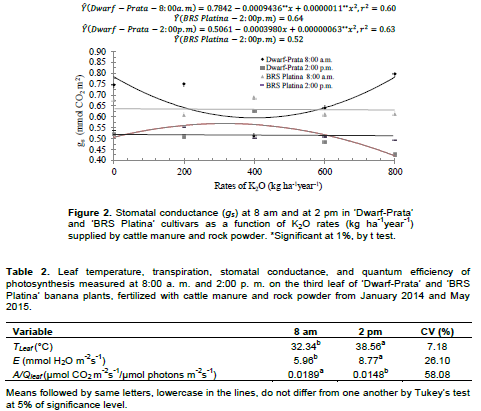
Few physiological variables were influenced by the treatments. This is probably due to the high soil fertility at the beginning of the experiment, whose average P and K contents before planting were 468.33 and 493 mg dm-3, respectively. Also, doing the physiological readings at specific times contributed to this outcome as these readings vary with the atmospheric conditions and soil moisture at the time of measurement, which might not reflect the conditions imposed by the treatments (Santos et al., 2013; Arantes et al., 2016). The ‘Dwarf-Prata’ cultivar, at 8:00 am exhibited a decreasing quadratic behavior for gs rates (Figure 2). The model estimates a minimum value of 0.59 mol H2O m-2s-1 for gs when 401.13 kg ha-1year-1 is applied and rates of 0.78 and 0.80 mmol H2O m-2s-1 in the absence of fertilization and when the maximum fertilizer rate is applied, respectively. As for 2:00 pm, the behavior was inverse, rising at first, up to a maximum conductance of 0.57 mol H2O m2s1 when 315.47 kg ha-1year-1 of K2O is applied, followed by a decrease, down to 0.4206 mol H2O m-2s-1 for the maximum fertilizer rate. Regarding ‘BRS Platina’ cultivar, at the two reading times, the average of gs rates were 0.64 and 0.52 mol H2O m-2s-1, respectively, without adjusting the curve.
The amounts of N and K2O supplied by cattle manure and rock powder contributed toward explaining the gs rates at both reading times. The large quantity of nitrogen and potassium supplied by manure and rock powder could have promoted a better osmoregulation of the stomata opening of leaf cells. This improvement is due to a better cooling of the plant as organic compounds in the soil enhance the capacity of water storage in the soil and, consequently, a better diffusive flux of nutrients (Malavolta et al., 1997).
Melo et al. (2009), using N and K rates in fertigation, verified values of up to 0.51 mol H2O m-2s-1 of gs for K2O rates of 580 kg ha-1year-1 and lower values when the amount of K2O is reduced and N is increased. This evidences the importance of the correct calibration of N and K rates for stomatal osmoregulation of the banana plant and the excess of N and absence of K reduce gs. Even though the N/K2O ratio was fixed at 1.7/1 herein, the amount of N applied was much increased. Other factors, such as the increase in vapor-pressure deficit (VPD), have also an influence on the rates of gs in leaves and, consequently, on the stomatal opening and closure, mainly in periods of higher stress, that is, 2:00 pm (Turner et al., 2007; Mahouachi, 2009; Dzomeku et al., 2016).
Under harsher conditions, the stomata close because of the uptake of K by the guard cells, which is induced by the production of ABA in the roots and, afterwards, by extruding malate by guard cells (Shimazaki et al., 2007). This is caused by the increase in VPD, which is a consequence of low relative humidity, irradiation, and high air temperature on the leaves (Ekanayake et al., 1994; Turner et al., 2007; Donato et al., 2016). Under seasons with more weather stress, such as September and October, the maximum ambient temperature over the period reached up to 39.2°C, relative humidity, from 30 to 35%, which characterizes a condition of high VDP, with recorded values above 2.5 kPa (Figure 2) and the incident radiation on the leaf reached up to 1.998 µmol photons m-2 s-1.
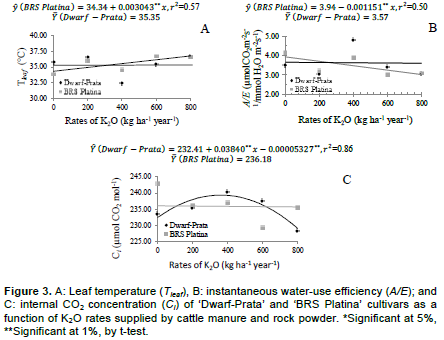
These weather conditions are above what is considered to be optimum for the photosynthesis of the banana plant (Robison and Galán Saúco, 2012), and the decrease in photosynthetic rates might occur because of the stomatal closure or enzymatic problems, leading to decreases in carboxylation efficiency (Arantes et al., 2016). Conversely, under milder conditions, potassium accumulates in cell guards; as a result, their osmotic potential increases, and in addition to water, their turgidity is also increased, which favors the stomatal opening; therefore, the positive quadratic behavior at 8:00 am is explained. Leaf temperature (Tleaf) linearly increased (p<0.05) for ‘BRS Platina’ cultivar. There was an increase of 0.003043°C for each kg of K2O applied, adding up to an increase of 8.1% for the maximum rate in comparison with no fertilized treatments. As for the ‘Dwarf-Prata’ cultivar, the model was not adjusted. The average Tleaf was 35.35°C (Figure 3 A). However, as the application of manure and rock powder maintained the soil moisture, as well as supplying K, which, together, helped in maintaining the stomatal activity for a longer time; thus, the cooling of leaves was favored, which decreased the possibility of thermal damage on the leaves.
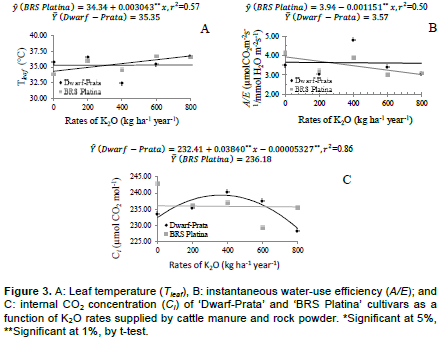
Al-Busaidi (2015) verified that plants fertilized with cattle manure had lower electrolyte loss in leaf tissues as a function of the increase in temperature, which means that there were fewer changes than in conventional management; such fact can be attributed to the compost’s capacity of supplying nutrients over a long time, such as Ca, important in cell membranes, and K, in the osmotic regulation. Besides, the osmotic regulation, K is associated with folding of the 2 halves of the lamina downwards; thus, reducing the stress caused by temperature and sunlight (Soto Ballestero, 2008). From a physiological standpoint, as leaf temperature increases, both instantaneous water-use efficiency and internal CO2 concentration decrease. This is observed in Figure 3B and C, respectively. The air temperature (Figure 1), which also affects the Tleaf (Donato et al., 2017), could have affected the variables A/E and Ci; nevertheless, under these conditions, there would be a lower stomatal activity, which, perhaps, did not occurred right away because the average stomatal conductance was kept high (above 0.50 mol H2O m-2 s-1), as shown in Figure 2. The maintenance of the soil moisture, promoted by the crop residue left on the ground and by fertilizers applied every 60 days, might justify the high values of soil moisture.
The instantaneous water-use efficiency (A/E) of the ‘BRS Platina’ cultivar linearly decreased (p<0.05) as a function of K2O rates supplied by cattle manure and rock powder. The decrease was of -0.00115 µmol CO2 m-2 s-1/mmol H2O m-2 s-1 for each kg of K2O applied. As for ‘Dwarf-Prata’ cultivar, a model was not adjusted; its average was 3.57 µmol CO2 m-2 s-1/mmol H2O m-2 s-1 (Figure 3B). The increase in leaf temperature, as observed in Figure 3A, caused a decrease in A/E (Donato et al., 2016), even when using a proper irrigation depth and a higher application of organic fertilizer. The fact that the experiment was conducted under semiarid conditions where the increases in temperature and background radiation are common and persistent in certain periods of the year, contributed much more to the lower A/E rates as carboxylation efficiency of rubisco decreases, which was a consequence of the increase in transpiration due to high temperatures (Arantes et al., 2016; Donato et al., 2017).
In banana orchards, where the management of crop residue in combination with the application of organic fertilizers contributes to higher rooting and enhanced chemical, physical, and biological attributes of the soil (Donato et al., 2016), there is an alleviation of conditions with excess of sunlight and high temperatures. As a consequence, the supply of K and N from manure and rock powder is essential as this supply is related to the water and osmotic regulation of the plant and protects against abiotic factors (Shimazaki et al., 2007). Melo et al. (2009) indicate that the increase in potassium fertilization provides the banana plant with lower N/K ratios and contributes to the maintenance of A/E. The internal CO2 concentration (Ci) adjusted an increasing quadratic model for ‘Dwarf-Prata’ cultivar (Figure 3C) as a function of K2O rates supplied by cattle manure and rock powder. A maximum Ci of 239.34 µmol CO2 mol-1 was observed for an application of 360.42 kg ha-1 year-1, and from this rate on, the Ci decreased. As for ‘BRS Platina’ cultivar, there was no adjustment to the curve with regard to the observed phenomenon; its internal CO2
concentration was 236.18 µmol CO2 mol-1.
Melo et al. (2009) recorded increases in Ci as the stomatal opening rate increases. This is due to the fact that during gas exchange processes, stomata regulate the Ci and keep it relatively constant; during the stomatal closure process, the stomata restrain the uptake of CO2 and, consequently, the photosynthetic activity (Farquhar and Sharkey, 1982).
The growing presence of N and K2O as a function of fertilizer rates might have influenced the initial increase in Ci of ‘Dwarf-Prata’ cultivar, followed by a fall in the cultivars since these nutrients are related to the opening and closure of stomata. The rise in leaf temperature caused by the weather (Figure 1) could also have influenced the decrease in Ci since the Tleaf has influence on opening and closing stomata, on functioning of enzymes, and on transpiration (Turner et al., 2007; Donato et al., 2016).
The Tleaf, E, and A/Qleaf rates differed from one another by the F-test (p<0.05) between 8:00 am and 2:00 pm, regardless of the cultivar and K2O rates (Table 2). At 2:00 pm, the Tleaf and E rates were higher and the opposite effect occurred at 8:00 am for A/Qleaf rates. The Tleaf and E exhibited, respectively, an increase of 16.13 and 32.34% between 8:00 am and 2 pm. Donato et al. (2016) and Arantes et al. (2016) observed higher rates for these same variables, at 2 pm as well. The increment in Tleaf is linked to the increase in ambient temperature between the period from 8:00 am to 2:00 pm, as ascertained for the ‘Dwarf-Prata’ clone called ‘Gorutuba’ (Donato et al., 2017). The temperature also influences the E as it is affected by the two temperatures (Donato et al., 2016). The reduction in quantum efficiency of the photosynthesis (Table 2) between the 2 times is because of the increase in vapor-deficit pressure in the afternoon and the change in the radiation quality, with wave lengths more suitable for photosynthesis at the beginning of the morning in comparison with the afternoon.
The VPD is influenced by the rise in ambient temperature and sunlight, and reduction in relative humidity, which naturally occur over the day and peak at 2 pm; this is verified by observing the other variables, Tleaf and E (Table 2). In addition to the VPD, the incident radiation on the leaf at 2 pm, in combination with the increase in leaf temperature, could have partially restrained the rubisco, which led to the decrease in CO2 fixation, which culminated, therefore, in reducing the A/Qleaf. There is a direct relationship between A and gs (Figure 4A), whose correlation coefficients were 0.72 and 0.78 for ‘Dwarf-Prata’ and ‘BRS Platina’ and 0.64 and 0.71 for A and A/E (Figure 4B) for the two cultivars, respectively. Moreover, the correlation between A and Tleaf (Figure 4C) occurred; however, in an inverse way and of moderate magnitude, with coefficients of 0.58 and -0.55 for ‘Dwarf-Prata’ and ‘BRS Platina’ cultivars, respectively, which agrees with Arantes et al. (2016).
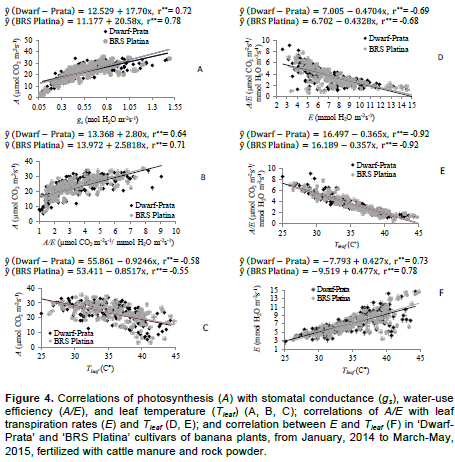
As for the correlation A×gs of ‘Dwarf-Prata’ and ‘BRS Platina’, respectively, there are increases of 17.70 and 20.58 units in A for each increment in gs. Similarly, for A×A/E, the cultivars has increases of 2.80 and 2.58 units in A for each increment in A/E. As for the correlation A×Tleaf, there were decreases of 0.9246 and 0.8517 units in A for each increment in Tleaf. Correlation studies, with inverse relationship, were also performed for A/E × E (Figure 4D), whose correlation coefficients were -0.69 and -0.68 for both cultivars; A/E × Tleaf (Figure 4E), whose correlation coefficients were -0.92 for both cultivars and direct association for E × Tleaf (Figure 4F) with coefficients of 0.73 and 0.78 for ‘Dwarf-Prata’ and ‘BRS Platina’, respectively.
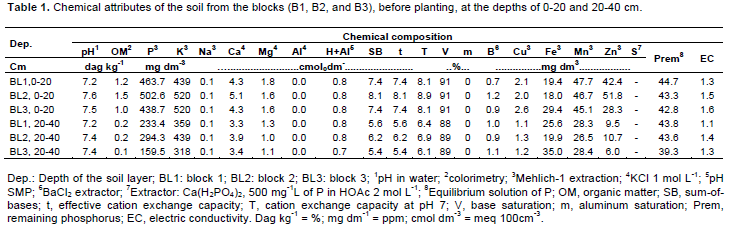
As for the correlation A/E × E, there are decreases of -0.4704 and -0.4328 units of A/E for each increment in E for ‘Dwarf-Prata’ and ‘BRS Platina’, respectively. As for A/E × Tleaf, the cultivars had decreases of 0.3658 and 0.3574 units in A/E for each increment in Tleaf. In the correlation E × Tleaf, there were increases of 0.427 and 0.477 units in E for each increment in Tleaf. Figure 4D, E, and F demonstrates that the increasing Tleaf, which depends on the air temperature (Donato et al., 2016; 2017), limits the enzymatic activity of rubisco, which decreases the photosynthesis, though, the transpiration rate increases, leading to a reduction in A/E, even under favorable conditions of water availability and fertile soil (Donato et al., 2016). Arantes et al. (2016), who studied ‘Dwarf-Prata’, ‘BRS Platina’, and other cultivars, verified correlation between A/E × Tleaf and E × Tleaf that were similar to those found in this paper. There was a difference across means (p<0.05) for bunch and hand weights when the effect of the cycle in isolation was observed (Table 3). The 3rd cycle was the most productive, with mean value of 39.23 t ha-1 of bunches and 35.48 t ha-1 of hands. The yield averages are corrected for the real yield, which was obtained by multiplying the population effectively harvested (76%) from the density at planting due to losses caused by the wind or other factors that are not related to the treatments.
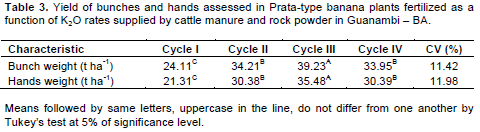
Donato et al., (2015) reported that well-managed banana orchards that were planted in soils with improved fertility exhibit yields above 40 t ha-1 cycle-1 (high yield > 32 t ha-1 cyle-1). Silva and Simão (2015) recorded similar bunch weight when carrying out trials with K fertilization; though, with lower overall yield due to the lower population (1,235 plants ha-1). Damatto Junior et al. (2011), with organic fertilization, reported an increase in bunch weight from the 1st to the 2nd cycle, with lower values than those herein, followed by a reduction in the 3rd cycle, reduction of which was claimed to be due to nutrient depletion of the soil.