ABSTRACT
The potential of cowpea to address food security in Burkina Faso in particular is well established as it is a nutritious, cash and cover crop. However, there is limited information on existing germplasm diversity in Burkina Faso. This study was designed to gather some information on the genetic diversity in a set of cowpea lines introduced from different breeding programs. The diversity was therefore assessed using 181 single nucleotide polymorphism (SNP) markers on 50 cowpea lines. Leaf samples of young plants were collected using LGC genomics genotyping platform protocols for DNA extraction and genotyping. Data were then analyzed using 3 software for pair-wise distance, phylogenetic pattern by UPGMA and for the descriptive statistics determination. The phylogenetic pattern of this germplasm revealed seven clusters. The lines were almost grouped based on their geographical origin, and the breeding background. Thus, materials which originated from Burkina Faso were clustered in the same group while those from IITA/Nigeria were also almost all clustered in the same group. The genetic distance was low (≤0.29) suggesting a narrow genetic base in the cowpea germplasm used in this study. SNPs were efficient in the study of the diversity and a core collection of 20 lines was generated for further use in the breeding program.
Key words: Cowpea, single nucleotide polymorphisms (SNPs), genetic diversity, germplasm, Burkina Faso.
Despite considerable phenotypic diversity that exists in cultivated cowpea germplasm, there is limited genetic variability in cowpea breeding programs (Pasquet, 1999, 2000). Breeding programs must focus most of their efforts on rapid delivery of varieties with a specific range of production and quality traits. However, most of the breeding programs tend to cross and re-cross cultivars with similar yield potentials and other traits and many of these cultivars are related to some degree. This leads to reduced genetic variability among cultivars that are released and among advanced breeding lines in the program, and in most cases the released varieties and the advanced lines are used as parents in new breeding cycles (Fang et al., 2007). The lack of diversity is a special concern because cowpea appears to have lower inherent genetic diversity than other cultivated crops as a result of a hypothesized single domestication event (Pasquet, 1999, 2000). Markers based on single nucleotide polymorphisms (SNPs) have rapidly gained the center stage of molecular genetics during the recent years due to their abundance in the genomes and their amenability for high-throughput detection formats and platforms (Mammadov et al., 2012). Among these platforms is the LGC genomics’ Kompetitive Allele Specific PCR (KASP) combined with the SNP line platforms in United Kingdom. SNP markers are increasingly being used for a large number of genetic studies including genetic diversities. Such studies have been reported in pea (Deulvot et al., 2010), cowpea (Huynh et al., 2013; Egbadzor et al., 2014), and cassava (Thompson, 2013). SNPs provide the simplest form of molecular markers as a single nucleotide base is the smallest unit of inheritance, and therefore, they can provide a large number of markers to be used in diversities or in marker assisted breeding. SNPs are co-dominant markers and they are most often linked to genes, and thus, they are the most attractive genetic markers in genetic studies (Jiang, 2013). The use of these markers could therefore help group germplasm which will also help breeders make informed choice of parents for breeding purposes. SNP markers therefore help in decision making when the variability within the germplasm is known. Available breeding materials should be well known and described in any breeding program for any crop for better exploitation of the potential variability. The description of the variability among breeding materials can be done by morphological, biochemical, and molecular characterization. There exist important cowpea genetic materials in the cowpea breeding program in Burkina Faso. However, no in-depth investigation has been made to establish the variability using molecular markers. Therefore, the objective of this study was to molecularly assess the genetic diversity in the set of cowpea germplasm using SNP markers.
Cowpea genotypes
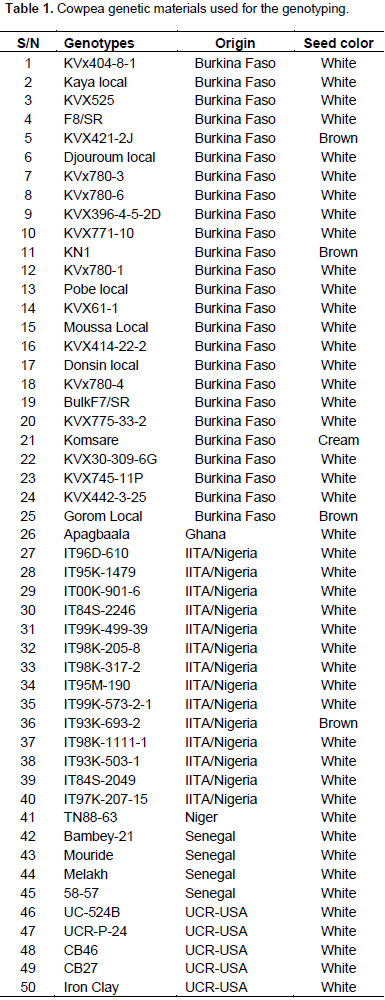
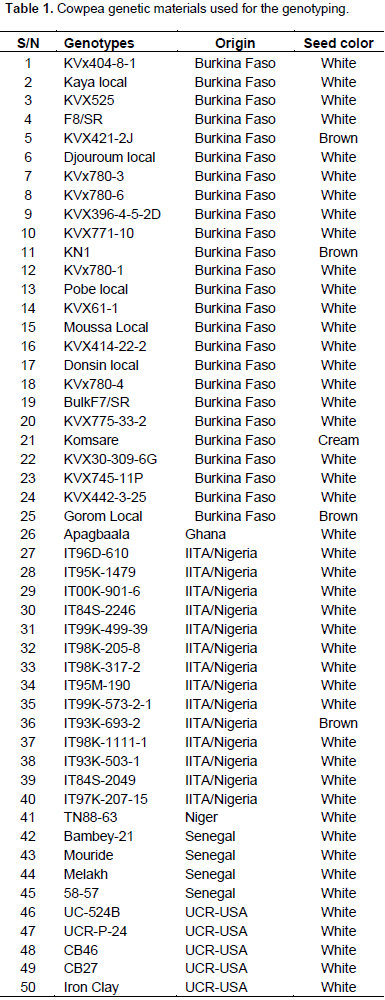
Fifty cowpea genotypes were used for the genetic diversity study using SNP markers. The origin and seed coat color of the 50 cowpea genotypes used in the study have been described in Table 1
SNP genotyping
Leaf samples of 2-weeks old plants were collected in a 96-wells plate and sent to LGC genomics in the United Kingdom for DNA extraction and SNP genotyping. The KASP technology as described by Thompson (2013)was used for the genotyping at LGC genomics. The DNA was extracted using LGC genomics internal protocol described. One hundred and eighty-one SNP markers selected from the Generation Challenge Programme (GCP) platform were used. After excluding the SNPs that were not informative enough (more than 10% missing data), a total of 170 markers and 47 cowpea lines were used for further analysis.
Analysis of genetic diversity
Pair-wise genetic distances between genotypes were measured with the software GGT 2.0 (Van Berloo, 2008)based on the allele-sharing method (Bowcock et al., 1994). The simple matching algorithm considers both presence and absence of markers in calculating degrees of similarity. Phylogenetic relationships dendrogram were generated based on the genetic-distance matrix using the un-weighted pair group method (UPGMA) with the software MEGA 6.0 (Tamura et al., 2013). Descriptive statistics like polymorphism information content (PIC) value, major allele frequency (MAF), and expected heterozygosity (He) were calculated for all the SNPs using PowerMarker 3.25 software (Lui and Muse, 2005). A core collection of genotypes was generated from GGT2.0 software based on the maximum diversity sum.
Descriptive statistics
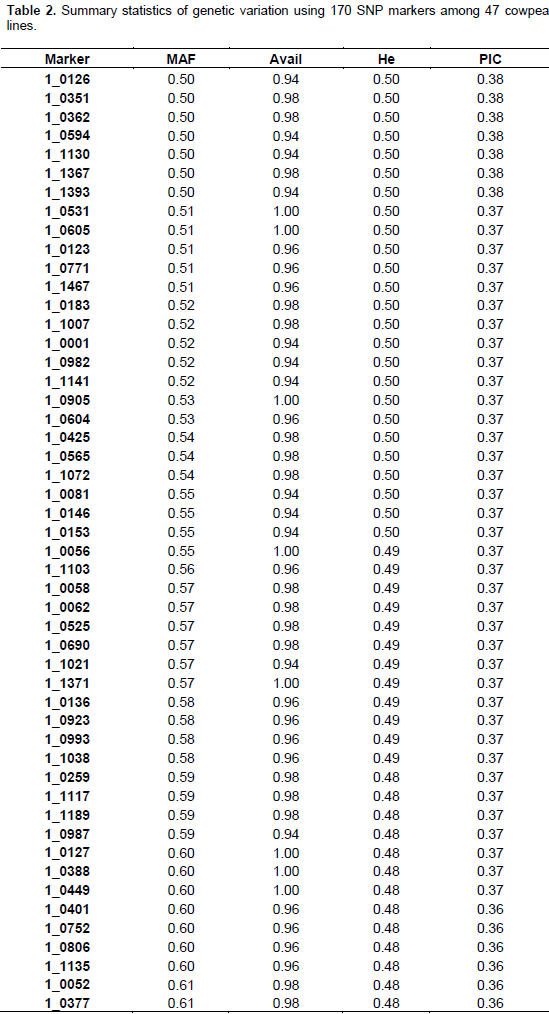
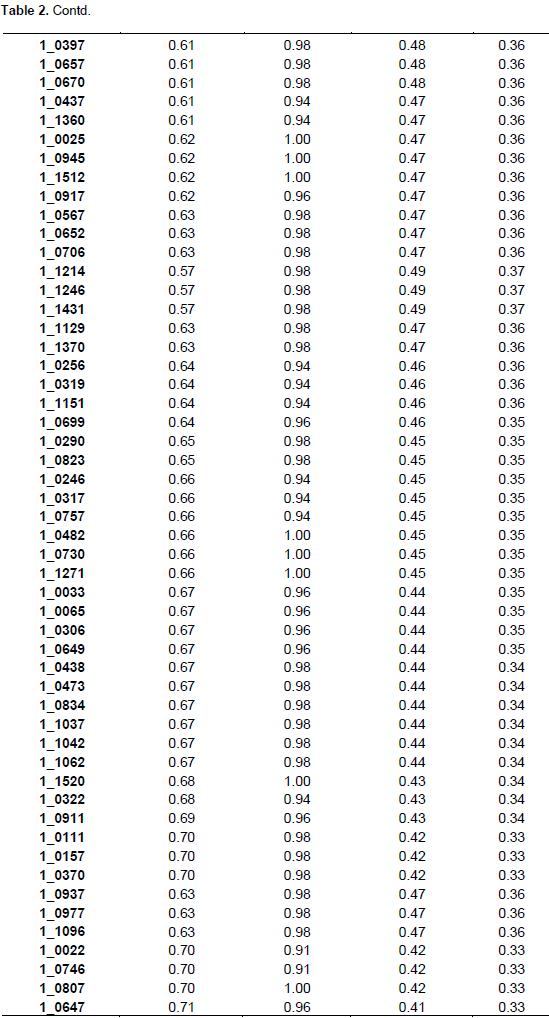
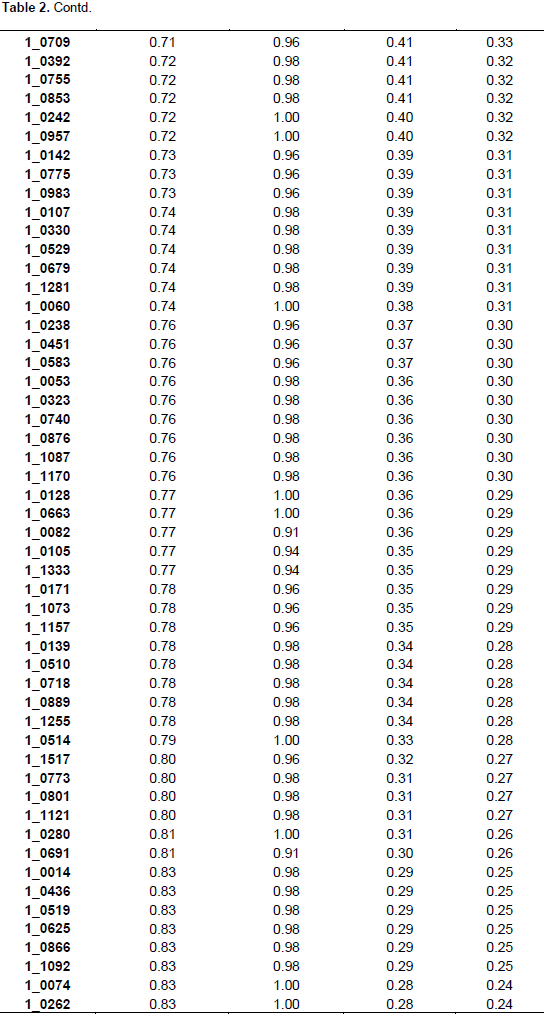
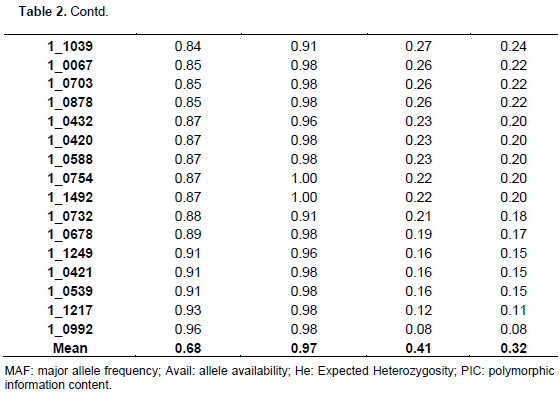
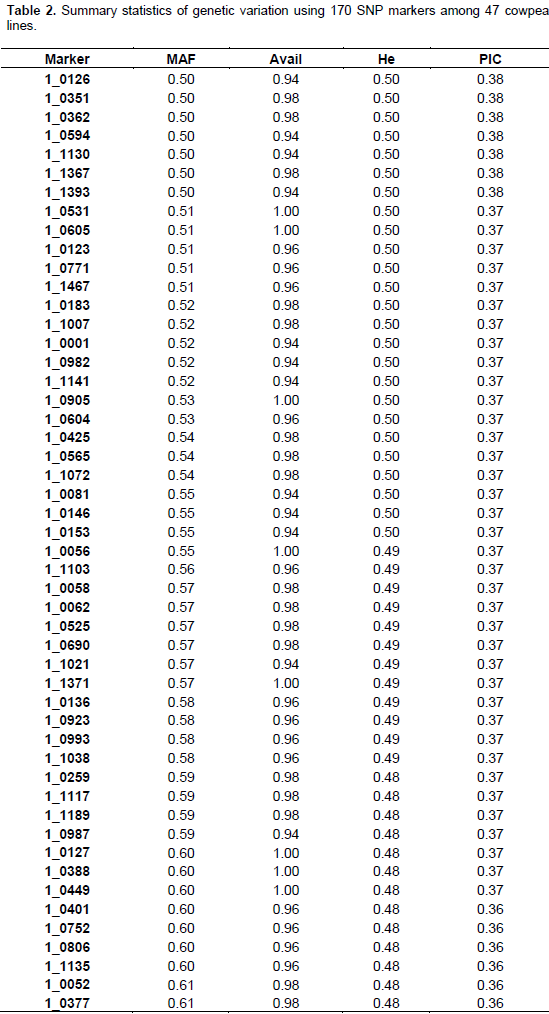
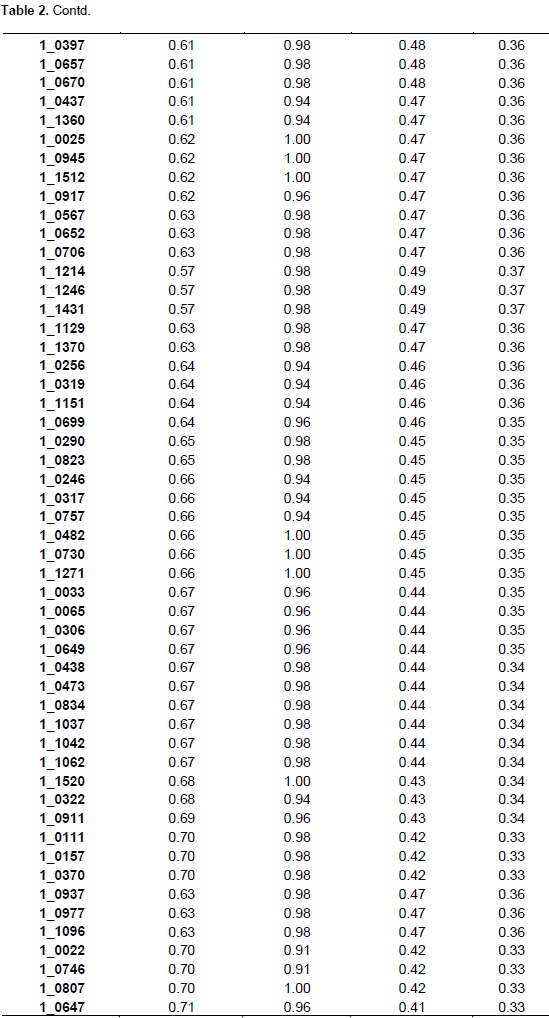
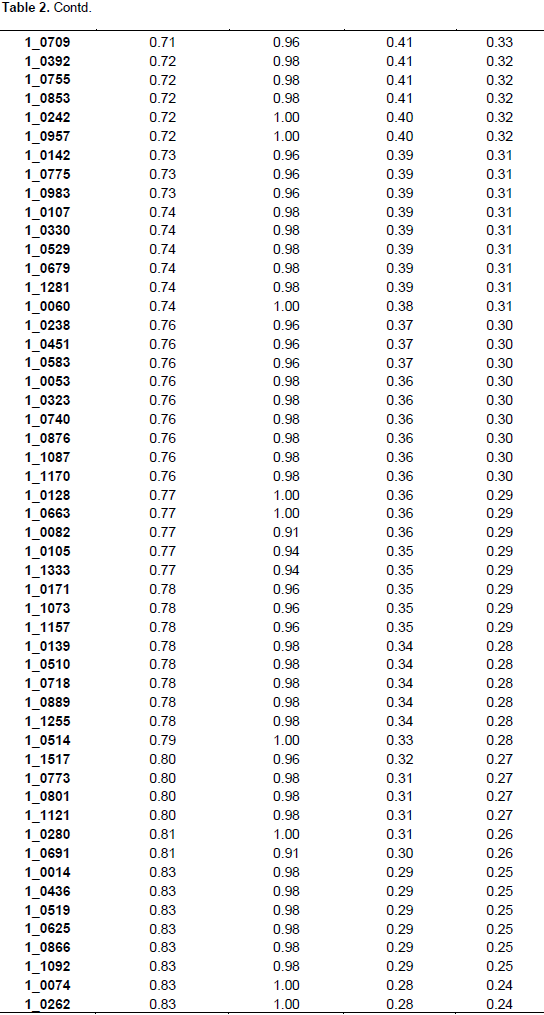
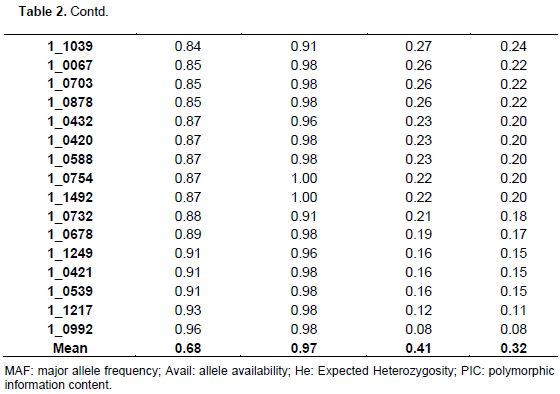
The summary statistics for major allele frequencies (MAF), expected heterozygosity (He), and polymorphic information content (PIC) is presented in Table 2. A low expected heterozygosity (0.08) was observed with the SNP marker (1_0992) that has the high major allele frequency (0.96). The mean of the expected heterozygosity was 0.41 and that of the major allele frequency was 0.68. The allele frequencies of all the SNP markers were greater than their corresponding expected heterozygosity values. The allele frequencies of all the markers were below 0.95 except 1_0992 (0.96), indicating the polymorphic nature of the SNP markers used. The PIC values ranged from 0.08 (1_0992) to 0.38 with an average of 0.32. Out of the 177 SNPs, 170 were useful representing 96.04% of the total. One hundred and three SNPs were the most informative markers with a PIC value greater than the mean which represents 60.59% of the useful SNPs. Out of the 103 SNPs seven have a PIC of 0.38, 40 a PIC of 0.37, 26 a PIC of 0.36, 13 a PIC of 0.35, nine a PIC 0.34, and eight a PIC of 0.33. The seven most informative markers were 1_0126,1_0351, 1_0362, 1_0594, 1_1130, 1_1367, and 1_1393.
Core collection of cowpea germplasm
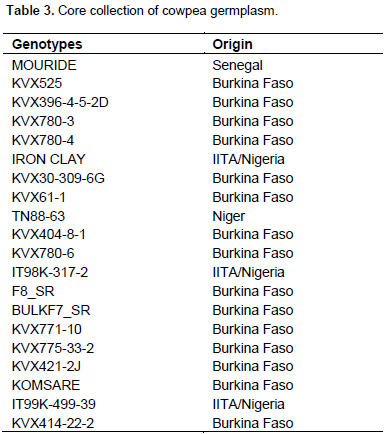
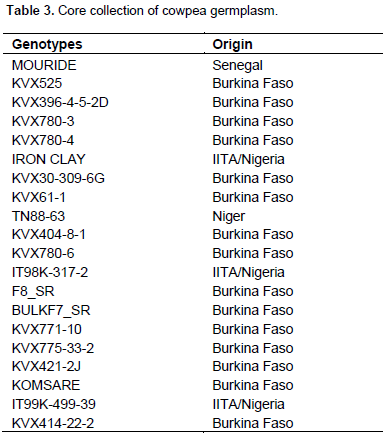
Twenty cowpea genotypes forming a core collection is presented in Table 3. This collection comprises 15 improved varieties from Burkina Faso, 3 advanced breeding lines from International Institute of Tropical Agriculture (IITA) in Ibadan – Nigeria, 1 line each from Niger and Senegal.
Phylogenetic relationships between cowpea lines
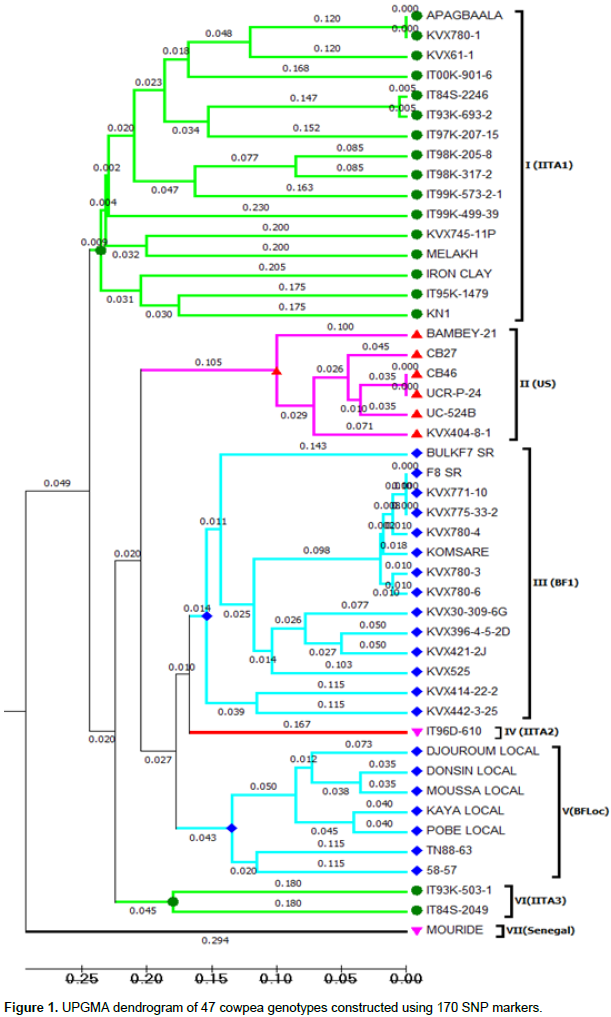
The cowpea lines were grouped into 7 clusters based on genetic distance based on the allele sharing similarity. The cluster analysis showed that lines are generally grouped together according to their geographical origin and traditional genetic background (Figure 1). Cluster VII and IV can be considered as outliers as they contained only one line (Mouride, IT86D-610). Cluster I consisted of 16 genotypes, Cluster II had 6 lines, Cluster III had 14 lines, Cluster V contains 7 lines, and Cluster VI has 2 lines. United States and Burkina Faso landraces respectively fell into Clusters II (US) and V (BF2Loc) while the improved varieties were all in Cluster III (BF1). The genetic materials from IITA fell into 2 main Clusters I (IITA1) and VI (IITA3) with slight mixture of some improved varieties from Burkina, Senegal, and Ghana.
In the present study, one hundred and seventy SNP markers were used to genotype forty-seven cowpea lines. The results showed a good level of polymorphism but a moderate level of diversity based on the average polymorphic information content values (0.32). Almost all of the 47 lines shared a very narrow genetic distance (≤0.29) which is consistent with the results reported by Li et al. (2001). Moreover, the markers enabled the grouping of lines based on their similarity. Likewise, the SNP markers were able to associate more or less the cluster to the geographical origin of the line. Breeding programs generally work within restricted pools of genetic variation (Huynh et al., 2013)and might be the cause of this narrow genetic diversity observed in this study. A number of authors have come to the conclusion that cowpea lacks significant variability (Pasquet, 1999, 2000; Fang et al., 2007). Narrow genetic base has also been observed within different lines from breeding programs (Li et al., 2001). The materials from IITA collection have been widely used by different breeding programs in different countries. This can explain the relatedness between some cowpea improved varieties from Burkina Faso (KVx745-11P, KN1, KVx780-6, and KVx61-1). Looking at also the pedigree of Melakh (IS86-292 x IT83S-742-13) (Diouf and Hilu, 2005), it becomes easy to understand why this line fell into the cluster of IITA lines because of its relatedness among line from the IITA breeding program. Huynh et al. (2013)provided some useful assumptions that tend to explain the reduction of the genetic distance among cowpea wild types, landraces, and improved germplasm within African germplasm accessions and among African and Non-African germplasm accessions.
These authors concluded that the small genetic differentiation observed between the African and non-African collections indicated that the entire genetic diversity in the African germplasm might already have spread over cowpea-growing regions in the world as a whole although not completely within any single region. Nevertheless, the clustering of these 47 lines into 7 distinct groups gives important insights that can improve the efficiency of germplasm used in cowpea for breeding purposes. With the exception of the materials from Senegal (Bambey in Cluster II, Mouride in Cluster VII, 58-57 in Cluster III, and Melakh in Cluster I), from Niger (TN88-63 in Cluster III), and from Ghana (Apaagbala in Cluster I) that were not grouped according to their geographical origin, the rest were clustered based on their country of origin. That could be helpful for new ways of genetic improvement of cowpea by exchanging material from different countries to broaden the genetic base of the crop. In contrast with these findings, a numbers of genetic diversity studies conducted on cowpea have reported absence of correlation between geographical origin of the accessions and their clustering pattern (Asare et al., 2010; Egbadzor et al., 2014). This was also observed in a genetic diversity study in maize using SSR markers (Oppong, 2013). In this study, the genotypes were clustering following a regional basis of maize cultivation in Ghana. The differences shown between landraces and the improved varieties from Burkina Faso may also be useful as a little diversity still exists among the local germplasm for new variety development. SNP markers have demonstrated their capacity in assessing genetic diversity in cowpea (Huynh et al., 2013; Egbadzor et al., 2014). Varshney et al. (2007)reported on the robustness of SNP markers.
As compared to SSR markers, SNPs are more robust as they are able to detect slight changes in the genome and discriminate genotypes. This assumption is confirmed by the findings from a genetic diversity study on sweet cherry (Prunus avium L.) (Marti et al., 2012). In this study, SNP markers were able to discriminate mutants from their original parents than SSR markers. In addition, SNP markers confirmed parentage and also determined relationships of the accessions in a manner consistent with their pedigree relationships. The latter statement confirmed our findings. Lines like Melakh from Senegal, KVx745-11P from Burkina Faso was grouped with the IITA accessions because of the large contribution in their genome of materials from IITA. Extension of gene pool is important for crop improvement (Varshney et al., 2007). As such a core collection of 20 lines was proposed from this study based on the maximum diversity among them. Several genetic diversity studies have been conducted in cowpea (Pamella and Gepts, 1992; Vaillancourt and Weeden, 1992; Fotso et al., 1994; Coulibaly et al., 2002; Ba et al., 2004). Despite of the presence of little diversity within the collection used for this study and the core collection, the separation of the broader germplasm of cowpea landraces into gene pools as done by Huynh et al. (2013)could be useful for expanding the genetic diversity within breeding materials and could lead to development of more efficient strategies and genetic gain within future breeding programs.
The present study was undertaken to determine the genetic variability in a set of germplasm used by INERA Cowpea Breeding Program in Burkina Faso using SNP markers. The germplasm used has some moderate variability with narrow genetic base. These results were comparable to previous studies that have also reported the narrow genetic base of cowpea. The phylogenetic patterns and clustering of relatively similar individuals into groups provide important information on the germplasm used for cowpea improvement. The materials were grouped based on the geographic origin and the genotypic background. Materials from United State/University of California Riverside clustered together. Likewise, materials from IITA/Nigeria, Burkina Faso clustered in country base. SNP markers were able to group the genotypes in a way that they could be used to link the genotype clusters and their pedigree. A panel of 20 genotypes representing the maximum variability of the germplasm used in the study was generated based on the maximum diversity sum. This panel constituted a collection that could be together with the information on the clustering of great importance for further plant breeding to develop superior varieties of cowpea.
The authors have not declared any conflict of interests.
This work was supported in part by The Generation Challenge Program, USAID Legume Innovation Lab/ Innovation Lab for climate Resilient Cowpea, Alliance for Green Revolution in Africa, Kirkhouse Trust and West Africa Centre for Crop Improvement. The guidance of Dr. J. D. Ehlers is gratefully appreciated.
REFERENCES
|
Asare AT, Gowda BS, Galyuon IKA, Aboagye LM, Takrama JF, Timko MP (2010). Assessment of the genetic diversity in cowpea (Vigna unguiculata (L.) Walp) germplasm from Ghana using simple sequence repeat markers. Plant Genetic Resources, 8:142-150.
Crossref
|
|
|
|
Ba FS, Pasquet RS, Gepts P (2004). Genetic diversity in cowpea [Vigna unguiculata (L.) Walp.] as revealed by rapd markers. Genetic Resources and Crop Evolution, 51:539-550.
Crossref
|
|
|
|
|
Bowcock AM, Ruizlinares A, Tomfohrde J, Minch E, Kidd JR, Cavallisforza LL (1994). High-resolution of human evolutionary trees with polymorphic microsatellites. Nature 368:455-457.
Crossref
|
|
|
|
|
Coulibaly S, Pasquet RS, Papa R, Gepts P (2002). AFLP analysis of the phenetic organization and genetic diversity of Vigna unguiculata (L.) Walp reveals extensive gene flow between wild and domesticated types. Theor. Appl. Genet. 104:358-366.
Crossref
|
|
|
|
|
Deulvot C, Charrell H, Marty A, Jacquin F, Donnadieu C, Lejeune-Henaut I, Burstin J, Aubert G (2010). Highly-multiplexed SNP genotyping for genetic mapping and germplasm diversity studies in pea. BMC Genomics 11:468-477.
Crossref
|
|
|
|
|
Diouf D, Hilu KW (2005). Microsatellites and rapd markers to study genetic relationships among cowpea breeding lines and local varieties in Senegal. Genet. Resour. Crop Evol. 52:1057-1067.
Crossref
|
|
|
|
|
Egbadzor KF, Ofori K, Yeboah M, Aboagye LM, Opoku-Agyeman MO (2014). Diversity in 113 cowpea [Vigna unguiculata (L.) Walp] accessions assessed with 458 SNP markers. Springer Plus, 3.
Crossref
|
|
|
|
|
Fang J, Chao CCT, Roberts PA, Ehlers JD (2007). Genetic diversity of cowpea [Vigna unguiculata (L.) Walp.] in four West African and United State of America breeding programs as determined by AFLP analysis. Genetic Resources and Crop Evolution, 54:1197-1209.
Crossref
|
|
|
|
|
Fotso M, Azanza JL, Pasquet RS, Raymond J (1994). Molecular heterogeneity of cowpea (Vigna unguiculata Fabaceae) seed storage proteins. Plant Systematics and Evolution, 191:39-56.
Crossref
|
|
|
|
|
Huynh BL, Close TJ, Roberts PA, Hu Z, Wanamaker S, Lucas MR, Chiulele RM, Cissé N, David A, Hearne S, Fatokun C, Diop NN, Ehlers JD (2013). Gene pools and the genetic architecture of domesticated cowpea. Plant Genome 6:8.
Crossref
|
|
|
|
|
Jiang GL (2013). Molecular markers and marker-assisted breeding in plants. In: Anderson SB (ed.) Plant Breeding from Laboratories to Field. Croatia: InTech pp. 45-83.
Crossref
|
|
|
|
|
Li CD, Fatokun CA, Ubi B, Singh BB, Scoles GJ (2001). Determining genetic similarities and relationships among cowpea breeding lines and cultivars by microsatellite markers. Crop Science, 41:189-197.
Crossref
|
|
|
|
|
Lui K, Muse SV (2005). Powermarker: Integrated analysis environment for genetic marker data. Bioinformatics 21:2128-2129.
Crossref
|
|
|
|
|
Mammadov J, Aggarwal R, Buyyarapu R, Kumpatla S (2012). SNP markers and their impact on plant breeding. International Journal of Plant Genomics, pp:1-12.
Crossref
|
|
|
|
|
Marti FIA, Athanson B, Koepke T, Front I Forcada C, Dhingra A, Oraguzie N (2012). Genetic diversity and relatedness of sweet cherry (Prunus avium L.) cultivars based on single nucleotide polymorphic markers. Frontiers in Plant Science, 3:1-13.
|
|
|
|
|
Oppong A (2013). Improving ghanaian maize (Zea mays L.) landraces for yield and resistance or tolerance to streak virus disease. Doctor of Philosophy, University of Ghana, Legon. 204.
|
|
|
|
|
Pamella L, Gepts P (1992). Genetic relationships within Vigna unguiculata (L.) Walp. Based on isozyme analyses. Genet. Resour. Crop Evol. 39:71-88.
|
|
|
|
|
Pasquet RS (1999). Genetic relationships among subspecies of Vigna unguiculata L. Walp. based on allozyme variation. Theor. Appl. Genet. 98:1104-1119.
Crossref
|
|
|
|
|
Pasquet RS (2000). Allozyme diversity of cultivated cowpea Vigna unguiculata L. Walp. Theoretical and Applied Genetics,101:211-219.
Crossref
|
|
|
|
|
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013). Mega6: Molecular evolutionary genetics analysis version 6.0. Molecular Biology and Evolution, 30:2725-2729.
Crossref
|
|
|
|
|
Thompson R (2013). Genetic analysis of postharvest physiological deterioration in cassava (Manihot esculenta Crantz) storage roots. PhD Thesis. University of Ghana.
|
|
|
|
|
Vaillancourt RE, Weeden NF (1992). Chloroplast DNA polymorphism suggests nigerian center of domestication for the cowpea, Vigna unguiculata (L.). American Journal of Botany,79:1194-1199.
Crossref
|
|
|
|
|
Van Berloo R (2008). GGT 2.0: Versatile software for visualization and analysis of genetic data. Journal of Heredity, 99:232-236.
Crossref
|
|
|
|
|
Varshney RK, Chabane K, Hendre PS, Aggarwal RK, Graner A (2007). Comparative assessment of EST-SSR, EST-SNP and aflp markers for evaluation of genetic diversity and conservation of genetic resources using wild, cultivated and elite barleys. Plant Sci. 173:638-649.
Crossref
|
|