ABSTRACT
Baobab (Adansonia digitata L.) is considered an under-utilized species even though it is an economically useful tree often used daily in the diet of rural communities in West Africa. Baobab seeds do not germinate immediately after they are released from ripe fruits due to dormancy imposed by hard seed coats. A study was carried out to assess (1) best soaking duration for seed pretreatment in concentrated sulfuric acid (96%) to increase germination rate of baobab seeds; and (2) relationships between seed morphometric traits as varying with provenances and germination rate. Length, width, thickness and weight of baobab seeds collected from seven provenances spread across an agro-climatic gradient of 250 to 1100 mm in Senegal were first measured. Then, seeds were pretreated by soaking them in distilled water (control) and concentrated H2SO4 (96%) for 12 min, 30 min, 1, 3, 6, 8 and 12 h, respectively. Pretreated seeds were placed on moistened filter paper in 16 cm diameter Petri dishes in an incubator at 25°C for germination. Highest germination rates were obtained for seeds that had been soaked for 6, 8, and 12 h in concentrated sulfuric acid as evidenced by germination rates at 3, 7 and 10 days after sowing, respectively, although we did not find any significant differences among these three soaking durations. Significant variation among provenances was observed in seed morphology traits and seed germination rates. Highest seed trait and germination rate values were observed for Sudano-Sahel and Sudan agro-ecological provenances. This study has shown a significant variation in these characters mainly related to geographical origin. Overall, results could be useful for a further domestication and integration of baobab in the agro-productive system in Senegal as knowledge on seed germination requirements is a critical factor in seedling production for subsequent planting and reforestation.
Key words: Agro-ecology, baobab seed, dormancy, germination physiology, germination rate, non-timber forest product (NTFP), under-utilized species.
Baobab (Adansonia digitata L.) is an African fruit tree species belonging to Malvaceae family. All eight species within genus Adansonia are distinctive trees growing in the tropics. A. digitata grows throughout the drier parts of Africa. A second species (A. gibbosa [A. Cunn.] D. Baum) is restricted to northwest Australia, whereas the remaining six species are endemic to Madagascar (Randall et al., 1998).
A. digitata is considered an under-utilized species even though it is a key economic species often used daily in the diet of numerous rural communities in West Africa (Codjia et al., 2001; De Caluwé et al., 2009; Assogbadjo et al., 2010) particularly in Senegal. Baobab has numerous medicinal and non-medicinal uses. Every part of the tree (bark, leaves, fruits, wood, roots, seeds, sap, etc.) is reported to be useful (Owen, 1970; Sidibé and Williams, 2002; Codjia et al., 2003).
For any specific species, germination responses vary according to site and environmental factors such as latitude, elevation, soil moisture and nutrient content, temperature, kind and density of plant cover and degree of habitat disturbance of the site where the seed matures (Ginwal et al., 2005; Assogbadjo et al., 2005b, 2006). Knowledge of seed germination dynamics is an important tool to increase reproduction success. In general, under adequate aeration and temperature conditions, non-dormant plant seeds need only access to water to be able to germinate after putting them in the soil. But even in such optimal conditions, baobab seeds often remain several weeks in the soil before germinating (Gebauer et al., 2002; ICUC, 2006, 2010).
Seeds may be dormant for a variety of reasons, including (1) seed immaturity at fruit harvest; (2) low seed coat permeability to water and/or oxygen; (3) seed coat resistance to embryo growth; (4) presence of metabolic blocks in the embryo; and (5) various combinations of the afore-mentioned (Kozlowski, 2002). According to Finch-Savage and Leubner-Metzger (2006), seed dormancy is an innate seed property that defines the environmental conditions in which the seed is able to germinate. Sometimes, viable seeds which have no primary dormancy lapse into a state of secondary dormancy as an adaptation when exposed to unfavorable environmental regimes (Mayer and Poljakoff-Mayber, 1989).
In natural conditions, baobab seedlings do not emerge immediately after seeds are released from ripe fruits due to a physical dormancy imposed by hard seed coats which appear to be non-permeable (Esenowo, 1991; Sidibé and Williams, 2002). In natural conditions, baobab seed dormancy is broken by a passage through the digestive system of arge mammals (Gebauer et al., 2002). In nursery conditions, dormancy may be broken by immersing seeds in hot water for several minutes or chopping the seed coat (Esenowo, 1991). According to Danthu et al. (1995), acid scarification for 6 to 12 h is the optimal pretreatment method for breaking baobab seed coat inhibition. The conditions necessary for seeds to “break” dormancy and germinate can be highly variable among species, within a species, or among seed sources of the same species (Falemara et al., 2014).
Based on the annual rainfall gradient and local agricultural practices, there are four different agro-climatic zones in Senegal (Salack et al., 2011) where A. digitata occurs. A. digitata is slightly more represented of the humid zones (Sudano-Sahel and Sudan zones) particularly in areas with alkaline soils. Its morphology (fruits, leaves and stem) differs according to habitats (Diaité, 2005). Fagg and Barnes (1990) said that the tree exhibits considerable phenotypic variability across its diverse habitats, which could be indicative of its high genetic variability. Considering the vast distribution of wild A. digitata over different parts of Senegal, it is reasonable to expect genetic differentiation among A. digitata L. populations in a number of traits which could be exploited through selection of superior populations for seed collection and domestication needs. In addition, baobab is known to be mammal-pollinated (visited by bats) (Sidibé and Williams, 2002), which probably reduces gene flow and thus enhances the genetic differentiation between populations. In Senegal, there is insufficient knowledge about provenance and genetic variability of important indigenous species in general and A. digitata L. in particular. Genetic variation among and within baobab (A. digitata L.) provenances in seed germination and seedling growth has been reported in selected natural populations in Malawi (Munthali et al., 2012). Assogbadjo (2006) has reported variability in seed morphology traits and germination of different baobab provenances in Benin. Several studies have found genotypic variation in the physiology and morphology of tree species which can often be related to the habitat from which the plants originate. The genetic component of this variation among populations from different regions can, therefore, be identified by provenance testing (seed morphometric traits and germination studies).
For reforestation and enhancement of A. digitata seedling production, there is a need to understand the basic requirements for promotion of baobab seed germination in general and more specifically from different provenances. Therefore, we tested first how dormancy could be broken by applying concentrated sulfuric acid (H2SO4) to different Senegalese baobab seed accessions in order to increase their germination rates and reduce germination duration. In this paper, we also report results on variations in seed morphometric characteristics and seed germination related parameters among seven provenances of A. digitata and their mutual relationships with climatic and environmental factors such as temperature, altitude, and humidity.
Seed collection and sampling procedures
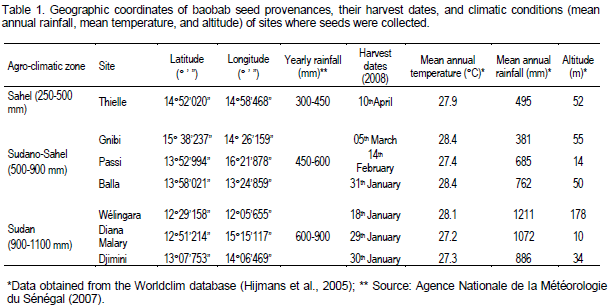
A. digitata seeds were obtained from trees collected during the dry season between January and April 2008 in three agro-climatic zones (Sahel: 250-500 mm; Sudano-Sahel: 500-900 mm; Sudan: 900-1100 mm) of Senegal (Salack et al., 2011) where baobab (A. digitata) is mostly represented (Table 1). Based on provenance sites, we extracted mean annual precipitation, mean annual temperature and altitudinal data using GIS maps of climatic variables of the Worldclim climate and altitude database (Hijmans et al., 2005). Worldclim maps represent interpolated surfaces at a spatial resolution of 30 arcseconds (~1 km) and are based on 50-year means calculated for the 1950 to 2000 period (De Smedt et al., 2011).
Sampling sites were situated along a decreasing rainfall gradient in order to reflect a climatic continuum. In each agro-climatic zone, three populations of baobab were randomly sampled (minimum distance between populations: 50 km). In each population, a random collection of trees separated by a minimum of 100 m was sampled with the distance chosen to reduce the probability of consanguinity between trees (Dawson et al., 2009). From 40 trees per population, 30 mature indehiscent fruits per tree were collected over the canopy. Each baobab population was geo-referenced with a GPS. Trees were marked with a metal tag to allow follow up measurements in subsequent years (on the tag: Population number-individual number).
In the laboratory, fruits were broken and seeds separated from pulp and fibers. Seeds extracted were soaked in distilled water for 6 HR (Sacande et al., 2006). Dead, empty or immature seeds float on the water while a fully healthy seed sinks (Sacande et al., 2006). Healthy seeds were collected and subsequently rinsed and dried under ambient temperatures for 2 days before storing in a cold chamber at 15°C.
Morphometric seed characterization
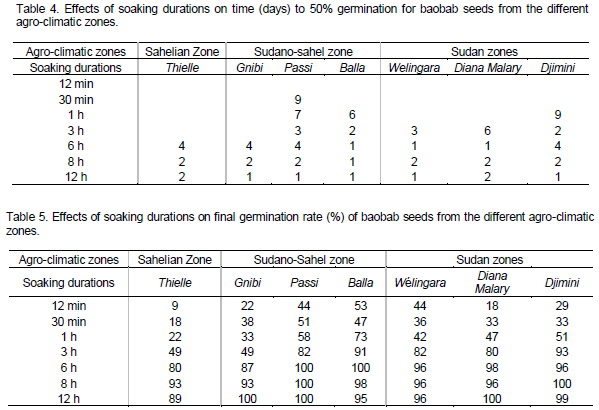
As seed germination rate may vary in relation to morphological seed traits (Loha et al., 2006), the following three morphometric parameters were measured on three lots of 20 seeds for each provenance, using a digital caliper (Titan 23175±0.02 mm): seed length, seed width and seed thickness. Three replicates of hundred (100) seeds of each provenance were weighed using an electronic balance (maximum = 400 g, d=0.01 g model Scout Pro Spu 402 s/n 7125050266).
Germination rate and germination parameters
The number of germinated seeds was counted every day to evaluate germination rates until seeds germination was over in each treatment. Germination parameters as defined by Evenari (1957) and Côme (1968) were calculated. Germination rate was calculated at 3, 7 and 10 days after sowing (DAS). Germination parameters monitored were germination speed (number of days to 50% of seeds germinated) and final germination rate (Gr) defined as the total number of seeds from the first to the last germinated seed. A seed was considered as germinated when the first radical pierced its integument (Redondo-Gómez et al., 2007).
Data analysis
At the end of the experiment, germination counts were calculated in percentages (%) and data subjected to an analysis of variance (ANOVA)) with Statistix version 8.1. Mean separation was performed using Tukey High Significant Difference (HSD) test at P < 0.05.
Morphological seed traits
Morphological seed trait values followed a wet to dry pattern with provenances from humid zones (Sudano-Sahel and Sudan zones) having the higher values. There were highly significant differences (P<0.05) among baobab provenances with regard to seed l ength, seed thickness and seed weight (Table 2). The average seed length, seed thickness, seed width and 100-seed weight are shown in Table 2. Overall, provenances from humid zones showed highest seed length, thickness and weight while one provenance (Balla) of the intermediate agro-climatic Sudano-Sahel zone had the lowest values for these morphological seed traits. High variation was observed in 100-seed weight for which almost all provenances differed significantly (P<0.001) among each other. Djimini and Diana Malary provenances both from the Sudan zone had the highest seed weight (50.59 and 44.99 g per 100 seeds, respectively), as compared to Balla which had the smallest value (34.19 g).
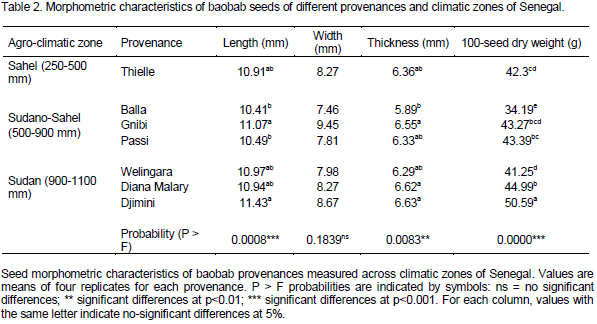
Seed germination rates associated with H2SO4 (96%) treatment
Baobab seeds from the seven provenances were subjected to concentrated sulfuric acid (96%) treatment in different soaking durations for increasing germination rate. Our results show highly significant differences among treatments. Seeds soaked in concentrated sulfuric acid for 12 h gave the best germination rates with 70, 96 and 97% of seeds germinated at 3, 7 and 10 DAS, respectively (Table 3). However, at 7 and 10 DAS, no significant differences were found between 6, 8 and 12 h soaking durations, even though germination rates increased with longer soaking durations for all treatments (Table 3). No germination was recorded for any seeds in the control treatments (To) for the duration of the experiment. Similar trends as for morphological seed traits were noticed in seed germination rates among the different provenances whereby highest values at 3, 7 and 10 DAS were recorded for provenances in the humid zone, particularly Balla and Passy from Sudano-Sahel zone which is characterized by small and intermediate seed sizes (Tables 2 and 3). Seed germination rate seems to be increased with increasing rainfall gradient.
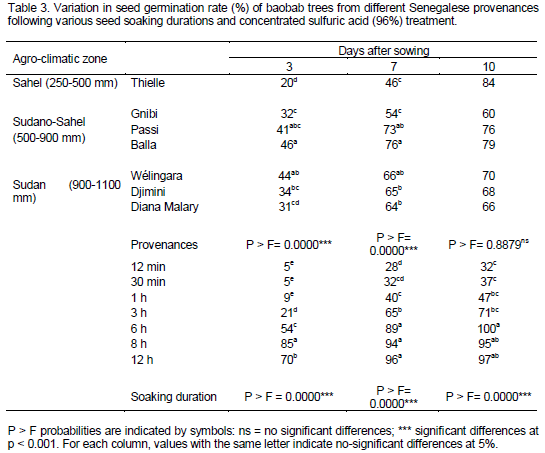
For each provenance, results on germination parameters (Tables 4 and 5) show that soaking durations shorter than 30 min do not allow 50% of seeds to germinate, whereas 6 to 12 h soaking durations take 4 or less than 2 day to reach 50% of seed germination (Table 4). Baobab seeds of Thielle and Gnibi provenances from respectively Sahel and Sudano-Sahel zones never reach 50% of germination when they were soaked in concentrated sulfuric acid between 12 min and 3 h.
Furthermore, for each provenance, final seed percentage germination was very high when seeds were soaked for 6 to 12 h in concentrated sulfuric acid (96%), whereas the highest value was obtained for humid zone material (Sudan and Sudano-Sahel zones) particularly Balla and Passy from the Sudano-Sahel zone, which is characterized, respectively, by small and medium seed size (Tables 2 and 5).
Relationships of seed traits with climatic parameters and final germination rate of the baobab provenances
A close relationship of seed morphometric traits was observed with final germination rate of baobab provenances and climatic parameters. Seed size affects its ability to germinate, with smaller seed sizes (low values of seed length, thickness, and/or weight) yielding higher germination rates. The Principal Component Analysis (PCA) explains 71.89% of data variation in the two first dimensions (Figures 1 and 2). Dimension 1 (50.58%) indicates a tendency of high morphological seeds traits (length, width, thickness, 100-seed dry weight) and weak final germination rate. Dimension 2 seems to be the component presenting climate parameters. It was negatively associated with yearly rainfall and positively correlated with mean annual temperature and altitude. As shown in Figures 1 and 2, PCA analysis revealed four groups of provenances: Djimini and Diana Malary accessions, with high rainfall, had larger and heavier seed sizes and consequently weak final germination rate. Balla from the Sudano-Sahel zone with small seed size and average rainfall had high final germination rate. Gnibi had heavier seed sizes with relatively high rainfall but low final germination rate. Passi, Thielle and Wélingara provenances (from respectively the Sudano-Sahel, Sahel and the Sudan zone), with intermediate seed sizes have average final germination rates.
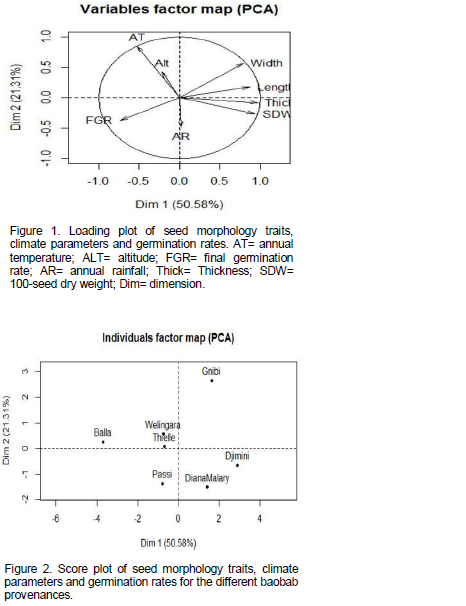
The baobab (A. digitata L.) provenances exhibited considerable amount of variation in seed morphometric traits. Analysis of variance of the data on seed weight and seed size showed that the difference between baobab provenances were significant (p<0.05). The variation found in seed morphometric traits of the humid zones (Soudano-Sahel and Sudan zone) tended to follow provenance variation from the Sudan zone (high rainfall), particularly seeds from Djimini which had the heaviest seeds. Since seed weight and size are adaptive traits to varying edapho-climatic environments (Raddad, 2007), their smaller mean values in the Balla population might suggest a strong genetic selection for small seed size as an adaptation to the greater desiccation stress in this rather arid zone (Dangasuk et al., 1997; Assogbadjo et al., 2005b). Size and 100-seed weight in Thielle (Sahel zone) were not significantly different from those measured in both humid zones (Sudan and Sudano-Sahel zones). Baobab trees in Thielle are located in an edaphic environment where clayey soils help maintain soil water retention.
Variation in A. digitata provenances with respect to their seed morphometric traits could be due to the fact that these species grow over a wide range of climatic conditions. Several authors found a close relationship between environment (particularly rainfall) and seed weight, the latter reveals that plants growing in areas with lower rainfall tend to develop both lower fruit and seed weights (Raddad, 2007; De Smedt et al., 2011). High inter- and intra-population variation in seed weight which might influence seed germination might suggest that this parameter is quite the most sensitive to environmental factors (Marteen et al., 2002; Assogbadjo et al., 2005a) and could probably contribute to explain the relative distribution pattern of A. digitata over the different agro-climatological zones. In Senegal, baobab populations are slightly more represented in the humid areas. Since the seeds were collected from different locations and from trees approximately of the same age, differences observed in seed parameters, therefore, may be attributed to genetic in nature as a result of adaptation to diverse environmental condition prevailing throughout their distributional range (Mathur et al., 1984). Apart from age, vigour, crown exposure and genotype of mother tree, soil and climate of the place of seed origin are important factors affecting the seed traits (Salazar and Quesada, 1987).
To improve their germination performance, baobab seeds require physical or chemical pretreatment before they can germinate (Sidibé and Williams, 2002). According to Levitt (1974), immersion of seeds in high H2SO4 concentration disrupts the seed coat. Many researchers use pretreatment with H2SO4 with soaking durations differing between 30 min and 1 h (Sacande et al., 2006) and 3 h (Razanameharizaka et al., 2006) to promote baobab seed germination. Our results show highly significant differences among baobab provenances in germination whereas seeds soaked in concentrated sulfuric acid for 6 to 12 h gave the best germination rates in our own experiment. This leads to germination rates of more than 90% (Danthu et al., 1995). In Mali, the World Agroforestry Center (ICRAF) used sulfuric acid for 90 min followed by rinsing in water for 24 h and obtained 92% germination for baobab seeds. Ibrahim and Otegbeye (2004) reported that soaking baobab seeds in a boiling solution of 4 ml of black potash/500 ml for 1 h consistently gave the best germination results, with 92.67% at 45 days after sowing (DAS). The same authors showed that the lowest germination result (4.33% at 45 DAS) was recorded for baobab seeds soaked in cold water for one hour and seeds soaked in boiling water for 24 h. However, Falemara et al. (2014) findings showed that seeds subjected to 98% acid concentration for 6 and 12 h gave a low germination percentage compared to average percentage germination on exposure to 10 and 50% acid concentrations at varying time intervals.
Our results evidence that provenances in the humid zone, particularly Balla and Passy from Sudano-Sahel zone which is characterized respectively by small and intermediate seed size gave the best performance in germination. Assogbadjo (2006) found that germination of freshly harvested baobab seed can start between 5 and 7 days after sowing without applying any scarification. He also reported that highest germination rates were recorded for more humid zone provenances, which is in agreement with our own results. In other studies, it was shown that germination of seeds was directly related to seed size and depth at which seeds had been buried upon planting (Bond et al., 1999; Ren et al., 2002). Small seeds have a better chance to enter into the soil than large seeds, and thus facilitate the build-up of a persistent soil seed bank, crucial for regeneration of the species (Ekta and Singh, 2000). However, the same authors reported that due to the availability of a large source of reserve material, larger seeds show higher germination percentages, greater seedling survival and better growth. Also, according to Milberg and Lamont (1997), large and heavy seeds contain larger amounts of reserves that promote germination. Subsequently, high seed reserves may enhance the abilities of larger and heavier seeds to persist by providing for metabolic requirements during the latent period, until suitable light or moisture conditions occur.
All baobab seeds from the various provenances yielded high final germination rates and rapidly reaching 50% of seed germination (1 to 4 days) when seeds were soaked in concentrated sulfuric acid for 6 to 12 h. Germination speed gives an idea of the vigour of the seed and of the seedling, which it produces (Willan, 1985). The interest in germination speed is based on the theory that only those seeds which germinate rapidly and vigorously under favourable conditions are likely to produce vigorous seedlings in field conditions, whereas weak or delayed germination is often fatal (Aldhous, 1972). Therefore, concentrated sulfuric acid (96%) for 6 to 12 h can be used to enhance baobab seed germination.
Results on correlations between parameters show that seed size affects its ability to germinate, with smaller seed sizes (low values of seed length, thickness, and/or weight) yielding higher germination rates. Climate parameters (particularly yearly rainfall) seemed to have a firm relation with our baobab morphological seed traits and germination rate. Close relationship within species between seed size and/or weight and percentage germination have been documented for many tropical tree seedlings (Ekta and Singh, 2000). These results are similar to those of Loha et al. (2006) who investigated on provenance variation in seed morphometric traits and germination of Cordia africana Lam. in Ethiopia.
The present study suggests that concentrated sulfuric acid (96%) disrupts the seed coat and facilitates rapid germination. We have shown that seed pretreatment with concentrated sulfuric acid (96%) significantly reduced time to full germination and increased percentage of germinated seeds for baobab seeds of all provenances tested. Seed germination increased with increasing soaking durations while the highest germination rates were recorded for provenances of the more humid zones (Sudano-Sahel and Sudan) and were obtained when baobab seeds were soaked in concentrated sulfuric acid for 6, 8 and 12 h. Results show that baobab germination can be stimulated by using concentrated sulfuric acid. However, optimum germination rate could also depend on seed size which might be influenced by environmental factors.
The present study revealed that considerable variation exists in baobab provenances with respect to seed morphology and seed germination characteristics. Based on our results, eventually, it may be concluded that baobab provenance with small seed size, rapid and high germination rates from Sudano-Sahel zone (Balla) seem to be easier to germinate and therefore could be used as a basis source material for domestication in Senegal. However, more efforts toward improving the germination rate of seeds from other provenances particularly Gnibi and Thielle need to be pursued.
In this study, we also noticed that the average values of morphological seed traits and germination rates are not always significantly different from one climate zone to another, even though climatic parameters, particularly yearly rainfall, that determine seed size were correlated with morphological seed traits and with germination rates. This may contribute to open a field of research to identify genotypic differences between the different baobab provenances that could also explain differences in morphometric seed characteristics and germination behavior which are generally genetically controlled.
The authors have not declared any conflict of interest.
This study was conducted in CERAAS, Senegal with the collaboration of LATSE (Laboratory of Tropical and Subtropical Agriculture and Ethnobotany) within the context of DADOBAT (Domestication And Development of Baobab and Tamarind) project financed by the European Union (EU) (project no. 032217). The authors are grateful to CERAAS staff, particularly technicians, and to the farmers for their help in field collection of baobab fruits. We would also like to thank Agricultural Engineer, Emile Codjo Agbangba for his help in data analysis.
REFERENCES
|
Aldhous JR (1972). Nursery practice. Forestry Commonwealth Bulletin, London. 45:16-17. |
|
|
|
Assogbadjo AE, Kakai RG, Kyndt T, Sinsin B (2010). Conservation Genetics of Baobab (Adansonia digitata L.) in the Parklands Agroforestry Systems of Benin (West Africa). Not. Bot. Hort. Agrobot. Cluj. 38:136-140. |
|
|
Assogbadjo AE, Kyndt T, Sinsin B, Gheysen G, Van Damme P (2006). Patterns of Genetic and Morphometric Diversity in Baobab (Adansonia digitata L.) Populations Across Different Climatic Zones in Benin (West Africa). Ann. Bot. 97:819-830.
Crossref |
|
|
|
Assogbadjo AE, Sinsin B, Codjia JTC, Van Damme P (2005a). Ecological diversity and pulp, seed and kernel production of the baobab (Adansonia digitata L.) in Benin. Belg. J. Bot. 138:47-56. |
|
|
Assogbadjo AE, Sinsin B, Van Damme P (2005b). Caractères morphologiques et production des capsules de baobab (Adansonia digitata L.) au Bénin. Fruits 60:327-340.
Crossref |
|
|
Bond WJ, Honig M, Maze KE (1999). Seed size and seedling emergence: an allometric relationship and some ecological implications. Oecologia 120:132-136.
Crossref |
|
|
|
Codjia JTC, Assogbadjo AE, Ekue MRM (2003). Diversité et valorisation au niveau local des ressources forestières alimentaires végétales du Bénin. Cah. Agric. 12:321-331. |
|
|
|
Codjia JTC, Fonton-Kiki B, Assogbadjo AE Ekue MRM (2001). Le baobab (Adansonia digitata L.), une espèce à usage multiple au Bénin. Cotonou, Bénin: Coco Multimédia. |
|
|
|
Côme D (1968). Problèmes de terminologie posés par la germination et ses obstacles. Bulletin Société Française. Physiol. Veg. 14:3-9. |
|
|
Dangasuk OG, Seurei P, Gudu S (1997). Genetic variation in seed and seedling traits in 12 African provenances of Faidherbia albida (Del.) A. Chev. at Lodwar, Kenya. Agroforest. Syst. 37:133-141.
Crossref |
|
|
|
Danthu P, Roussel J, Gaye A, El Mazzoudi EH (1995). Baobab (Adansonia digitata L.) seed pre-treatments for germination. Seed. Sci. Technol. 23:469-475. |
|
|
Dawson I, Lengkeek A, Weber J, Jamnadass R (2009). Managing genetic variation in tropical trees: linking knowledge with action in agroforestry ecosystems for improved conservation and enhanced livelihoods. Biodivers. Conserv. 18:969–986.
Crossref |
|
|
De Caluwé E, De Smedt S, Assogbadjo AE Samson R, Sinsin B, Van Damme P (2009). Ethnic differences in use value and use patterns of baobab (Adansonia digitata L.) in northern Benin. Afr. J. Ecol. 47:433-440.
Crossref |
|
|
De Smedt S, Alaerts K, Kouyate AM, Van Damme P, Potters G, Samson R (2011). Phenotypic variation of baobab (Adansonia digitata L.) fruit traits in Mali. Agroforest. Syst. 82:87-97.
Crossref |
|
|
|
Diaité I (2005). Agroforesterie et sécurité alimentaire au Sénégal, Journée Biodiversité, IRD Hann, Dakar, Sénégal P. 24. |
|
|
|
Ekta K, Singh JS (2000). Ecology of seed and seedling growth for conservation and restoration of tropical dry forest: a review. Environ. Conserv. 28:39-52. |
|
|
Esenowo GJ (1991). Studies on germination of Adansonia digitata L. seeds. J. Agrc. Sci. 117:81-84.
Crossref |
|
|
|
Evenari M (1957). Les problèmes physiologiques de la germination. Bulletin Société Française. Physiol. Veg. 3:105-124. |
|
|
|
Fagg CW, Barnes RD (1990). African acacias: study and acquisition of the genetic resources. Final Report on Overseas Development Administration Research Scheme R. 4348 OFI, UK. P. 170. |
|
|
|
Falemara BC, Chomini MS, Thlama DM, Udenkwere M (2014). Pre-Germination and Dormancy Response of Adansonia digitata L. Seeds To Pre-treatment techniques and growth media. Eur. J. Agric. For. Res. 2(1):31-41. |
|
|
Finch-Savage WE, Leubner-Metzger G (2006). Seed dormancy and the control of germination. New Phytol. 171:501-523.
Crossref |
|
|
|
Gebauer J, El Siddig K, Ebert G (2002). Baobab (Adansonia digitata L.): a review on a multipurpose tree with promising future in the Sudan. Gartenbauwis senschaf. 67:155-160. |
|
|
|
Ginwal HS, Phartyal SS, Rawat PS, Srivastava RL (2005). Seed source variation in morphology, germination and seedling growth of J. curcas Linn. in Central India. Silvae Genet. 54:76-80. |
|
|
|
Ginwal HS, Rawat PS, Srivastava RL (2004). Seed source variation in growth performance and oil yield of Jatropha curcas Linn. Silvae. Genet. 53:186-192. |
|
|
Hijmans RJ, Cameron SE, Parra JL, Jones PG, Jarvis A (2005). Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 25:1965-1978.
Crossref |
|
|
|
Ibrahim A, Otegbeye G (2004). Methods of achieving optimum germination of Adansonia digitata L. Bow. J. Agric. 1(1):53-60. |
|
|
|
ICUC (2006). Baobab Manual, Field manual for extension workers and farmers. University of Southampton, Southampton, UK. 4: P. 27. |
|
|
|
ICUC (International Center for Underutilized Crops) (2010). Baobab (Adansonia digitata L.), Manuel pratique pour les médiateurs et paysans. University of Southampton, Southampton, UK. 1:34. |
|
|
Kozlowski TT (2002). Physiological ecology of natural regeneration of harvested and disturbed forest stands: implications for forest management. Forest. Ecol. Manage. 158:195-221.
Crossref |
|
|
|
Levitt J (1974). Introduction to plant physiology. CV Mosby Company USA. pp. 277-286. |
|
|
Loha A, Tigabu M, Teketay D, Lundkvist K, Fries A (2006). Provenance variation in seed morphometric traits, germination, and seedling growth of Cordia Africana Lam. New For. 32: 71-86.
Crossref |
|
|
|
Mathur RS, Sharma KK, Rawat MMS (1984). Germination behaviour of provenances of Acacia nilotica sp. indica. Indian For. 110:435-449. |
|
|
|
Mayer AM, Poljakoff-Mayber A (1989). The Germination of Seeds. 4th Edn Pergamon Press, Oxford, UK. |
|
|
Milberg P, Lamont BB (1997). Seed/cotyledon size and nutrient content play a major role in early performance of species on nutrient-poor soil. New Phytol. 137:65-72.
Crossref |
|
|
Munthali CRY, Chirwa PW, Akinnifesi FK (2012). Genetic variation among and within provenances of Adansonia digitata L. (Baobab) in seed germination and seedling growth from selected natural populations in Malawi. Agroforest. Syst. 86:419-431.
Crossref |
|
|
|
Owen J (1970). The Medico-Social and Cultural Significance of Adansonia digitata L. (Baobab) in African Communities. African Notes: Inst. Afr. Stud. Occas. Publ. 60:24-36. |
|
|
Raddad EL AY (2007). Ecophysiological and genetic variation in seedling traits and in first-year field performance of eight Acacia Senegal provenances in the Blue Nile, Sudan. New For. 34:207-222.
Crossref |
|
|
Randall A, Baum JL, Small WF (1998). Biogeography and Floral Evolution of Baobabs Adansonia, Bombacaceae as Inferred From Multiple Data Sets. Syst. Biol. 47:181-207.
Crossref |
|
|
Razanameharizaka J, Grouzis M, Ravelomanana D, Danthu P (2006). Seed storage behavior and seed germination in African and Malagasy baobabs (Adansonia species). Seed. Sci. Res. 15:83-88.
Crossref |
|
|
|
Redondo-Gómez S, Naranjo EM, Garzón O, Castillo JM, Luque T, Figueroa ME (2007). Effects of Salinity on Germination and Seedling Establishment of Endangered Limonium emarginatum (Willd.) O. Kuntze. J. Coastal Res. 24 (1): 201-205 |
|
|
|
Ren J, Tao L, Liu XM (2002). Effect of sand burial depth on seed germination and seedling emergence of Calligonum L. species. J. Arid. Environ. 51:603-611. |
|
|
|
Sacande M, Charlotte R, Sanon M, Joker D (2006). Adansonia digitata L. Seed Leaflet No. 109. Forest & Landscape Denmark, Hørsholm, Denmark, and Millenium Seed Bank Project, Kew, UK. P. 2. |
|
|
Salack S, Muller B, Gaye AT (2011). Rain-based factors of high agricultural impacts over Senegal. Part I: integration of local to sub-regional trends and variability. Theor. Appl. Climatol. 106(1-2):1-22.
Crossref |
|
|
|
Salazar R, Quesada M (1987). Provenance variation in Guazuma ulmifolia L. in Costa rica. Commonw. For. Rev. 66:317–324. |
|
|
|
Sidibé M, Williams JT (2002). Baobab. Adansonia digitata. Fruits for the Future 4. International Centre for Underutilised Crops, Southampton, UK. P. 96. |
|
|
|
Willan RL (1985). A guide to forest seed handling with particular reference to the topics. FAO Forestry P 20/2. |