ABSTRACT
Soil microorganisms play an important role in the decomposition of organic and inorganic matter in soil, being capable of breaking down different types of xenobiotic substances in soil. Application of herbicides in crops is commonly used in agriculture to increase agricultural productivity and weed destruction. Two sulfonylurea herbicides (tribenuron methyl and nicosulfuron) were applied to the soil samples in different doses. The effect of the herbicides on soil enzymatic activities and on the main groups of microorganisms involved in the soil nitrogen cycle (ammonifying, nitrifying and denitrifying bacteria) was studied under laboratory conditions. As far as the values ​​of enzymatic activities are concerned, it was determined that they were strongly inhibited by the application of these two herbicides. Values of the ​​dehydrogenase and urease activities from the experimental variants, most closely reflects the inhibitory effect of herbicides. Enzymatic indicator of soil quality shows values ​​recorded between 0.390 to 0.541 for soils treated with tribenuron-methyl, respectively 0.401 to 0.431 in those treated with nicosulfuron. The results showed that nicosulfuron had a lower inhibitory effect on the enzymatic activity of soil from tribenuron-methyl. Ammonifying, nitrifying, and denitrifying bacteria responded differently to the action of herbicides, depending on the dose that was applied. Bacterial indicator of soil quality recorded values between 1.645 to 1.706 ​​for soils treated with tribenuron-methyl and 1.651 to 1753 in those treated with nicosulfuron. Tribenuron-methyl and nicosulfuron affect negative bacterial metabolic activity and bacterial communities.
Key words: Nicosulfuron, tribenuron-methyl, soil, enzymatic activity, bacteria nitrogen cycle.
Most xenobiotic substances used in agriculture have more or less toxic effects on living organisms. Studies attempt to determine the impact of xenobiotic substances on soil microorganism communities, given the role these communities play in the biogeochemical cycle in nature. Application of xenobiotic substances in arable crops to
destroy weeds and pests results in irreversible soil pollution. This process has toxic effects on soil microorganism communities, which are essential players in organic matter decomposition. Among the negative effects of xenobiotic substances applied on soil are their quantitative and qualitative reduction, biochemical activity inhibition and changes in community structure which affect its diversity and weight of microbial communities. The present study comparatively investigates the impact of two sulfonylurea herbicides, that is, tribenuron-methyl and nicosulfuron, on microorganisms that are involved in the nitrogen cycle in the soil.
Sulfonylurea herbicides are a distinctive group of herbicides that are widely used in arable crops to destroy weeds. They have become very popular, due to their low toxicity to small mammals, low application rates and effective weed control (
Sarmah and
Sabadie, 2002).
The application of herbicides to destroy weeds in arable crops has beneficial effects on agricultural production, but negatively influences soil microorganism growth, metabolic activity, and population dynamics (Sannino and Gianfreda, 2001; Zhou et al., 2009). Such negative effects were regularly reported for other xenobiotic substances, such as fungicides (Latijnhouwers et al., 2000), insecticides (Yao et al., 2006; Cycon et al., 2010) or heavy metals (Avidano et al., 2005; Gülser and Erdo?an, 2008), when investigating their impact on soil ecosystems.
The impact of sulfonylurea herbicides on soil microorganism communities has been studied for chlorsulfuron, thifensulfuron, amidosulfuron (Borozan et al., 2010), tribenuron-methyl, thifensulfuron-methyl (Andersen et al., 2001), bensulfuron-metil (
Saeki and
Toyota, 2004) and metsulfuron-methyl (Wang et al., 2010).
Cinosulfuron and bensulfuron applied in normal rates and 100 times higher had no effect on the total bacteria number and respiratory activity in the soil, but inhibited nitrification. Of the two sulfonylurea herbicides applied, bensulfuron showed more toxic effects than cinosulfuron on 17 bacteria strains and 12 fungal strains used for analysis (
Gigliotti and
Allievi, 2001).
A combination of two sulfonylurea herbicides (60% bromoxynil + 3% prosulfuron) used to provide a broad spectrum weed control in arable crops indicated effects on microorganism growth and activity. Their impact on bacterial communities (heterotrophic bacteria, but mostly nitrifiers), actinomycetes and fungi was demonstrated. The dehydrogenase activity proved to be an important indicator of the side effects of these herbicides (
Pampulha and
Oliveira, 2006).
Cinosulfuron, another sulfonylurea herbicide, causes the inhibition of microorganism community development in the soil. Aerobic bacteria, autotrophic nitrifying bacteria,
Azotobacter bacteria and fungi showed increased sensitivity to this herbicide (
Allievi and
Gigliotti, 2001).
Enzymatic activities of soil biota are reliable biomarkers for assessing the effects of herbicide application on ground and below-ground ecosystems. Consequently, certain enzymes, such as oxidases (e.g., dehydrogenase, catalase) or hydrolases (e.g., protein phosphatase, urease, inverters), are routinely used for this purpose (Benitez et al., 2004). Among various bacterial enzymes, the dehydrogenase activity provides the most `natural` approach for understanding the impact of herbicides on agricultural ecosystems because it is directly related to soil respiratory processes; therefore, this effect criterion is often employed in toxicity tests with herbicides.
Xenobiotic biodegradation takes place mainly within rhisophere, wherein the populational diversity and abundance of bacterial biotas are higher than in the upper or lower soil horizons. This allows an effective decomposition of various chemical compounds, such as metsulfuronmethyl (Ghani and Wardle, 2001), dichlorophenoxyacetic acid (Shaw and Burns, 2004), and atrazine (Piutti et al., 2002). Several groups of soil bacteria are able to use herbicides as an energy source for their growth and development. For example, gram-negative and gram-positive bacteria (van Eerd et al., 2003; Gimsing et al., 2004) as well as heterotrophic bacteria (Merini et al., 2007; Zabaloy et al., 2010) were shown to use glyphosate as nitrogen, carbon, or phosphor source. As a result, assessing the structure of local soil microbiota may allow researchers to identify anthropic-induced soil disturbance.
This study involves highlighting the action of xenobiotic substances (herbicides of the sulfonylureas group), tribenuron-methyl and nicosulfuron on the community of microorganisms implicated in biogeochemical cycle of nitrogen in soil and on the main enzymatic activities that decompose organic matter.
Uncontaminated soil (cambic chernozem) was sampled to 0 to 20 cm from the experimental fields of the Banat’s University of Agricultural Sciences and Veterinary Medicine of Timisoara (Romania). The samples were spiked, under laboratory conditions, with two sulfuronureic herbicides, namely tribenuron-methyl and nicosulfuron. Tribenuron-methyl, also known as 2-methox-6 -[[[[( 4-methyl-1 ,3,5-triazine-2-il) methylamino] carbon] sulfone] benzoic amino acid is also present in herbicides from local markets under the trade name „Helmstar” (Tellurium Chemical, Romania). Nicosulfuron known as 2 - [[(4,6-dimethoxypyrimidin-2-il) aminocarbon] aminosulfon]-N, N-dimethyl-3-pyridinecarboxamide is also present in herbicides from local markets under the trade name „Mistral” (ISK Biosciences Europe, Romania).
Soil treatment with herbicides
The soil was passed through a 2 mm sieve and put in polyethylene bags. Different doses of herbicides were prepared using distilled water, and then used for spiking the soil samples so that the soil humidity was 40%. The dose of herbicide applied onto the dry soil was calculated assuming a uniform distribution in the plow layer (Atlas et al., 1978). Apart from the control group (R: 0 μg tribenuron-methyl, 0 μg nicosulfuron) the following herbicide concentrations were considered in the present study: (1) group A, A1: 0.6 μg tribenuron-methyl, A2: 0.4 μg nicosulfuron; (2) group B, B1: 1.2 μg tribenuron-methyl, B2: 0.8 μg nicosulfuron; (3) group C, C1: 1.8 μg tribenuron-methyl, C2: 1.2 μg nicosulfuron; (4) group D, D1: 3 μg tribenuron-methyl, D2: 2 μg nicosulfuron; (5) group E, E1: 4.2 μg tribenuron-methyl, E2: 2.8 μg nicosulfuron. After herbicide application, the samples were incubated for 7 days, at 28°C.
Enzymatic activity
The following enzymatic activities were assessed: catalase activity (CA), actual dehydrogenase activity (ADA), potential dehydrogenase activity (PDA), and urease activity (UA).
CA was determined by using the permanganometric method, that is, titration with a 0.05 N KMnO4 solution until a weak pink coloration occurred). The enzymatic activity was expressed as milligrams of undecomposed oxygenated water (H2O2) per gram of soil (Dragan-Bularda, 2000).
Actual dehydrogenase activity (ADA) and potential dehydrogenase activity (PDA) were detected using the method described by Casida et al ., (1964). This technique involves the incubation of soil samples with TTC (2, 3, 5-triphenyltetrazolium chloride) for 48 h, at 37°C. ADA and PDA was shown as milligrams formazan per gram of sediment/soil. The adsorbance of reduced TTC was measured at 485 nm.
Urease activitity (UA) assesses the rate of urea decomposition in ammonia (NH3) and carbon dioxide (CO2). The reaction mixture was incubated at 37°C, for 24 h. The ammonia was extracted with a 2 N potassium chloride (KCl) solution, and determined by spectrophotometric nesslerization at 445 nm; the corresponding calibration curve was created using an ammonium chloride (NH4SO4) solution (Dragan-Bularda, 2000).
Ecophysiological groups of bacteria
Serial dilutions were performed from the herbicide-spiked soil samples (viz. 10-1 to 10-6). These solutions were incubated on elective culture media for 7 to 21 days (28°C), depending on the preferences of investigated groups of bacteria.
Samples were incubated for a week at 27°C on Ashby culture medium to determine the number of nitrogen-fixing bacteria (NFB). The presence of aerobic NFB resulted in formation of a fine velum on culture medium surface or/and in marked and adherent rings on test-tube walls. Depending on the species, the veil was either greenish yellow (A. vinelandii) or brown (A. chroococum). In contrast, anaerobic NFB (Clostridium sp.) induced the formation of gas bubbles in positive test tubes (Zarnea, 1994).
The culture medium for ammonifying bacteria (AMB) had the following chemical composition: NaCl (0.5 g), peptone (2 g), distilled water (1000 ml). The samples were incubated under anaerobic conditions for 14 days, at 22°C. AMB number was assessed based on the reaction between ammonia and Nessler reagent; the Alexander table was used as screening benchmark (Cusa, 1996).
The culture medium for nitrifying bacteria (NB) consisted of standard saline traces (50 ml), (NH3)SO4 (0.5 g), CaCO3 (1 g), and distilled water (950 ml). Samples were incubated for 20 days, at 28°C. Tubes containing nitrate were identified with diphenylamine sulphuric acid. A blue colour reaction showed that nitrite and nitrate were formed, and therefore, the tube was scored positive (Dunca et al., 2007).
The denitrifying bacteria (DNB) were grown in selective culture medium, which contained: standard saline solution (50 ml), KNO2 (20 g), glucose (10 g), KCO3 (5 g), oligoelement solution (1 ml), and distilled water (1000 ml). The samples were incubated at 28°C, for 7 to 15 days, and diphenylamine-sulphuric acid was added in each test tube. Positive samples were colourless due to nitrate metabolization by DNB (Dunca et al., 2007).
Indicators of soil quality
The enzymatic indicator of soil quality (EISQ) and the bacterial indicator of soil quality (BISQ) were calculated by using the formulas (1) and (2), as proposed by Muntean (1995-1996, 1996):
EISQ = 1/n Σ Vr (s)/Vmax (i) (1)
where: EISQ - enzymatic indicator of soil quality, n - number of enzymatic activities, Vr (i) – individual, real value, Vmax (i) – theoretical, maximum, individual value.
BISQ = 1/n x Σ log10 N (2)
where: BISQ - bacterial indicator of soil quality, n - number of ecophysiological groups, N - number of bacteria belonging to each ecophysiological group.
Statistical analysis
Statistical data interpretation was conducted with Microsoft Excel 2003 and Statistica 10 software packages.
Our study showed that PDA and ADA are both inhibited by tribenuron-methyl and nicosulfuron application (Figure 1). However, ADA values were higher than PDA values, and therefore, this biomarker is more reliable when assessing the effect of these two herbicides on soil bacterial communities. A recent study showed that metsulfuron-methyl and 2,4-D (syn. 2,4 dichlorophenoxyacetic acid) greatly inhibited dehydrogenase activity (Zabaloy et al., 2008). Similarly, glyphosate application resulted in a significant decrease for the same enzymatic activity (Araújo et al., 2003; Zabaloy et al., 2008). Moreover, dehydrogenase activity proved to be an important indicator of side effects that are associated with the use of sulfuronureic herbicides (Pampulha and Oliveira, 2006). This information suggests that ADA may serve as relevant field biomarker for assessing the effect of herbicide application on soil bacterial biota.
The current experiment revealed that CA can be either stimulated or inhibited by herbicide application, depending on the herbicide type and exposure dose. UA and PDA, however, were, by far, less sensitive when applying different doses of tribenuron-methyl and nicosulfuron. Field studies found that metsulfuron-methyl reduced urease, amylase, and protease activity (Ismail et al., 1998). Similar results were reported for other herbicides, such as glyphosate or glufosinate, which displayed a dose-dependent effect on soil enzymatic activity (Accinelli et al., 2002; Accinelli et al., 2007; Lupwayi et al., 2007). In contrast, bromine application had discrepant effects on different soil enzymes (that is, cellulase, alkaline phosphatase, acid, sulphatase); thus, this compound was found to significantly reduce cellulase and sulphatase activity, but to increase phosphatase activity, as a potential resistance mechanism to herbicide action (Omar and Abdel-Sater, 2001).

Enzymatic indicator of soil quality shows values ??recorded between 0.390 to 0.541 for soils treated with tribenuron-methyl, respectively 0.401 to 0.431 in those treated with nicosulfuron. The measured values of EISQ were higher for tribenuron-methyl than for nicosulfuron, thereby indicating that nicosulfuron is more toxic for soil bacterial biota than tribenuron-methyl (Figure 2).
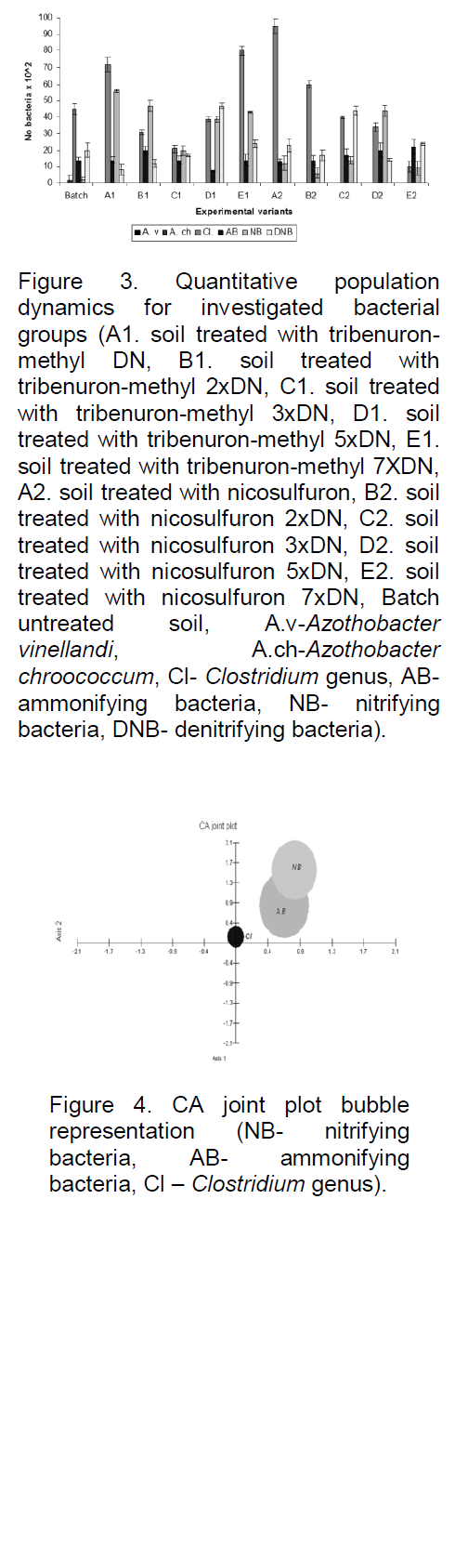
Our result indicated that herbicide application diminished in a dose-dependent manner AMB, NB, and NFB (Clostridium genus) number in soil. In contrast, DNB displayed a more heterogeneous pattern of variation (Figure 3), and hence, the quantitative population dynamics for this bacterial group cannot serve as a pertinent biomarker of herbicide-induced soil disturbance.
The bacteria of the Clostridium genus are the most sensitive bacterial group to tribenuron-methyl and nicosulfuron impact, with the strongest effect being reported when the exposure dose was seven-fold higher than the normal dose. In addition, quantitative population dynamics for these bacteria revealed a linear trend with exposure dose.
In this study, NFB of the Clostridium genus showed a distinctly different behaviour to herbicide exposure when
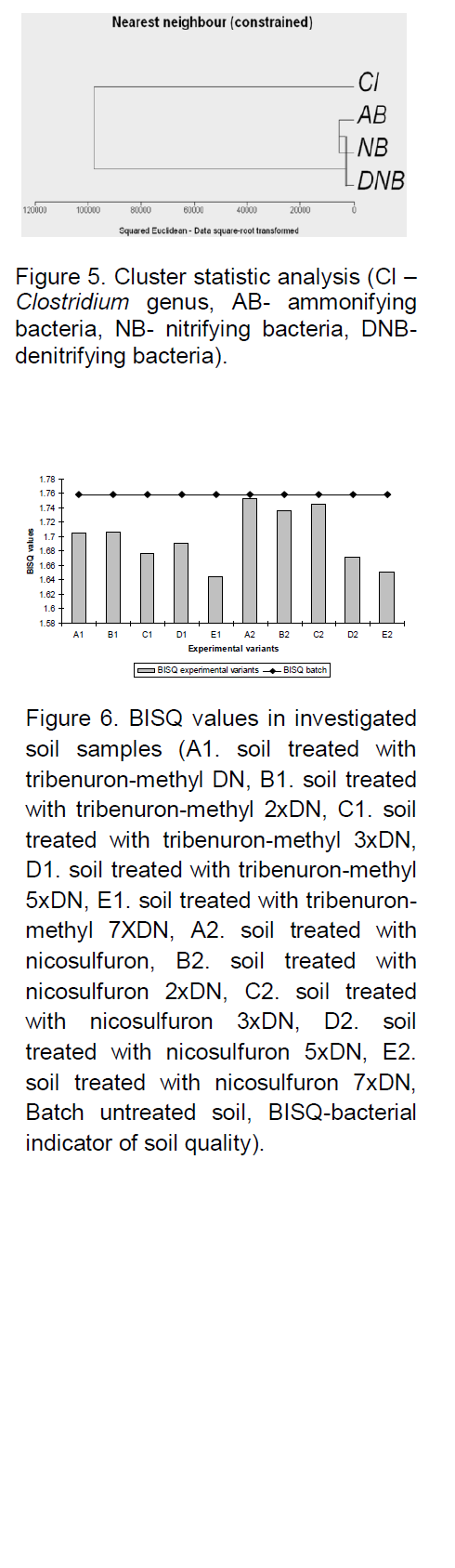
compared to AMB, NB, and DNB. Thus, CA graphical representation revealed that highest dissimilarity between the herbicide type and exposure dose for these bacteria (0.000 to 0.109), whereas AMB and NB displayed similar responses to tribenuron-methyl and nicosulfuron exposure, irrespective of exposure dose: AMB: 0.320 to 0.646; NB: 0.290 to 0.782 (Figure 4). This observation is supported by the results of cluster analysis (Figure 5).
The bacterial indicator of soil quality (BISQ) reflects the effects of tribenuron-methyl and nicosulfuron on soil bacterial biota. Bacterial indicator of soil quality recorded values between 1.645 to 1.706 ??for soils treated with tribenuron-methyl and 1.651 to 1753 in those treated with nicosulfuron. In our study, BISQ values were lower for nicosulfuron (range: 1.645 to 1.706) than those reported for tribenuron-methyl (range: 1.651 to 1.753).
This suggests that tribenuron-methyl is more toxic than nicosulfuron for bacterial communities that are involved in nitrogen cycle in soil (Figure 6).
The effects of nicosulfuron on soil bacteria enzymatic activity are lower as compared to tribenuron-methyl. The actual and potential dehydrogenase activities, as well as urease activity are essential for assessing the effects of xenobiotics substances on metabolic activity of soil microbial biota. Tribenuron-methyl and nicosulfuron induce a dose-dependent inhibition of enzymatic activity of ammonifying, nitrifying and denitrifying bacteria in soil. Tribenuron-methyl and nicosulfuron affect negative bacterial metabolic activity and bacterial communities.
The authors have not declared any conflict of interests.
REFERENCES
Accinelli C, Screpanti C, Dinelli G, Vicari A (2002). Short-time effects of pure and formulated herbicides on soil microbial activity and biomass. Int. J. Environ. Analyt. Chem. 82:519-527.
Crossref |
|
|
Accinelli C, Koskinen WC, Becker JM, Sadowsky MJ (2007). Environmental fate of two sulfonamide antimicrobial agents in soil. J. Agric. Food Chem. 55: 2677-2682.
Crossref |
|
|
Allievi L, Gigliotti C (2001). Response of the bacteria and fungi of two soils to the sulfonylurea herbicide cinosulfuron. Part B: Pesticides, Food Contaminants, and Agricultural Wastes. J. Environ. Sci. Health 36(2):161-175.
Crossref |
|
|
Andersen SM, Hertz PB, Holst T, Bossi R, Jacobsen CS (2001). Mineralisation studies of 14C-labelled metsulfuron-methyl, tribenuron-methyl, chlorsulfuron and thifensulfuron-methyl in one Danish soil and groundwater sediment profile. Chemosphere, 45(6-7):775-782.
Crossref |
|
|
Araújo ASF, Monteiro RTR, Abarkeli RB (2003). Effect of glyphosate on the microbial activity of two Brazilian soils. Chemosphere 52:799-804.
Crossref |
|
|
Atlas RM, Parmer D, Partha R (1978). Assessment of pesticide effects on non-target soil microorganisms. Soil Biol. Biochem. 10:231-239.
Crossref |
|
|
Avidano L, Gamalero E, Cossa GP, Carraro E (2005). Characterization of soil health in an Italian polluted site by using microorganisms as bioindicators. Appl. Soil Ecol. 30:21-33.
Crossref |
|
|
Benitez E, Melgar R, Nogales R (2004). Estimating soil resilience to a toxic organic waste by measuring enzyme activities. Soil Biol. Biochem. 36:1615-1623.
Crossref |
|
|
|
Borozan AB, Filimon MN, Misca CD, Bordean D, Darlea A, Dogaru D (2010). Prokaryotes and eukaryotes microorganisms type with some important agricultural chemical resistant. J. Agroaliment. Proc. Technol. 16(3):387-389. |
|
|
Casida LEJr, Klein DA, Santoro T (1964). Soil dehidrogenase activity. Soil Sci. 98:371-376.
Crossref |
|
|
|
Cusa V (1996). Instrucţiuni metodologice pentru analiza microbiologica a sedimentelor din ecosistemele acvatice. Instit. Cercet. Ing. Mediului Bucuresti 4:14-20. |
|
|
Cycon M, Piotrowska-Seget Z, Kozdroj J (2010). Response of indigenous microorganisms to a fungicidal mixture of mancozeb and dimethomorph added to sandy soils. Int. Biodeterorat. Biodegradat. 64:316-323.
Crossref |
|
|
|
Dunca S, Ailiesei O, Nimitan E, Stefan M (2007). Microbiologie aplicata. Editura Demiurg Iasi. |
|
|
Ghani A, Wardle DA (2001). Fate of 14C from glucose and the herbicide metsulfuron-methyl in a plant–soil microcosm system. Soil Biol. Biochem. 33:777-785.
Crossref |
|
|
Gimsing AL, Borggaard OK, Jacobsen OS, Aamand J, Sørensen J (2004). Chemical and microbiological soil characteristics controlling glyphosate mineralisation in Danish surface soils. Appl. Soil Ecol. 27:233-242.
Crossref |
|
|
|
Gigliotti C, Allievi L (2001). Differential effects of the herbicides bensulfuron and cinosulfuron on soil. J. Environ. Sci. Health Part B: Pesticides, Food Contamin. Agric. Wastes Microorg. 36(6):775-782. |
|
|
Gülser F, ErdoÄŸan E (2008). The effects of heavy metal pollution on enzyme activities and basal soil respiration of roadside soils. Environ. Monit. Assess. 145:127-133.
Crossref |
|
|
Ismail BS, Yapp KF, Omar O (1998). Effects of metsulfuron-methyl on amylase, urease, and protease activities in two soils. Austr. J. Soil Res. 36:449-456.
Crossref |
|
|
Latijnhouwers M, de Wit PJ, Govers F (2000). Oomycetes and fungi: similar weaponry to attack plants. Trends Microbiol. 11:462-469.
Crossref |
|
|
Lupwayi NZ, Hanson KG, Harker KN, Blackshaw RE, O'Donovan GWC, Johnson JT, Gan EN, Irvine Y, Monreal MA (2007). Soil microbial biomass, functional diversity and enzyme activity in glyphosate-resistant wheat-canola rotations under low-disturbance direct seeding and conventional tillage. Soil Biol. Biochem. 39:1418-1427.
Crossref |
|
|
Merini LJ, CuadradoV, Flocco CG, Giulietti AM (2007). Dissipation of 2,4–D in soils of the Humid Pampa region, Argentina: a microcosm study. Chemosphere 68:259-265.
Crossref |
|
|
|
Muntean V (1995-1996). Bacterial indicator of mud quality. Inst. de Cercet Biologice. Contributii Botanice. pp. 73-76. |
|
|
Muntean V, Crisan R, Pasca D (1996). Enzimological classification of salt lakes in Romania, Netherlands: Kluwer Academic Publishers. Int. J. Salt Lake Res. 5:35-44.
Crossref |
|
|
Omar SA, Abdel-Sater MA (2001). Microbial Populations and Enzyme Activities in Soil Treated with Pesticides. Water, Air Soil Pollut. 127(1-4):49-63.
Crossref |
|
|
Pampulha ME, Oliveira A, (2006). Impact of an Herbicide Combination of Bromoxynil and Prosulfuron on Soil Microorganisms. Curr. Microbiol. 53(3):238-243.
Crossref |
|
|
Saeki M, Toyota K (2004). Effect of bensulfuron-methyl (a sulfonylurea herbicide) on the soil bacterial community of a paddy soil microcosm. Biol. Fertil. Soils 40(2):110-118.
Crossref |
|
|
Sannino F, Gianfreda L (2001). Pesticides influence on soil enzymatic activities. Chemosphere 45:417-425.
Crossref |
|
|
Sarmah AK, Sabadie J (2002). Hydrolysis of Sulfonylurea Herbicides in Soils and Aqueous Solutions: A Review. J. Agric. Food Chem. 50(22):6253-6265.
Crossref |
|
|
Shaw LJ, Burns RG (2004). Enhanced mineralization of (U-14C) 2,4- dichlorophenoxyacetic acid in soil from the rhizosphere of Trifolium pretense. Appl. Environ. Microb. 70:4766-4774.
Crossref |
|
|
van Eerd LL, Hoagland RE, Zablotowicz RM, Hall JC (2003). Pesticide metabolism in plants and microorganisms. Weed Sci. 51:472-495.
Crossref |
|
|
Wang H, Xu J, Yates SR, Zhang J, Gan J, Ma J, Wu J, Xuan R (2010). Mineralization of metsulfuron-methyl in Chinese paddy soils. Chemosphere 78(3):335-341.
Crossref |
|
|
Yao X, Min H, Lü Z, Yuan H (2006). Influence of acetamiprid on soil enzymatic activities and respiration. Europ. J. Soil Biol. 42:120-126.
Crossref |
|
|
Zabaloy MC, Gómez MA (2008). Microbial respiration in soils of the Argentine Pampas after metsulfuron-methyl, 2,4–D and glyphosate treatments. Commun. Soil Sci. Plant Anal. 39:370-385.
Crossref |
|
|
Zabaloy MC, Garland JL, Gómez MA (2010). Assessment of the impact of 2,4- dichlorophenoxyacetic acid (2,4-D) on indigenous herbicide-degrading bacteria and microbial community function in an agricultural soil. Appl. Soil Ecol. 46:240-246.
Crossref |
|
|
|
Zarnea Gh (1994). Degradarea microbiana a substantelor xenobiotice. Tratat de microbiologie generala. Ed. Academiei Romane. pp. 976-991. |
|
|
Zhou X, Wang Y, Li W (2009). Effect of probiotic on larvae shrimp (Penaeus vannamei) based on water quality, survival rate and digestive enzyme activities. Aquaculture 287:349-353.
Crossref |