Full Length Research Paper

ABSTRACT
The present work focused on evaluation of the physiological and metabolic behavior of alfalfa (Medicago sativa L., var. Siriver), when faced with two important ecosystems constraints, namely salinity and flooding-associated hypoxia. After germination, seedlings in symbiosis with Sinorhizobium meliloti were grown on different salt concentrations (0, 20 and 50 mM NaCl). At four weeks post-germination, plants were submitted to a five-day long flooding treatment and then analyzed for various physiological (growth, biomass) and biochemical (proteins, chlorophylls, ions, proline) parameters. Our results showed that when M. sativa was cultured on moderate (20 mM NaCl) or average (50 mM NaCl) salt stress, the classical mechanisms of stress response (ions and proline accumulation) are set up and the growth (FW, DW, growth of aerial parts) was moderately affected by salt. In a situation of average salt stress (50 mM NaCl), the combined salinity and short-term flooding led to an early alteration of the photosynthetic machinery and a modification of ions and proline contents in the leaves.
Key words: Chlorophyll, hypoxia, legume, NaCl, proline, salinity.
INTRODUCTION
Agricultural soils in North Africa are subject to numerous environmental constraints such as salinity, drought during the dry season, flooding or waterlogging during the rainy season, and pollutions caused by industrial and mining discharges (Zahran, 1999). Soil reduced fertility is often due to the presence of salt, and the use of salted water supplies for irrigation resulting in decreased crop productivity (Glenn et al., 1999). Depending on the agricultural region, limitation of water reserves and irrigation practices lead to periodic irrigation of the fields. Thus, either during the rainy seasons or between two irrigation turns, agricultural areas may be flooded or waterlogged for several days, even weeks, at the time of vegetative growth of crops. Both flooding and waterlogging result in hypoxia due to the rapid consumption of oxygen in the rhizosphere and a near 104 reduction in diffusion of oxygen in water relative to air (Armstrong, 1980).
Alfalfa, or lucerne (Medicago sativa L.), is a forage legume cultivated around the world, and among others in North Africa (Bouton, 2012; Zahran, 1999). M. sativa is considered to be relatively tolerant to salinity (Bruning and Rozema, 2013; Zahran, 1999) and drought (Bouton, 2012), but sensitive to flooding-induced hypoxia (Striker and Colmer, 2017). Several studies have analyzed its behavior under either saline stress or flooding-induced hypoxia (Li et al., 2010; Rogers et al., 2008, 2009, 2011; Smethurst et al., 2005), but the effects of the double saline and flooding constraints on its growth have not been investigated. In the present work, we initiated the analysis of young M. sativa (var. Siriver) plant response to moderate/average salinity combined to short-term flooding treatment. This was accomplished through the measurement of biometric and physiological parameters of the aerial part to evaluate the effects of the combined constraint on forage production, as well as concentrations of mineral ions (Na+, K+, Cl-) and proline, chosen as markers of salt-stress response.
MATERIALS AND METHODS
Ionic analysis of irrigation waters of M. sativa cultures
To carry out this study, the salinity of irrigation water of two agricultural soils in northern Tunisia, where alfalfa (M. sativa var siriver) is cultivated, was analyzed. The first site of Ghezala Mateur (37° 05' 02" North, 9° 32' 08" East) is located in the heart of an area of 5,300 ha from the Northwest of the town of Mateur to the South of Lake Ichkeul, and it includes the plain of Mateur and the perimeter of Ghezala, which is fed from the dam of Joumine. The second site of Oum Hani, in the suburb of Menzel Bourguiba (37° 09' 13" North, 9° 47' 09" East) is located in agricultural land close to the lakes of Ichkeul and Bizerte. Electrical conductivity and salinity of irrigation waters during the winter of 2014 were measured as in Bahri (1993). The averaged electrical conductivity (one measurement) and nitrate and NaCl concentrations (3 measurements) of the irrigation water were as follows: 1) water of the Ghezala well: 0.86 dS/m, 0.21 ± 0.04 mM NO3-, 19 ± 5 mM NaCl; 2) water of the Oum Hani well: 4.65 dS/m, 0.92 ± 0.03 mM NO3-, 52 ± 6 mM NaCl.
Based on nitrate and NaCl contents measured in irrigation waters, we chose to test the behavior of M. sativa seedlings under the following salt conditions: 1) a control condition (Ctrl0), with 0.2 mM KNO3 (a low concentration of nitrate which allows a minimum supply of nitrogen without affecting the symbiotic process) in the absence of NaCl, 2) a T20 treatment, with 0.2 mM KNO3 and 20 mM NaCl, representing the irrigation water characteristics of the Ghezala well, which corresponds to a moderate salt stress, and 3) a T50 treatment, with 0.9 mM KNO3 and 50 mM NaCl, representing the characteristics of the irrigation water of the Oum Hani well, which corresponds to an average salt stress.
Plant material
Seeds of M. sativa (var. Siriver) were sterilized with sodium hypochlorite (2% active chloride) for 20 min, thoroughly rinsed, and imbibed in sterile distilled water for 3 h. Then, germination was carried out at 22°C in Petri dishes, lined with filter papers moistened with distilled water. Germination, identifiable by the emergence of radicles, took place in the dark after 5 days. The germinated seeds were transplanted in plastic pots (8 cm deep) filled with B5 sand (0.6-1.6 mm diameter), provided with adequate drainage device and then grown in culture room at 25°C, with a 16 h/8 h (day/night) photoperiod, and 60/80% (day/night) humidity. Plants were watered every 3 days, once with water, once with a mineral solution containing the following basic macro- and micro-nutrients: 2.5 mM MgSO4, 0.5 mM K2SO4, 2 mM KH2PO, 1 mM CaCl2, 0.05 mM Fe-EDTA, 16 µM H3BO3, 5 µM MnSO4,4H2O, 1 µM ZnSO4,7H2O, 1 µM CuSO4,5H2O, 0.7 µM NaMoO4,2H2O, and 0.5 µM CoCl2,6H2O. In addition to the basic nutrients, either water or mineral solution also contained either 0.2 mM KNO3 (Ctrl0), 0.2 mM KNO3 and 20 mM NaCl (T20), or 0.9 mM KNO3 and 50 mM NaCl (T50). Plants were inoculated 7 days post-imbibition with Ensifer meliloti 2011 (previously named Sinorhizobium meliloti 2011), the predominant microbial symbiotic partner (Frendo et al., 2005).
At 28 days post-imbibition (three-leaf vegetative stage approximately), the plants were either maintained under normoxia (N), or subjected to flooding (H) for 5 days. Flooding treatment was done by submerging pots containing plants to 1 cm above substrate level in the mineral solution supplemented, or not, with NaCl and KNO3 to maintain a medium close to that of the irrigation waters of Ghezala and Oum Hani. At 33 days post-imbibition, the shoots of the plants were harvested, and either measured for biometric parameters or quickly frozen in liquid nitrogen and stored at -80°C until further analysis.
Vegetative growth analysis
The growth and biomass production of shoots was assessed by measuring the following parameters: height, fresh weight (FW), dry weight (DW) and water content, expressed as WC (%) = 100 (FW – DW)/FW, and leaf area (LA), that was measured using the MESURIM free access software: (http://acces.ens-lyon.fr/acces/logiciels/mesurim/telechargement). The dry weight is measured after drying the fresh material for 48 h at 70°C.
Protein measurements
Shoot samples (50 to 150 mg) were ground in a mortar and pestle in a buffer containing 25 mM Tris-HCl, pH 7.0, 0.5 mM β-mercaptoethanol, and 0.3% (p/v) polyvinylpolypyrrolidone. Homogenates were centrifuged for 15 min at 13400 g, and the clarified extracts in the supernatant were recovered. Soluble proteins were quantified on clarified extracts (Bradfor, 1976), using bovine serum albumin as standard.
Determination of chlorophylls
The extraction and determination of the concentration of leaf chlorophyll a, chlorophyll b and total chlorophyll were performed as in Arnon (1949), and calculations were according to the following formula:
Chlorophyll a = 12.7 (A663) - 2.69 (A645),
Chlorophyll b = 22.9 (A645) - 4.68 (A663),
Total chlorophyll = 20.2 (A645) + 8.02 (A663).
Determination of proline
Proline extraction and measurement was carried out according to the method of Bates et al. (1973), on alfalfa shoots previously dried in an oven at 60°C. The optical density was measured at 520 nm. The standard range was established based on pure proline.
Determination of mineral elements
Mineral elements were extracted using the technique of cold nitric extraction from dried plant material according to (Gauquelin et al., 1992). Potassium (K+) and sodium (Na+) were determined by flame emission spectrometry according to Morard and Gullo (1970). The determination of chloride (Cl-) was done by coulometry (chloridometer Buchler-Cotlove) according to Morard and Gullo (1970).
Statistical analysis
Every parameter was analyzed in three independent experiments, and measurements were carried out on samples of five to nine plants per treatment. One-way ANOVA followed by a Tuckey test (http://www.statskingdom.com/index.html) was carried out to determine the effect of the treatments in the analyzed variables. Differences were considered to be significant at the threshold of P < 0.05.
RESULTS AND DISCUSSION
Vegetative growth analysis
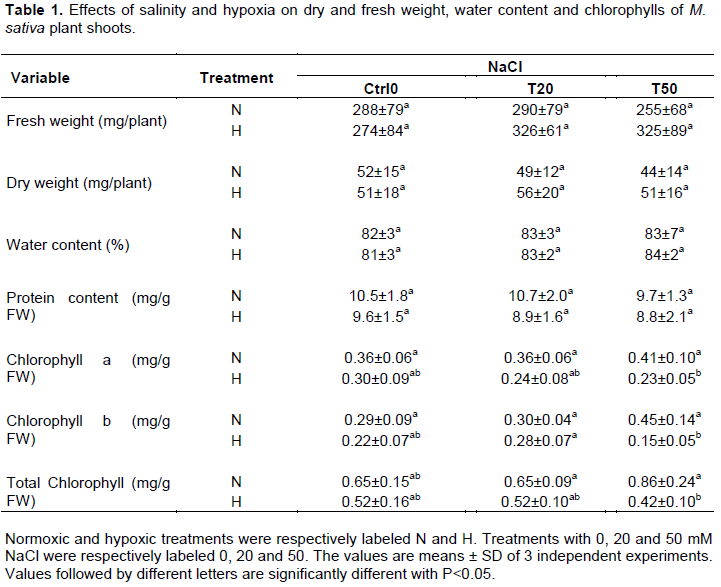
The effects of saline and hypoxic constraints were first analyzed on the growth of M. sativa shoots (Figure 1). As compared to Ctrl 0 in normoxia, after 33 days of growth, the size of the shoots and the leaf area (LA) were not significantly affected by moderate salt treatment (T20), whereas they decreased by 33% and 65%, respectively, in the presence of 50 mM NaCl (T50) (Figure 1A and B). The fresh weight (FW) and dry weight (DW) masses of the shoots were not significantly modified by the salt treatment (Table 1), and the water contents (WC) remained constant (close to 80%) regardless of the NaCl content in the nutrient medium (Table 1). A similar pattern was maintained in shoot dry weight in response to salt stress in 3 month-old M. sativa (var sativa) plants subjected to varying concentrations of NaCl for 4 weeks (Rogers et al., 2008, 2009), which confirms that M. sativa is quite tolerant to salinity. The imposition of 5-day flooding stress did not result in a profound change in growth parameters (Figuure 1 and Table 1), which means that short-term hypoxia did not significantly affect the growth of Medicago sativa in the absence as well as in the presence of NaCl.
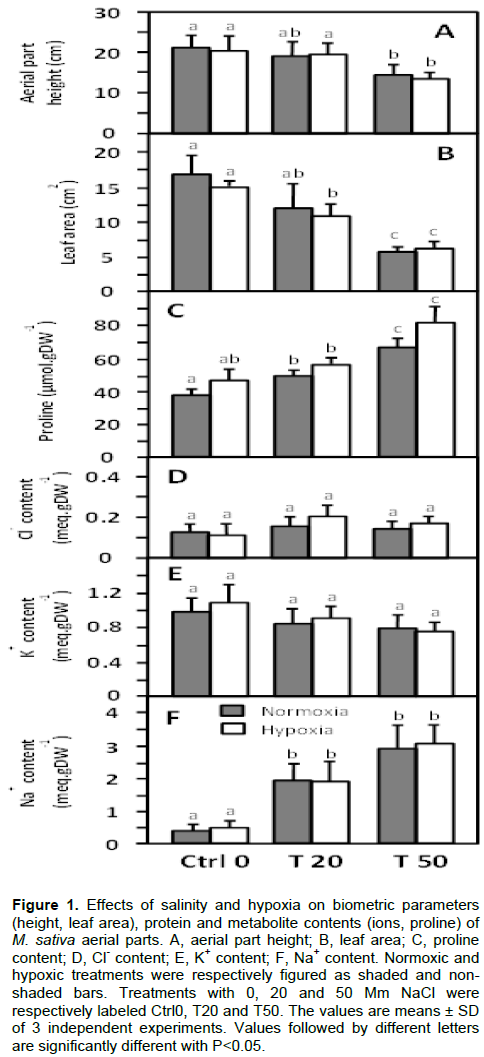
Considered together, these results show that the biomass production of M. sativa was not significantly affected by average salt stress, while the growth of aerial parts and LA already showed a moderate reduction. Such a pattern of maintaining biomass production, while aerial parts and LA are reduced, may be explained by an increase in leaf thickness and mass per area as already observed in M. sativa response to salinity (Smethurst et al., 2008). Interestingly, an increase in leaf mass area was also observed in M. truncatula response to hypoxia (El Msehli et al., 2016), and it may be thought that such response is reinforced in response to combined salinity and hypoxia stresses. Actually, the maintenance of water content revealed that M. sativa has a good ability to adjust its osmotic potential. The absence of additional deleterious effects on the growth parameter, when exposed to the combined treatments of salinity and flooding-induced hypoxia, indicated that, at least over a 5 day-long hypoxia period, M. sativa is able to cope with this double constraint. To go deeper into the physiological study of the effects of single and combined stress, we analyzed parameters related to shoot protein content and photosynthetic capacity on the one hand; and we measured the major ions accumulated under salt constraint (Na+, K+, Cl-), as well as proline which is a salt stress response marker in plants (Tujeta, 2007), on the other hand.
Effect of salinity and hypoxia on protein and chlorophyll contents
The soluble protein content is a valuable indicator to evaluate the impact of stress on the overall functioning of cells and tissues. As shown in Table 1, shoot protein contents were not modified significantly by either saline treatment, or after 5 days of flooding, indicating that M. sativa tolerates the imposition of the combined constraint at least for short periods. Chlorophyll contents were also analyzed to evaluate the capacity of M. sativa shoots to maintain their photosynthesis under combined salinity and hypoxia. Table 1 shows that under normoxia chlorophyll contents (a, b and total) were unaffected by salinity treatments. Similarly, a 5-day long flooding treatment did not change significantly the global chlorophyll contents when compared to the other hypoxically treated plants. But, it should be noted that for plants combining both 50 mM NaCl and flooding treatments (T50, H), their chlorophyll contents were approximately 50% lower than that of 50 mM treated normoxic plants (T50, N). Decreased chlorophyll levels (Table 1), as well as reduced leaf area in plants treated with 50 mM NaCl (Figure 1B), indicate a decline in overall plant photosynthetic capacity. It has been reported that under salt stress, as well as under hypoxia, a close relationship exists between chlorophyll level and photosynthetic capacities (Netto et al., 2005; Striker and Colmer, 2017). Present data indicate, first, that the photosynthetic machinery is affected earlier than is the overall protein content, and second that each stress, considered separately, does not affect the photosynthetic machinery; as already observed in previous studies (El Msehli et al., 2016; Li et al., 2010). However, the combination of the two stresses clearly affects it and impairs the capacity of M. sativa plants to maintain photosynthesis. In this situation, insufficient sugar production cannot cover the needs in maintaining the carbon skeleton for symbiotic nitrogen assimilation and plant growth, and compromises the plant capacity to cope with longer-lasting stress conditions.
Effect of salinity and hypoxia on mineral nutrition
The submission of M. sativa plants to increase salt stress in normoxia was accompanied by a significant increase in the Na+ content in the leaves, from 0.43 meq.g-1 DW in Ctrl0 shoots to 2.0 and 2.95 meq. g-1 DW in T20 and T50 shoots, respectively (Figure 1F). The content of K+, on the order of 1 meq. g-1 DW, did not change significantly as a function of salinity; while the Cl- concentration remained close to 1.6-1.8 meq.vg-1 DW in T50 plants (Figure 1D and E). Such observations are comparable to the results obtained for other saline-treated varieties of M. sativa (Li et al., 2010; Rogers et al., 2008, 2009, 2011). However, in the present study, the Na+/K+ ratios measured in T20 and T50 plant shoots (2.3 and 3.7, respectively) are much higher than those measured in previous reports (Li et al., 2010; Rogers et al., 2009). This is likely due to the fact that in these reports M. sativa plants were first grown in the absence of NaCl for several weeks before being subjected to 2-4 weeks salt stress, whereas in our study plants were subjected to salinity from germination, as is the case in the field, and they have developed from the beginning a system of salt storage in the aerial parts. It also shows that M. sativa can store high amounts of NaCl in the leaves without significantly affecting plant growth. In comparison with normoxia treated plants, the application of flooding stress did not change the Na+, K+ and Cl- contents in the shoots (Figure 1D, E and F), which means that ion transport and metabolism is not significantly altered by short-term flooding.
Effect of salinity and hypoxia on proline accumulation
As shown in Figure 1C, compared to normoxic treatment (Ctrl0), exposure to increasing concentrations of NaCl resulted in a significant and expected increase in leaf proline contents of 29 and 79%, in T20 and T50 shoots, respectively. Proline is a general marker of salt stress response in most plants (Mansour and Ali. 2017; Per et al., 2017), including legumes (Bruning and Rozema, 2013). Proline biosynthesis was expected to occur as a consequence of disturbance in cell homeostasis and to reflect damage in response to salt stress (Khalil et al.,2016). Either an external supply (Ehsanpour and Fatahian, 2003), or an increased cellular level (Campanelli et al., 2013) in proline, promotes salt tolerance in Medicago species. Moreover, salt tolerance was found to be enhanced in Medicago truncatula plants transformed with a gene for proline synthesis (de la Pena et al., 2010). As compared with normoxia-treated plants, the application of hypoxia led to a slight increase (8-24%) in proline contents after 5 days of treatment. Although the difference was not significant between N-50 and H-50 plants, such an increase is of interest, because proline accumulation has recently been observed in Phaseolus vulgaris response to hypoxia (Velasco et al., 2019). It was suggested to serve as a source of energy, nitrogen or carbon to reinitiate growth after the hypoxia stress period (Aloni and Rosenshtein, 1982). However, further study would deserve investigation to confirm this observation and to analyze the potential role of proline in the combined salinity-hypoxia stress response.
CONCLUSION
M. sativa is a legume crop known to be tolerant to salinity (Bruning and Rozema, 2013; Zahran, 1999) and drought stress (Bouton, 2012), and sensitive to waterlogging and hypoxia (Rogers, 1974; Striker and Colmer, 2017). In northern Africa, M. sativa is often grown in areas where, during the rainy season or after irrigation turn, cultivated fields are subject to the double stress of salinity and flooding. Our results showed that when M. sativa is cultured on moderate (20 mM) and medium (50 mM) NaCl concentrations, the classical mechanisms of stress response (mineral ion accumulation, proline synthesis) are set up and the growth (FW, DW, growth of aerial parts) is only moderately affected by salt. At low NaCl concentration, short-term hypoxia did not significantly affect growth. But, in a situation of average salt stress (that is, 50 mM NaCl), the combined stresses seemed to lead, starting from 5 days of hypoxia, to an early alteration of the photosynthetic machinery and a modification of the compartmentalization of ions between roots and leaves. A study of the combined effects of salt and hypoxia during a longer period of hypoxia is needed to evaluate more deeply the impact of combined stresses on growth and yield of M. sativa. However, as a first step, it is recommended to control crop irrigation regimes to avoid too long period of flooding and the risk to significantly impact final yields.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Aloni B, Rosenshtein G (1982). Effect of flooding on tomato cultivars: The relationship between proline accumulation and other morphological and physiological changes. Physiologia Plantarum 56:513-517. |
|
|
Armstrong W (1980). Aeration in higher plants. In: Woolhouse HW (ed) Advances in Botanical Research 7:225-332. |
|
|
Arnon D (1949). Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiology 24(1):1-15. |
|
|
Bahri A (1993). Evolution de la salinité dans un périmètre irrigué de la Basse Vallée de la Medjerda en Tunisie. Science du Sol 31(3):125-140. |
|
|
Bates LS, Waldren RP, Teare ID (1973). Rapid determination of free proline for water-stress studies. Plant and Soil 39:205-207. |
|
|
Bouton JH (2012). An overview of the role of lucerne (Medicago sativa L.) in pastoral agriculture. Crop and Pasture Science 63:734-738. |
|
|
Bradford M (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry 72:248-254. |
|
|
Bruning B, Rozema J (2013). Symbiotic nitrogen fixation in legumes: Perspectives for saline agriculture. Environmental and Experimental Botany 92:134-143. |
|
|
Campanelli A, Ruta C, Morome-Fortunato I, De Mastro G (2013). Alfalfa (Medicago sativa L.) clones tolerant to salt stress: in vitro selection. Central European Journal of Biology 8(8):765-776. |
|
|
de la Pena TC, Redondo FJ, Manrique E, Lucas MM, Puyeo JJ (2010). Nitrogen fixation persists under conditions of salt stress in transgenic Medicago truncatula plants expressing a cyanobacterial flavodoxin. Plant Biotechnology Journal 8:954-965. |
|
|
El Msehli S, Rima N, Sghaier H, Aschi-Smiti S, Brouquisse R (2016). Impact of hypoxia on nodulation and growth of the legume plant Medicago truncatula in symbiosis with Sinorhizobium meliloti. International Journal of Current Research in Biosciences and Plant Biology 3(11):53-60. |
|
|
Ehsanpour AA, Fatahian N (2003). Effects of salt and proline on Medicago sativa callus. Plant Cell, Tissue and Organ Culture 73:53-56. |
|
|
Frendo P, Harrison J, Norman C, Hernandez-Jiménez MJ, Van De Sype G, Gilabert A, Puppo A (2005). Glutathione and homoglutathione play a critical role in the nodulation process of Medicago truncatula. Molecular Plant-Microbe Interactions 18(3):254-259. |
|
|
Gauquelin T, Fromard F, Badri W, Dagnac J (1992). Apports d'éléments minéraux au sol par l'intermédiaire de la litière, des pluies et des pluviolessivats dans un peuplement à genévrier thurifère (Juniperus thurifera L) du Haut Atlas occidental (Maroc). Annales des Sciences Forestières 49(6):599-614. |
|
|
Glenn EP, Brown JJ, Blumwald E (1999). Salt Tolerance and Crop Potential of Halophytes. Critical Reviews in Plant Sciences 18(2):227-255. |
|
|
Khalil C, Haddioui A, El Hansali M, Boufous AH (2016). Growth and proline content in NaCl stressed plants of annual medic species. International Journal of Advanced Research in Biological Sciences 3(9):82-90. |
|
|
Li R, Shi F, Fukuda K, Yang Y (2010). Effects of salt and alkali stresses on germination, growth, photosynthesis and ion accumulation in alfalfa (Medicago sativa L.). Soil Science and Plant Nutrition 56(5):725-733. |
|
|
Mansour MM, Ali EF (2017). Evaluation of proline functions in saline conditions. Phytochemistry 140:52-68. |
|
|
Morard P, Gullo JH (1970). Minéralisation de tissus végétaux en vue du dosage de P, K, Ca, Mg et Na. Annales d'Agronomie 21(2):229-236. |
|
|
Netto AT, Campostrini E, de Oliveira GJ, Bressan-Smith RE (2005). Photosynthetic pigments, nitrogen, chlorophyll a fluorescence and SPAD-502 readings in coffee leaves. Scientia Horticulturae 104(2):199-209. |
|
|
Per TS, Khan NA, Reddy PS, Masood A, Hasanuzzaman M, Khan MIR, Anjum NA (2017). Approaches in modulating proline metabolism in plants for salt and drought stress tolerance: Phytohormones, mineral nutrients and transgenics. Plant Physiology and Biochemistry 115:126-140. |
|
|
Rogers VE (1974). The response of lucerne cultivars to levels of waterlogging. Australian Journal of Experimental Agriculture and Animal Husbandry 14(69):520-525. |
|
|
Rogers ME, Colmer TD, Frost K, Henry D, Cornwall D, Hulm E, Deretic J, Hughes SR, Craig AD (2008). Diversity in the genus Melilotus for tolerance to salinity and waterlogging. Plant Soil 304: 89-101. |
|
|
Rogers ME, Colmer TD, Frost K, Henry D, Cornwall D, Hulm E, Hughes S, Nichols PGH, Craig AD (2009). The influence of NaCl salinity and hypoxia on aspects of growth in Trifolium species. Crop and Pasture Science 60:71-82. |
|
|
Rogers ME, Colmer TD, Nichols PGH, Hughes SJ, Frost K, Cornwall D, Chandra S, Miller SM, Craig AD (2011). Salinity and waterlogging tolerance amongst accessions of messina (Melilotus siculus). Crop and Pasture Science 62:225-235. |
|
|
Smethurst CF, Garnett T, Shabala S (2005). Nutritional and chlorophyll fluorescence responses of lucerne (Medicago sativa) to waterlogging and subsequent recovery. Plant and Soil 270:31-45. |
|
|
Smethurst CF, Rix K, Garnett T, Auricht G, Bayart A, Lane P, Wilson SJ, Shabala S (2008). Multiple traits associated with salt tolerance in lucerne: revealing the underlying cellular mechanisms. Functional Plant Biology 35(7):640-650. |
|
|
Striker GG, Colmer TD (2017). Flooding tolerance of forage legumes. Journal of Experimental Botany 68(8):1851-1872. |
|
|
Tujeta N (2007). Mechanisms of high salinity tolerance in plants. Methods in Enzymology 428:419-438. |
|
|
Velasco NF, Ligarreto GA, Díaz HR, Fonseca LPM (2019). Photosynthetic responses and tolerance to root-zone hypoxia stress of five bean cultivars (Phaseolus vulgaris L.). South African Journal of Botany 123:200-207. |
|
|
Zahran HH (1999). Rhizobium-Legume Symbiosis and Nitrogen Fixation under Severe Conditions and in an Arid Climate. Microbiology and Molecular Biology Reviews 63(4):968-989. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


