Full Length Research Paper

ABSTRACT
This research estimated litter production and analyzed its relation to environmental variables such as maximum temperature, insolation, and rainfall. The study was conducted on a 300 × 300 m experiment as part of the project titled mapping of native Brazil nut stands and socio-environmental and economic characterization of Brazil nut production systems (MapCast), in the Tapajós National Forest (FLONA Tapajós). Every 30 days for one full year (August 2015 to July 2016), litterfall was collected and stored in a laboratory. After drying, the material was separated into leaves, wood, flowers and fruits, and miscellaneous and weighed. Statistical tests conducted were Shapiro-Wilk (5%), Principal coordinate analysis, t-test, Pearson’s linear correlation, cross-correlation, and canonical redundancy analysis. Rainfall and temperature data were inferior and superior, respectively, to normal climate conditions in the region, and data for solar insolation had an abnormal pattern compared to normal climate conditions. Leaf production varied between 169.9 and 965.6 kg ha-1 month-1, and that of wood between 26.7 and 501.3 kg ha-1 month-1, while that for flowers and fruit varied from 0.6 to 19.6 kg ha-1 month-1. The greatest leaf production was measured during the months with the lowest amount of rainfall and highest temperatures, and variation in leaf production and total litterfall was partially explained by temperature and insolation.
Key words: Litter, Bertholletia excelsa, Amazon, El Niño, FLONA Tapajós.
INTRODUCTION
MATERIALS AND METHODS
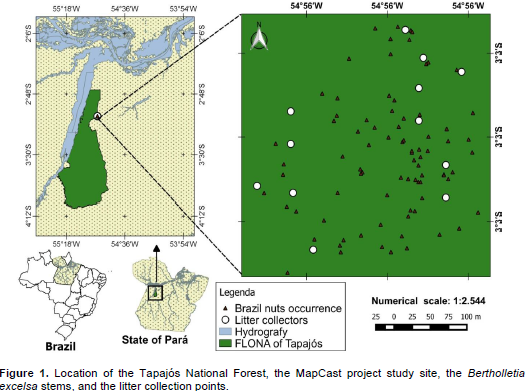
RESULTS
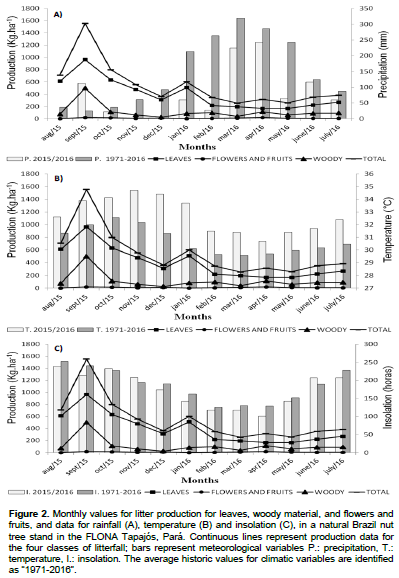
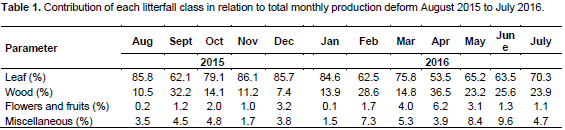
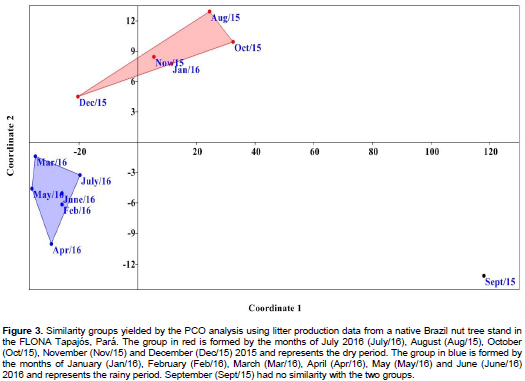
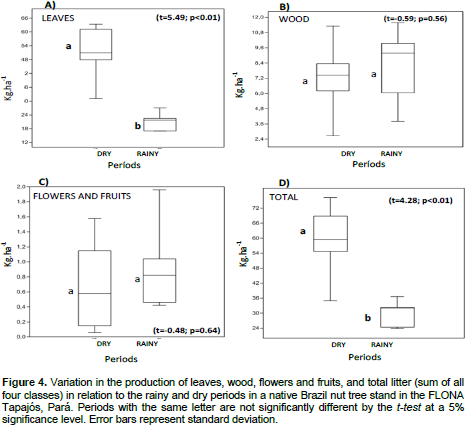
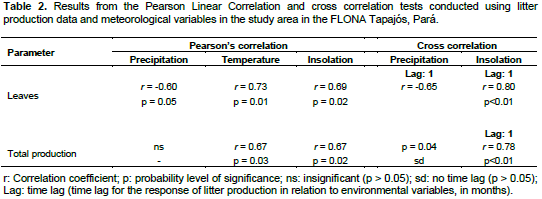
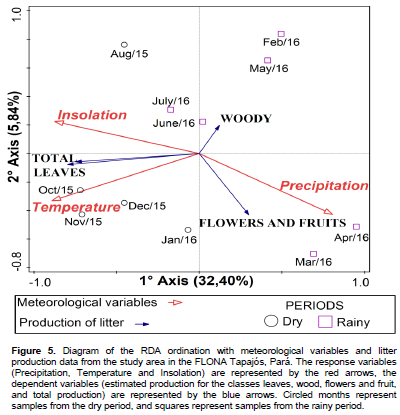
DISCUSSION
CONCLUSION
CONFLICT OF INTERESTS
ACKNOWLEDGEMENT
REFERENCES
|
Almeida EJ, Luizão F, Rodrigues DJ (2015). Produção de serrapilheira em florestas intactas e exploradas seletivamente no sul da Amazônia em função da área basal da vegetação e da densidade de plantas. Acta Amaz. 45(2):157-166. |
|
|
Andrade DF, Gama JRV, Melo LO, Ruschel AR (2015). Inventário florestal de grandes áreas na Floresta Nacional do Tapajós, Pará, Amazônia, Brasil. Biota Amaz. 5(1):109-115. |
|
|
Bi J, Knyazikhin Y, Choi S, Park T, Barichivich J, Ciais P, Fu R, Ganguly S, Hall F, Hilker T, Huete A, Jones M, Kimball J, Lyapustin AI, Mõttus M, Nemani RR, Piao S, Poulter B, Saleska SR, Saatchi SS, Xu L, Zhou L, Myneni RB (2015). Sunlight mediated seasonality in canopy structure and photosynthetic activity of Amazonian rainforests. Environ. Res. Lett. 10:064014. |
|
|
Bianchin JE, Marques R, Britez RM, Capretz RL (2016). Deposição de fitomassa em formações secundárias na Floresta Atlântica do Paraná. Floresta Ambient. 23(4):524-533. |
|
|
Borchert R, Calle Z, Strahler AH, Baertschi A, Magill RE, Broadhead JS, Kamau J, Njoroge J, Muthuri C (2015). Insolation and photoperiodic control of tree development near the equator. New Phytol. 205:7-13. |
|
|
Calvi GP, Pereira MG, Espíndula AJR (2009). Produção de serapilheira e aporte de nutrientes em áreas de floresta atlântica em Santa Maria de Jetibá, ES. Ciênc. Florest. 19(2):13-138. |
|
|
Carvalho JOP (1992). Structure and dynamics of a logged over Brazilian Amazonian rain forest. PhD thesis, University of Oxford, Oxford. 215p. |
|
|
Carvalho JOP (2001). Estrutura de matas altas sem babaçu na Floresta Nacional do Tapajós. In: Silva JNM, Carvalho JPde, Yared JG (Ed.) A silvicultura na Amazônia Oriental: contribuições do projeto Embrapa/DFID. Belém: Embrapa Amazônia Oriental/DFID. pp. 277-290. |
|
|
Cetin M (2015). Evaluation of the sustainable tourism potential of a protected area for landscape planning: a case study of the ancient city of Pompeipolis in Kastamonu. Int. J. Sustain. Dev. World Ecol. 22(6):490-495. |
|
|
Cetin M (2016). Sustainability of urban coastal area management: A case study on Cide. J. of Sustainable Forestry. 35(7):527-541. |
|
|
Chagas GFB, Silva VPR, Costa ACL, Dantas VA (2012). Impactos da redução da pluviometria na biomassa aérea da Floresta Amazônica. Rev. Bras. Eng. Agríc. Ambient. 16(1):72-79. |
|
|
Chave J, Navarrete D, Almeida S, Álvarez E, Aragão LEOC, Bonal D, Châtelet P, Silva-Espejo JE, Goret JY, Hildebrand PV, Jiménez E, Pati-o S, Pe-uela MC, Phillips OL, Stevenson P, Malhi Y (2010). Regional and seasonal patterns of litterfall in tropical South America. Biogeosciences 7(1):43-55. |
|
|
Costa MC, Costa ACL, Coelho LTS, Silva TML, Azevedo AF (2014). Correlação entre precipitação pluviométrica E umidade do solo na produção de serapilheira em Caxiuanã (PA). Rev. Iberoâ€Americana de Ciênc. Ambient. 5(1):170-179. |
|
|
Davis JC (1986). Statistics and Data Analysis in Geology. Nova York: John Wiley & Sons. 646p. |
|
|
Espírito-Santo FDB, Shimabukuro YE, Aragão LEOC, Machado ELM (2005). Análise da composição florística e fitossociológica da Floresta Nacional do Tapajós com o apoio geográfico de imagens de satélites. Acta Amaz. 35(2):155-173. |
|
|
Ferreira LS, Cattânio JH, Jardim MAG (2015). Efeito da topografia e da precipitação na florística e na produção de liteira em Caxiuanã, Pará. Rev. Árv. 39(6):995-1005. |
|
|
Ferreira ML (2014). Incremento diamétrico arbóreo em diferentes grupos funcionais e produção de serapilheira em duas florestas brasileiras. PhD thesis, University of São Paulo, São Paulo. 201p. |
|
|
Giácomo RG, Pereira MG, Machado DL (2012). Aporte e decomposição de serapilheira em áreas de Cerradão e Mata Mesofítica na Estação Ecológica de Pirapitinga – MG. Ciênc. Florest 22(4):669-680. |
|
|
Godinho TO, Caldeira MVW, Brun EJ (2015). Ciclagem de nutrientes via serapilheira em ecossistemas florestais naturais no Brasil. In: Faria ÁBdeC, Brun EJ, Ferrari F (Org). Ciências Florestais e Biológicas. Curitiba: UTFPR. pp. 13-52. |
|
|
Godinho TO, Caldeira MVW, Rocha JHT, Caliman JP, Trazzi PA (2014). Quantificação de biomassa e nutrientes na serapilheira acumulada em trecho de Floresta Estacional Semidecidual Submontana, ES. Cerne. 20(1):11-20. |
|
|
Golden GWS (2016). El Ni-o and La Ni-a Years and Intensities Based on Oceanic Ni-o Index (ONI). Accessed in: http://ggweather.com/enso/oni.htm. |
|
|
Golley FB (1978). Ciclagem de minerais em um ecossistema de floresta tropical úmida. Editora da Universidade de São Paulo: São Paulo. 105p. |
|
|
Gonçalves FG, Santos JR (2008). Composição florística e estrutura de uma unidade de manejo florestal sustentável na Floresta Nacional do Tapajós, Pará. Acta Amaz. 38(2):229-244. |
|
|
Guerreiro QLM, Oliveira Júnior RC, Santos GR, Ruivo MLP, Beldini TP, Carvalho EJM, Silva KE, Guedes MC, Santos PRB (2017). Spatial variability of soil physical and chemical aspects in a Brazil nut tree stand in the Brazilian Amazon. Afr. J. Agric. Res. 12(4):237-250. |
|
|
Hammer O, Haper DAT, Ryan PD (2001). Past: paleontological statistics software package for education and data analisys. Palaeontol. Eletr. 4:1-9. |
|
|
Hayashi SN (2006). Dinâmica da serapilheira em uma cronossequencia de florestas no município de Capitão Poço-Pa. MSc thesis, University Rural of da Amazônia, Belém 75p. |
|
|
Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis (IBAMA) (2004). Floresta Nacional do Tapajós – Plano de Manejo. Belterra: IBAMA, 373p. |
|
|
Intituto Nacional de Meteorologia (INMET) (2010). Ocorrência de La nina e El Ni-o. Accessed in: View. |
|
|
Kapos V, Wandelli E, Camargo JL, Ganade G (1997). Edge-related changes in environmental and plant responses due to forest fragmentation in central Amazonia. In: Laurance WF, Bierregard Jr RO (Eds.). Tropical forest remnants: ecology, management, and conservation of fragmented communities. Chicago: University of Chicago Press pp. 33-44. |
|
|
Lima RP, Fernandes MM, Fernandes MRM, Matricardi EAT (2015). Aporte e decomposição da serapilheira na Caatinga no Sul do Piauí. Floresta Ambient. 22(1):42-49. |
|
|
Locatelli M, Vieira AH, Martins EP, Souza VF, Macedo RS (2005). Crescimento em diâmetro de castanha-do-brasil (Bertholletia excelsa H.B.K.) cultivada em solo de baixa fertilidade. PortoVelho: Embrapa 4 p. |
|
|
Loureiro AA, Silva MF, Alencar JC (1979). Essências florestais madeireiras da Amazônia. Manaus: INPA. 245 p. |
|
|
Malhi Y, Saatchi S, Girardin C, Aragão LEOC (2009). Produção, estoques e fluxo de carbono nas florestas amazônicas. Geophys. Monogr. 186:355-371. |
|
|
Maués MM (2002). Reprodutive phenology and pollination of the Brazil nut tree (Bertholletia excelsa Humb. & Bompl. Lecythidaceae) in eastern Amazonia, In: Kevan P, Imperatriz-Fonseca VL (Ed.). Pollination bees: the conservation link between agriculture and nature. Brasília: Ministério do Meio Ambiente pp. 245-254. |
|
|
Meir P, Brando PM, Nepstad D, Vasconcelos S, Costa ACL, Davidson E, Almeida S, Fisher RA, Sotta ED, Zarin D, Cardinot G (2009). Os efeitos da seca nas florestas chuvosas Amazônicas. Geophys. Monogr. 186:429-449. |
|
|
Mochiutti S, Queiroz JAL, Melém Junior NJ (2006). Produção de serapilheira e retorno de nutrientes de um povoamento de taxi-branco e de uma Floresta Secundária no Amapá. Bol. Pesqui. Florest. 52:3-20. |
|
|
Moraes RM (2002). Ciclagem de nutrientes na floresta do PEFI: produção e decomposição da serapilheira. In: Bicudo D, Forti M, Bicudo C (Ed.). Parque Estadual das Fontes do Ipiranga, unidade de conservação que resiste à urbanização de São Paulo. São Paulo: Secretaria de Estado do Meio Ambiente pp. 133-142. |
|
|
Mori SA, Prance GT (1990). Taxonomy, ecology, and economic botany of the Brazil nut (Bertholletia excelsa Humb. & Bonpl.: Lecythidaceae). Adv. Econ. Bot. 8:130-150. |
|
|
O'brien JJ, Oberbauer SF, Clark DB, Clark DA (2008). Phenology and stem diameter increment seasonality in a Costa Rican wet tropical forest. Biotrópica 40(2):151-159. |
|
|
Ourique LK, Silva RO, Souza CAS, Higushi N (2016). Relação da produção de serapilheira com incremento em diâmetro de uma floresta madura na Amazônia Central. Scientia Forestalis. 44(112):875-886. |
|
|
Parolin P, Lucas C, Piedade MTF, Wittmann F (2010). Drought responses of flood-tolerant trees in Amazonian floodplains. Ann. Bot. 105(1):129-139. |
|
|
Pinho GSC, Fiedler NC, Lisbôa CDJ, Rezende AV, Martins IS (2004). Efeito de diferentes métodos de corte de cipós na produção de madeira em tora na Floresta Nacional do Tapajós. Ciênc. Florest. 14(1):179-192. |
|
|
Proctor J (1983). Tropical Forest Litterfall, In: Sutton SL, Whitmore TC, Chadwick AC (Ed.) Tropical rain forest: Ecology and management. Oxford: Blackwell Scient. Public. pp. 267-273. |
|
|
Pyle EH, Santoni GW, Nascimento HEM, Hutyra LR, Vieira S, Curran DJ,Haren JV, Saleska SR, Chow VY, Carmago PB, Laurance WF, Wofsy SC (2008). Dynamics of carbon, biomass, and structure in two Amazonian forests. J. Geophys. Res. 113:1-20. |
|
|
Quesada CA, Lloyd1 J, Anderson LO, Fyllas NM, Schwarz M, Czimczik CI (2011). Soils of Amazonia with particular reference to the RAINFOR sites. Biogeosciences 8(6):1415-1440. |
|
|
Restrepo-Coupe N, Rochac HR, Hutyra LR, Araujo AC, Borma LS, Christoffersen B, Cabral OMR, Camargo PB, Cardoso FL, Costa ACL, Fitzjarrald DR, Goulden ML, Kruijt B, Maia JMF, Malhi YS, Manzi AO, Miller SD, Nobre AD, Randow CV, Sá LDA, Sakai RK, Tota J, Wofsy SC, Zanchi FB, Saleskaa SR (2013). What drives the seasonality of photosynthesis across the Amazon basin? A cross-site analysis of eddy flux tower measurements from the Brasil flux network. Agric. For. Meteorol. 82(1):28-44. |
|
|
Rivera G, Elliott S, Caldas LS, Nicolossi G, Coradin VT, Borchert R (2002). Increasing day-length induces spring-flushing of tropical dry forest trees in the absence of rain. Trees 16: 445-456. |
|
|
Salman AKD, López GFZ, Bentes-Gama MM, Andrade CMS (2008). Espécies arbóreas nativas da Amazônia Ocidental Brasileira com potencial para arborização de pastagens. Porto Velho: Embrapa. (Boletim técnico, 127). 20p. |
|
|
Salomão RP (2014). A castanheira: história natural e importância socioeconômica. Bol. Mus. Para. Emílio Goeldi, sér. Ciênc. Nat. 9(2):259-266. |
|
|
Santos Junior UM (2008). Fisiologia e indicadores de estresse em árvores crescendo em ambientes alagados pela hidroelétrica de Balbina na Amazônia Central. PhD thesis, National Institute for Space Research, Manaus. 144p. |
|
|
Santos Neto AP, Barreto PAB, Gama-Rodrigues EF, Novaes AB, Paula A (2015). Produção de serapilheira em floresta estacional semidecidual e em plantios de Pterogyne nitens Tul. E Eucalyptus urophylla S. T. Blake no sudoeste da Bahia. Ciênc. Florest. 25(3):633-643. |
|
|
Scoles R (2010). Ecologia e extrativismo da castanheira (Bertholletia excelsa, lecythidaceae) em duas regiões da Amazônia brasileira. PhD thesis, National Institute for Space Research, Manaus. 209p. |
|
|
Scoles R, Canto MS, Almeida RG, Vieira DP (2016). Sobrevivência e frutificação de Bertholletia excelsa Bonpl. em áreas desmatadas em Oriximiná, Pará. Florest. Ambient. 23(4):555-564. |
|
|
Scoriza RN, Pereira MG, Pereira GHA, Machado DL, Silva EMR (2012). Métodos para coleta e análise de serrapilheira aplicados à ciclagem de nutrientes. Florest. Ambient. 2(2):1-18. |
|
|
Sevik H, Cetin M (2015). Effects of water stress on seed germination for select landscape plants. Pol. J. Environ Stud. 24(2):689-693. |
|
|
Silva AD (2014). Produção e concentração de nutrientes via deposição de liteira na Floresta Nacional do Tapajós, Belterra – PA. MSc thesis, University of West of Pará, Santarém 90p. |
|
|
Silva AD, Oliveira Junior RC (2010). Produção de liteira na Floresta Nacional do Tapajós no ano de 2007. Espaço Científico 11(1/2):1-12. |
|
|
Silva RM, Costa JMN, Ruivo MLP, Costa ACL, Almeida SS (2009). Influência de variáveis meteorológicas na produção de liteira na Estação Científica Ferreira Penna, Caxiuanã, Pará. Acta Amaz. 39(3):573-582. |
|
|
Silva WR (2013). Produção de liteira final em uma área de contato capinarama-Floresta Ombrófila na Amazônia setentrional. MSc thesis, University of Roraima, Boa Vista 43p. |
|
|
Ter Braak CJF, Smilauer P (2002). Canoco reference manual and canodraw for Windows user's guide, software for canonical comunity ordination (version 4.5). Netherlands: Biometris, Wageningen and Ceske Budejovice 500p. |
|
|
Valentin JL (2012). Ecologia Numérica: Uma introdução à análise multivariada de dados ecológicos. Rio de Janeiro: Interciência 168 p. |
|
|
Vasconcellos RLF, Bini D, Paula AM, Andrade JB, Cardoso EJBN (2013). Nitrogênio, carbono e compactação do solo como fatores limitantes do processo de recuperação de matas ciliares. Rev. Bras. Ciênc. Solo 37:1164-1173. |
|
|
Veloso HP, Rangel Filho AL, Lima JCA (1991). Classificação da vegetação brasileira, adaptada a um sistema universal. Rio de Janeiro: IBGE. |
|
|
Wagner FH, Hérault B, Bonal D, Stahl C, Anderson LO, Baker TR, Becker GS, Beeckman H, Souza DB, Botosso PC, David MJ, Bowman S, Bräuning A, Brede B, Brown FI, Camarero JJ, Camargo PB, Cardoso FCG, Carvalho FA, Castro W, Chagas RK, Chave J, Chidumayo EN, Clark DA, Costa FRC, Couralet C, Mauricio PHS, Dalitz H, Castro VR, Milani JEF, Oliveira EC, Arruda LS, Devineau JL, Drew DM, Dünisch O, Durigan G, Elifuraha E, Fedele M, Fedele LF, Filho AF, Finger CAG, Franco AC, Freitas Júnior JL, Galvão F, Gebrekirstos A, Gliniars R, Graça PMLA, Griffiths AD, Grogan J, Guan K, Homeier J, Kanieski MR, Kho LK, Koenig J, Kohler SV, Krepkowski J, Lemos-Filho JP, Lieberman D, Lieberman ME, Lisi CS, Santos TL, Ayala JLL, Maeda EE, Malhi Y, Maria VRB, Marques MCM, Marques R, Chamba HM, Mbwambo L, Melgaço KLL, Mendivelso HA, Murphy BP, O'Brien JJ, Oberbauer SF, Okada N, Pélissier R, Prior LD, Roig FA, Ross M, Rossatto DR, Rossi V, Rowland L, Rutishauser E, Santana H, Schulze M, Selhorst D, Silva WR, Silveira M, Spannl S, Swaine MD, Toledo JJ, Toledo MM, Toledo M, Toma T, Filho MT, Hernández JIV, Verbesselt J, Vieira SA, Vincent G, Castilho CV, Volland F, Worbes8 M, Zanon MB, Aragão LEOC (2016). Climate seasonality limits leaf carbon assimilation and wood productivity in tropical forests. Biogeosciences 13:2537-2562. |
|
|
Yoon JH, Zeng N (2010). An Atlantic influence on Amazon rainfall. Clim. Dyn. 34:249-264. |
|
|
Zar JH (1999). Biostatistical analysis. New Jersey: Prentice Hall, Upper Saddle River 662p. |
|
|
Zhang H, Yuan W, Dong W, Liu S (2014). Seasonal patterns of litterfall in forest ecosystem worldwide. Ecol. Complex. 20:240-247. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


