Full Length Research Paper

ABSTRACT
Chickpea is a major legume grown in Kenya. The agronomic performance, nodulation ability and grain crude protein are not fully exploited. Field experiments were conducted at Embu, Kenya to determine the effect of plant spacing and N-fertilizer rates on nodulation and protein content of selected chickpea varieties. A split-split plot design was used and replicated three times. Treatments consisted of main factor, variety in four levels (Saina K, Mwanza 2, Chania I, Chania III), sub factor, spacing in three levels (S1: 50 x 10; S2: 50 x 20; S3: 50 x 30 cm), and sub-sub factor, nitrogen fertilizer rates in four levels of (0; 30; 60; 90 kg ha-1). Data on nodulation and CP was subjected to GLM. Spacing and N-fertilizer rates significantly affected nodule numbers, fresh weight and their sizes. Mwanza 2 x 50x10 cm x N30 posted highest nodule numbers while Chania III had highest protein content. Interactions of VS, VN, SN and VSN had no consistent pattern under nodule factors but were highly significant in percent CP. Hence, optimal interactive effects of VSN can improve nodulation factors and CP of chickpea. The information will help growers make informed decisions, and researchers in developing improved varieties.
Key words: Chickpea, variety, spacing, N-fertilizer, nodulation, crude protein
INTRODUCTION
Chickpea (Cicer arietinum L.) is a legume crop belonging to fabaceae family. Globally, chickpea is ranked third, after common bean (Phaseolus vulgaris L.) and field pea (Pisum sativum L.) (FAO, 2019). The crop is drought tolerant and can survive under scanty to moderate rainfall conditions (Beyene et al., 2015) with an optimum precipitation of 152-254 mm and temperatures of 18-290 C (McVay and Crutcher, 2016). Considering climate change realities of extreme weather conditions, chickpea is likely to be an important grain legume for smallholder rural communities.
As a grain legume, chickpea has numerous nutritional benefits. It is a source of dietary fiber, polyunsaturated fatty acids, vitamins A, C and E, folate, magnesium, potassium, and iron (Wallace et al., 2016). In addition, chickpea contains phenolic compounds that render health benefits beyond basic nutrition through prevention of cardiovascular diseases (De Camargo et al., 2019). Moreover, according to Hossain et al. (2016), the crop plays a significant role in enhancing soil fertility through biological nitrogen fixation (BNF) of up to 140kg N ha-1 from air. In addition, the crop has ability to bio-control grassy weeds in farms (Rao et al., 2014) and is good for rotation crop as it helps in disease and pest control (Reen et al., 2014).
Major world producing regions of chickpeas include Asia, Europe, America and Australia (Beyene et al., 2015). Although, the production of chickpeas has steadily increased since 1961 to about 11.5 million tons per annum, over 1.3 million tons enter the world market yearly to supplement the deficit estimated at 12.7 million tons (Merga and Haji, 2019). Main chickpea growing areas in Africa are parts of North, West and Eastern Africa (Maya and Maphosa, 2020), that contributes about 63% of the total production (Abebe and Debebe, 2020). In Kenya, the crop ranks fourth after common bean, pigeon peas and cowpeas (FAO, 2015; Fao and Unicef, 2015). However, the production is dominant in drier areas of the Rift Valley and Eastern Kenya (Mallu, 2015) having an average yield of 3 tonnes ha-1 per annum (FAO, 2019). However, according to Monyo et al. (2015), on farm yield of chickpea varieties in Kenya range between 1200 to 1500 kg ha-1, which is 99% lower than the global average of 850 tonnes ha-1 (Muehlbauer and Sarker, 2017).
Conventionally, tropical soils have high mineralization rate that affects loss of soil nitrogen (N) during harsh environmental conditions, such as high temperatures, drought and soil acidity (Hungria and Vargas, 2000). There is need to incorporate crops that can enrich the soils with N-fixation for improved production and productivity (Giller et al., 2013). Legume formulation and nitrogen fixation depend on the balance between the supply of mineral N in the soil and the N requirement of the legume (Zhou et al., 2011). In the presence of effective rhizobia in the soil, the legume crop will fix N if there is less availability of mineral N in the soil than the plant requires. Nonetheless, if soil management affects the N requirement of the legume, nodulation and BNF will also be affected (Karkanis et al., 2016).
The presence of effective rhizobia in the soil is a condition for efficient legume-rhizobia symbiosis to deliver high N to the host plant and to enrich the soil N for the preceded crops (Choudhry, 2012). Thus, low crop productivity in sub-Saharan Africa (SSA), pronounced in grain legumes are often associated with declining soil fertility and reduced N2-fixation due to biological and environmental factors (Chianu et al., 2011). Historically, legume crops have played an important role in soil fertility through BNF, enhancement of soil organic matter, prevention of nutrient leaching and key process in sustainable land management (Kimutai, 2017; Kouyaté et al., 2012; Niste et al., 2013). In spite of chickpeas being the most important grain legume in tropical and sub-tropical countries (Jukanti et al., 2012), information on chickpea nodule development is quite limited (Mandal and Sinharoy, 2019).
Low chickpea production in Kenya is attributed to poor nodulation caused by improper varietal selection and poor agronomic practices, such as spacing and poor soil fertility (MoA, 2013; Lemma et al., 2013). Previous studies show that among other factors Variety (V), spacing (S) and N-fertilizer (N) are the main determinants of low nodulation (Bejandi et al., 2012; Liu et al., 2011) since they affect root temperatures and nitrogen assimilation. The higher root temperatures upset the formation of bacteroid tissue and hasten its degeneration while low temperatures retard nodule development. Common varieties are Chaina (Desi 1), ICCV 00108 (Desi), Saina K 1, ICCV 00305 (Kabuli), ICCV 92944 (Desi) and ICCV 97126 (Desi) (Kimurto et al., 2014; Rao et al., 2012). This study therefore, aimed to determine most appropriate interactions under study where high numbers, larger sizes and effective root nodules are formed to advise growers accordingly.
According to various findings, chickpea is an important grain having crude protein (CP) content of Desi and Kabuli chickpea cultivars range from 18 to 31%, being higher in Desi compared to the Kabuli types (Porkka et al., 2013). Further still, preceding works indicates that CP content of commonly grown chickpea varieties ranges between 17 to 24% CP (Mallu et al., 2014). However, other studies have shown that available soil N may influence the average CP concentration ranging from 18.3 to 23.2% (Singh et al., 2015). Studies in Turkey revealed a cumulative variance of between 82.7 and 90.5% for CP and total nitrogen (Sreerama et al., 2012). However, there is limited report on the grain protein content for the selected varieties for the proposed site of study. This study therefore, aimed to determine optimum interactive effects of variety, spacing and N application on optimum nodulation and crude protein content of selected chickpeas varieties in Embu, Kenya.
MATERIALS AND METHODS
Site description
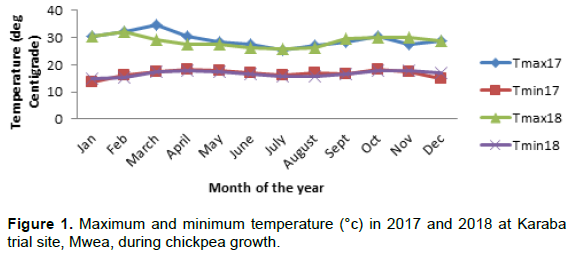
Field experiments were conducted in Mbeere South sub county of Embu County, Kenya. The study site was located at Karaba, Latitude 0°46’14.4822’S and Longitude 37° 22’23.79324’E with an altitude of 980 m above sea level. The site lies under lower midland (LM4) agro-ecological zone, receiving a bimodal annual rainfall amount of 600-700 mm (Jaetzold et al., 2006). The long rains (LR) are received from mid-March to June and short rains (SR) from mid-October to December and therefore has two cropping seasons per year. The average minimum temperatures (Tmin) range between 18 and 20°C; while maximum temperature (Tmax) ranges between 19-24°C (McVay and Crutcher, 2016). The site has black cotton soils and most farmers practice conventional tillage, with limited use of inorganic fertilizers. Since, the seasonal rainfall amounts are low; farmers prefer drought tolerant crops such as pigeon peas, cowpeas, chickpeas and sorghum. Livestock farming is also practiced with many farmers rearing local breeds (Zebu).
Weather data
Data on weather parameters, mean daily maximum and minimum temperatures, relative humidity and cumulative monthly precipitations were obtained from Mwea Irrigation Agricultural Development (MIAD) weather station (KMD, 2017) (Figures 1-3).


Soil sampling
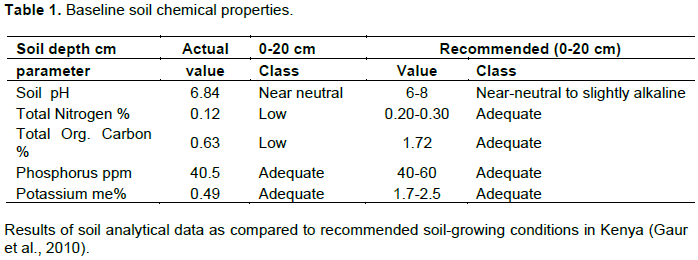
Baseline and endline soil samples were collected in March 2017 and August 2018 using an Eijkelkamp Gouge auger (Eijkelkamp Agrisearch Equipment, Giesbeek, and The Netherlands). Ten soil samples were randomly taken at 0 to 15 cm depth across the experimental plots. The samples were then packed in labeled Ziplock bags and transported to University of Embu Physical Science Laboratory for chemical analysis. In the laboratory, the soil samples were sieved through 2-mm sieve for analysis. For soil pH, soil and water were mixed in the 1:2.5 soil: water ratio and analyzed using glass probe pH meter. Soil carbon, nitrogen, potassium and phosphorous concentration were then analyzed using standard procedure as described by Okalebo et al. (2002) (Table 1).
Experimental design and layout
The field experiment was laid out in a split-split design of a 4x3x4 factorial arrangement with three replicates. The main plot was varieties while the sub plots were spacing and the sub-sub plots were the inorganic fertilizer. The plots dimensions were 3 x 3 m separated by 1m between plots and 2 m between replicates, with six (6) rows of chickpea variety and 30, 15 and 10 seed holes per row based on the inter-row spacing of 10, 20 and 30 cm.
Land preparation
Bush clearing was done by hand using a slasher. Ploughing was done once by tractor mounted with a disc plough. This was followed by harrowing using a disc harrow. Levelling was done manually using a hand hoe then plots demarcated using a tape measure and ropes. Marking of plots was done according to treatments once before each planting. In the consecutive seasons, hand hoes were used to prepare plots before planting.
Farm inputs
Chickpea seeds were sourced from Kenya Seed Company, Nakuru. Four varieties were selected, Kabuli (Saina K and Mwanza 2) and Desi (Chania I and Chania III). The varieties were selected because they are locally planted in Kenya and East Africa, are drought tolerant and have a degree of resistance to Fusarium wilt (Fusarium oxysporum). Di-ammonium phosphate (DAP) comprising of N: P: K in the ratio of 18:46: 0 was used to supply Nitrogen (N) nutrient. The choice of fertilizer was determined by farmers’ practice and the recommendations by the Ministry of Agriculture (Karugia et al., 2012).
Planting and management of crops
Two seeds were directly, sown into the planting holes according to the spacing of [S1 (50 x 10), S2 (50 x 20) and S3 (50 x 30) cm]. Thinning of plants was done 2 weeks after emergence to reduce competition among plants. Total plant population per plot of 9 m2 was 180, 90 and 60 for S1, S2 and S3, respectively. Planting was done in 3 seasons in the months of April 2017, October 2017 and April 2018. N-fertilizer was applied at planting time, using four rates of Zero (0 kg), 30kg, 60 kg and 90 kg ha-1 to minimize weather constraints that could reduce its uptake during growth. Weedall, a pre-emergence herbicide and sumithion pesticide was applied immediately after sowing to control weeds and crickets correspondingly. Weeding was done at three and 8 weeks after planting using a hand hoe. Collection of data was done up to 50% physiological maturity of the crop. Grains were harvested when the crop was completely dry, when 90% of the leaves have dropped on the ground.
Sampling unit
In each plot, sampling was done within a net plot measuring 1.5m x 2m. Three plants were selected randomly within the net plot for nodulation assessment. Data was collected on plant growth attributes (number of nodules, types of nodules and nodule fresh weight) for 3 consecutive seasons long rains in 2017 (LR17), long rains 2018 (LR18) and short rains in 2017(SR17).
Assessment of root nodules
At 50 % physiological maturity, three plants from each experimental plot were randomly dug out from within the sampling unit. The roots were gently cleaned in water to remove any adhering soil particles and nodules plucked off. Through observation, they were categorized into small, medium and large nodules, counted and an average recorded. Categorization of nodules were important as nodule size plays a significant role in BNF. The percentage of fixing and non fixing nodules were determined by observing colour of root nodules.
Determination of protein in grains using Modern Kjeldahl method
The method involved catalytically digesting organic material in a boiling mixture of Sulfuric acid and Sulfate salt at temperatures above 400 °C. During the process, the organically bonded nitrogen was converted into ammonium sulfate. Alkalizing the digested solution liberated ammonia, which was quantitatively steam distilled and N was determined by titration (Bicsak et al., 1993). The titrant used was diluted H2SO4 (0.2N) solution until the colour changed from green to pink (end- point) (Association of Official Analytical Chemists (AOAC) method 981 01 international).
Hence, percentage of nitrogen (% N) in 100 g sample was calculated according to equation 1.

Where Vs and Vb are the titration volumes of the sample and blank, and N is normality of H2SO4 (as found with titration with H2SO4), 14=equivalent weight of N in g, a= volume of digest taken for distillation in ml, b=weight of sample taken for analysis (I g). Using an appropriate conversion factor (F), 6.25 according to KJeldahl, method (Bremner, 1960), N was converted to crude protein (CP) using equation 2.
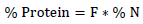
Data analysis
Data on nodulation and CP content was analyzed using Generalized Linear Model (GLM) according to equation 3 where numerical variables were converted to standard units.

Where, Yijkln = Number of nodules per plant; μ = General mean number of nodules experiment; Ri = Effect due to ith replication; Vj = Effect due to jth variety; Sk =Effect due to kth spacing; Nl=Effect due to lth N-levels; V SNjlk =Interaction effect between j, k, l for variety, spacing and Nitrogen levels; ?ijkl= Residual effect (random error). The means were separated by Tukeys honest significant difference (HSD) test for normally distributed data at 95% confidence level. All analyses were conducted in SAS 9.4 (SAS, 2015; Rutherford, 2011). Simple linear correlation analysis was also done to determine the association of various agronomic variables.
RESULTS
Assessment of root nodules
Nodule numbers and fresh weight
The selected chickpea varieties showed significant differences in number of nodules for all the 3 seasons. The average number of nodules recorded for the three seasons were 5.2, 6.7, 9.1 and 11.1 for varieties, Chania III, Chania I, Saina K and Mwanza 2 respectively (Table 2). The highest number of nodules per season were observed under Mwanza 2 (16.4) which was statistically similar to Saina K (15.58) during LR18 while the lowest was for Chania III (4.0) in SR17 (Table 2). The Mwanza 2 variety consistently had higher nodule numbers for the three seasons.

Spacing had a significant effect on the number of nodules in LR17 (p=0.0004) and SR17 (p<0.0001), with nodule averages of 14.5, 8.8 and 7.4 for 50 x 10, 50 x 20 and 50 x 30 cm spacing correspondingly. The 50x30 cm spacing recorded lowest nodules of 4.3 over LR17 and highest (13.1) over LR18. However, 50x20 cm and 50x10cm had statistically similar and superior nodules across all seasons that ranged from 6.2 to 12.2 nodules per plant.
The distribution of nodule numbers by N-fertilizer rates gave averages of 10.5, 9.0, 6.4 and 6.1 for N30, N0, N60 and N90 across the three seasons. The highest nodule number was under N30 (11.5) while the lowest was under N90 (1.9) during the SR17. The N30 and N0, application rates had significantly higher number of nodules in LR17 (p=0.0001) and SR17 (p<0.0001). During the LR18 season, N-fertilizer rates had insignificant effects on nodule numbers (Table 2). There was a significant interaction between Variety and N-rates in SR17 (p=0.0014). Other interactions posted non-significant effects on nodule numbers (Table 2).
The effect of variety on nodule fresh weight was significant at P=0.0006 (LR17), P=0.0002 (SR17) and P<0.001 (LR18). Kabuli varieties (Saina K and Mwanza 2) had nearly 100% more nodule fresh weight compared to the Desi varieties (Chania I and III). The lowest nodule fresh weight was 0.06g in Chania III (SR17) while the highest nodule fresh weight was 0.25g noted under Mwanza 2 although it was statistically similar to Saina K in SR17 and LR18 (Table 2).
Intra-row spacing was not significant on nodule fresh weight in the LR17 and SR17, but was significant in LR18 (P=0.0027). Lowest fresh weight was observed in the 50x10cm spacing (0.10 g) in the SR17 and the highest recorded in the 50x30 cm (0.21 g) in LR18 (Table 2). Similarly, N-fertilizer rates had no significant effect on nodule fresh weight in LR17 and SR17, while it was significant in LR18 (P=0.0018). The lowest fresh weight of 0.12g was seen under N90 N-application but statistically similar with N60 (0.14 g). Highest nodule fresh weight was seen under N30 (0.22 g) and control (0.16 g) which were statistically not significant from each other (Table 2).
Distribution of nodule sizes
Distribution of the number of nodule sizes in the 3 classes: small, medium and large impacted by variety was significantly different across all seasons (Table 3). Averages nodule numbers per size class among the chickpea varieties were 1.7, 2.0, 1.9 and 1.5 (small), 3.6, 4.7, 2.8 and 2.4 (medium), 3.8, 4.5, 1.9 and 1.2 (large) for Saina K, Mwanza 2, Chania I and Chania III varieties respectively. Noteworthy differences were in SR17 (P<0.0001) (small), P=0.0076 (LR17) and P=0.0005 (SR17) (medium) and P=0.0002 (LR17), P=0.0007 (SR17) and P<0.0001 (LR18) (large). Highest nodule numbers per size class were 2.3 (small) for Mwanza 2 in SR17, 6.8 (medium) for Chania I in LR18, and 8.1 (large) for Mwanza 2 in LR18.
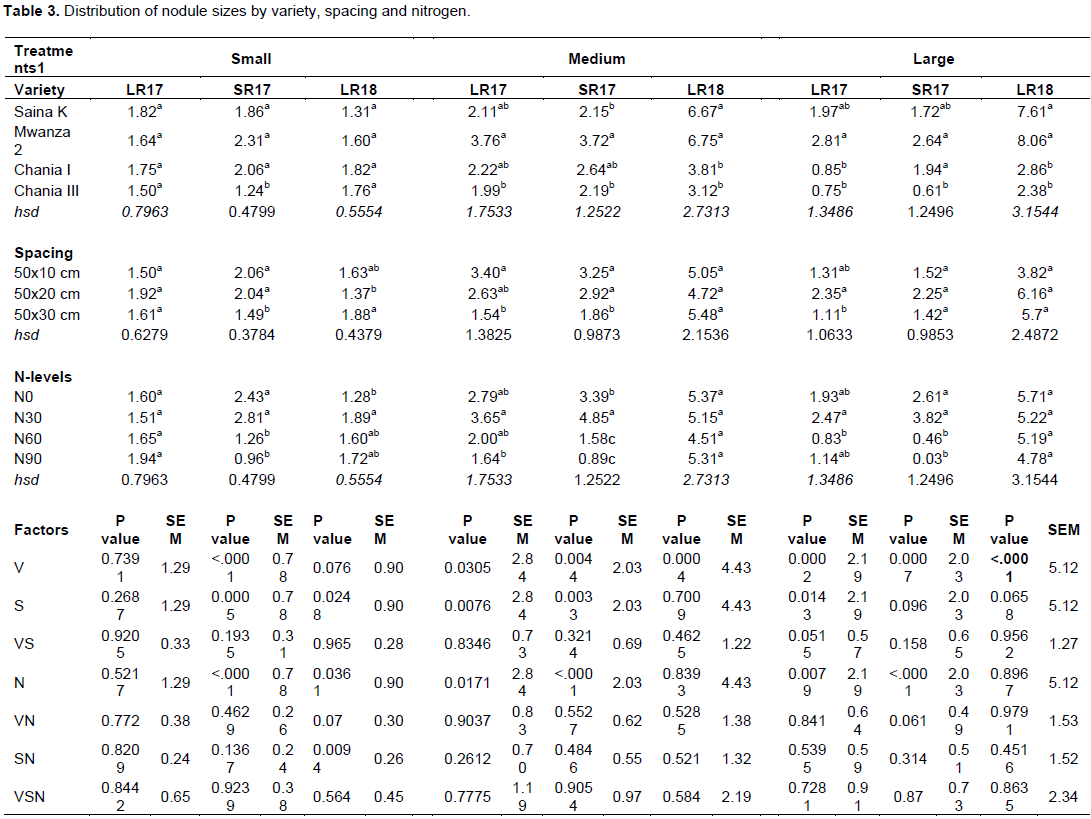

Distribution of nodule numbers according to spacing was 1.7, 1.8, and 1.7 (small nodules), 4.2, 3.7 and 3.5 (medium nodules), 2.2, 3.6 and 2.7 (large nodules) for he 50x10, 50x20 and 50x30cm spacing respectively. Effect of spacing on numbers of nodule per size class was significant at P=0.0005 (SR17) and 0.0248 (LR18) for small sizes, P=0.0076(LR17) and P=0.0033 (SR17) for medium sizes and P = 0.0143 (LR17) for large sizes. Highest number of nodules per size class under spacing was 2.1 at 50x10 cm for small nodules, 3.4 at 50x10 cm for medium nodules, and 6.2 at 50x20 for large nodules (Table 3).
Application of nitrogen had a significant effect on the number of small nodules formation in SR17 (P<0.0001) and LR18 (P=0.0361), medium nodules in LR17 (P=0.0171) and SR17 (P<0.0001) and large nodule numbers in LR17 (P =0.0079) and SR17 (P<0.0001). Highest nodule size numbers were 2.81 under N30 (SR17) and 3.65 at N30 (LR17) while lowest was 1.0 (N90) and 0.9 (N90) for small and medium sizes correspondingly. Major interactions were noted for large nodules between VS (P=0.0515) over LR17, 0.0002 (Table 3).
Colour of nodules and effective nodules
The colour of nodules defines the ability of nodules to fix (effective) or not to fix (non-effective) nitrogen. The average number of nodules recorded were 5.3, 6.7, 4.0, 2.9 (pink nodules) and 0.2, 0.2, 0.1, 0.1 (yellow nodules) for Saina K, Mwanza 2, Chania I and Chania III correspondingly. Chickpea variety had a significant effect on nodule colour was at (P<0.0001) and P=0.0010 (pink), P=0.0078 and P<0.0001 (yellow) (Table 5). The highest number of pink nodules was 8.89 for Mwanza 2 (LR18) while the lowest was 2.31, Chania III (SR17). Yellow nodules were highest for Saina K and Mwanza 2 (0.2) and lowest for Chania I and III (0.1) singly (Table 4).


Nodule colouration as influenced by spacing showed mean values of 4.5 (50x10, 30 cm), 5.1 (50x20 cm) and 0.1 (50x10 cm), 0.2 (50x20, 30 cm) for pink and yellow respectively. Pink nodules were significant (P=0.0047, P=0.0005, P=0.0103) while yellow-green nodules were significant (P=0.0002, P=0.0010) over LR17 and SR17. Highest pink nodules were 7.39 over LR18, while lowest was 2.84 (LR17) at 50x30 cm. Highest yellow-green nodules were 5.80 (LR18) under 50x20 cm while lowest was 1.47 (LR17) under 50x30 cm spacing (Table 4).
Effect of N rates on number of pink nodule means were 5.3, 6.3, 3.7 and 3.5 under N0, N30, N60, and N90 respectively. Yellow nodule averages were 0.2 (N0, N30) and 0.1 (N60, N90). Highest pink nodules were 7.58 (N30) in SR17 season, while the lowest was 1.10 (N90)-SR17. Highest yellow-green were 3.89 (N30) and lowest 0.78 (N90) over SR17. LR18 resulted in insignificant effects of fertilizer on both pink and yellow green nodules (Table 4).
Significant interactions for pink nodules were observed VN (P<0.0001) over SR17 and SN (P=0.0296) over LR18 (Table 4). The percentage of effective nodules showed insignificant results (Table 4).
Grain crude protein levels
The CP of the chickpea varieties was determined and presented in Table 5. Significant differences were noted among the different varieties with Chania III producing grains with the highest CP in all the three seasons compared to the other varieties whose crude protein was statistically similar. The CP values ranged from 21.57% for Mwanza 2 under SR17 to 26.91% for Chania III in the same season. Spacing had no effect on the CP in the three growing seasons. The CP values varied between 22.39% at 50x30 cm spacing to 24.86% at 50x20 cm spacing in the SR17 and LR17, correspondingly.

The nitrogen (N) fertilizer had no significant effect on the CP in the LR17 and LR18 seasons. In the SR17 season the 30 Kg-N/ha resulted in the highest CP compared to the 60 Kg-N/ha fertilizer rate. The CP values ranged from 22.45% to 26.03% for the 60 and 30 Kg-N/ha rates in the SR17 season. The effect of Variety was seen across all seasons at P<0.0001 (LR17), P=0.0002 (SR17) and P<0.0001 (LR18). Crude grain protein was highest among Chania III (26.91, 26.94 and 26.85 %) across all seasons of LR17, SR17 and LR18. The lowest % protein was seen under Mwanza 2 (21.57 %) which was statistically similar to Saina K and Chania I (Table 5. Insignificant effects of spacing on percent protein were observed across all seasons (Table 5). N-application was significant over SR17 at P=0.0190 where highest CP was illustrated under N30 (26.03 %) while the lowest was N60 (22.45 %) (Table 5). Major interactions were also noted under VS cross all seasons (P=0.0044, 0.0015 and 0.0002), VN at P=0.0010 (LR17), SN at P=0.0381 (SR17) and P=0.0348 (LR18) and VSN across all seasons (P=0.0005, 0.0262 and 0.0009) (Table 5).
DISCUSSION
Chickpea nodulation
Number of nodules and nodule fresh weight
Nitrogen has been most limiting nutrient in plants (Lori et al., 2018) and legumes will use N-fixing bacteria to counter N limitation (Fagodiya et al., 2020; Rondon et al., 2007). Root nodulation offers a symbiotic relationship between N-fixing bacteria and leguminous plant roots (Suzaki et al., 2015), commonly found in nitrogen-limited environments. Therefore, formation of well-established nodules is thus important for N-fixation. Differences in nodule numbers and nodule fresh weight among chickpea varieties could be due to competition for limited N. Similar results were observed by Girma (2015) who noted differences in numbers of nodules ranging from 20.92 and 10.94 among Kabuli and Desi chickpeas varieties respectively. In addition, Asante et al. (2020) and Roy et al. (2019) also reported the same.
Higher nodule numbers under smaller spacing (50 x 10 cm, 50x20 cm) is attributed to increased competition for N that activated higher nodulation in the roots. In wider spacing (50x30 cm), at the same level of fertilizer application, there was higher N available per plant reducing nodule formation. Concurring with the result is Merga (2020) who found low nodule numbers among common bean varieties under higher spacing of 60 cm. Other findings are of Sethi et al. (2015). Results were however, in contravention with findings of Thalji (2006) and Kumar et al., (2016) who found non-significant effects of spacing on nodules in faba beans and mung beans trials respectively.
Although nitrates play a significant role in plant growth, many findings have revealed the need for minimal use in leguminous crops to encourage nodule formation.
Legume crops overcome conditions of N limitation, during active growth by increasing nodule number per plant, nodule size, and the increase of nitrogenase activity (Hungria and Vargas, 2000) hence significant results at N0 and N30. N90 decreased nitrogenase activity of rhizobia bacteria, inhibiting root infection and nodule development. Namvar et al. (2011) echoed similar results in his study on chickpeas where application of 50 kg N ha-1 gave higher nodule numbers and nodule dry weight compared to the control and 100 kg N ha-1. Significant effect of N on nodule fresh weight during LR18 can be attributed to higher supply of nitrates due to adequacy in rainfall that improved growth of shoots and root nodules. This is in line with Tajer (2016); Weisany et al. (2013); Gan et al. (2010) who documented that high N fertilization suppress nodulation of pulses.
The effect of N depends on the interaction between roots and aboveground parts of a plant. Although, spacing and N affected nodule numbers and fresh weight, they had a positive correlation with amount of rainfall increasing over LR18 compared to LR17 and SR17 by over 77 and 11% for both correspondingly. Significant interactions were noted in VN and SN, In agreement is Erdemci et al. (2017) who noted similar interactions in chickpeas. Worku and Astatkie (2015) reported insignificant results of VS on soybean which is similar to this study.
Nodule size distribution
It has been proven that the activity of BNF of root nodules is closely associated with the nodule size and nodule mass (Tajima et al., 2007). Categorization of nodules in terms of size and abundance can help understand their contribution in plant growth (King and Purcell, 2001). Large nodules may help improve drought tolerance due to greater size of non-cortical tissue than in small nodules. Distribution of the nodule sizes in the three classes: small, medium and large impacted by variety was significantly different across all seasons. This is in agreement with Hazra et al. (2020) who found various sizes in chickpea cultivar JG 16. Besides, Flowers et al. (2010) documented similar outcomes. Smaller spacing, which gave higher numbers of plants per unit area, increased competition for N making it limiting. This prompted formation of many numbers of small and medium sizes of nodules due to minimal growth factors that reduced expansion of nodules.
Although high levels of Nitrate-N have no significant effect on both the initial formation and number of nodules (Xia et al., 2017), they distinctly reduce both nodule size and activity of rhizobia (David and Khan, 2001). Nodule sizes increased as N-application increased (N0 to N30) and then decreased with increase of N application across all sizes and seasons. The increase in number of all sizes on application of 30 kg ha-1 N is accredited to presence of mineral N, which on absorption is used for biological processes including root growth and expansion (Tajer, 2016). In agreement with the results, Isoi and Yoshida (1991) found differences in nodule sizes in common bean and soybean varieties ranging between 5 to 131.
Addition of nitrogen (N60 and N90) severely depressed nodule size numbers due to inhibitory activity of nitrogenase that reduces root infection and growth. In similarity with the findings were reports by Xia et al. (2017), who found reduced sizes of nodules under low N-concentrations (control) in soybeans. Thus, use of 30 kg N ha-1 is important for growth and enlargement of nodules. Important interactions, VS, were due to competition among chickpea plants for growth factors (nutrients, moisture, light and space). In line with the results is Chianu et al. (2011). However, Worku and Astatkie (2015) documented non-significant effects of VS on nodules per plant in soybeans. More notably is SN interactions for small nodules due to competition for N-nutrients that resulted in less growth and expansion of nodules. Other scientists, David and Khan, (2001) reported similar findings. Therefore, it is worth mentioning that decreased spacing and high N rates reduce nodule sizes.
Nodule colour distribution and effective nodules
Nodule colour in leguminous plants is important as it shows whether BNF will take place or not (Bergersen, 1982). The colour is associated to the type of bacteria in the root nodules, genetic ability of the crops or the type of nutrition under which the crop is exposed. Chickpea nodules can be red, brown, white or green in pigmentation depending on the level of leghemoglobin pigment. Active nodules are coloured inside (pink-red-brown) as a result of pigmented protein (Achakzai, 2007). Significant nodule colour variations among Kabuli and Desi varieties could also be attributed to interactions between the N and carbon (C) cycles that affect the crop’s N:C balance and nodulation (Kumar et al., 2020). This is in agreement with Chemining’wa et al., (2013) who determined active nodules in chickpea varieties. In similarity with the findings is Ibewiro et al. (2001). This study suggests that, aspect of competition of chickpea crops for available N resulted in increased root hairs that enhanced nodule formation, which stimulated nitrogen fixation. Interactions of VN on pink nodules agree with findings of Egamberdieva et al. (2017) that showed effective nodulation of chickpeas. This shows that root nodulation may prefer the uptake of mineral N for energy reasons shrinking rate of BNF. Besides, effective root nodulation and N fixation may not take place if the right rhizobia association is limited (Tena et al., 2016).
Grain crude protein levels
Chickpea plants are an important source of proteins for smallholder farmers in Kenya. Known concentrations of Desi and Kabuli chickpeas around the world have shown a range of 18 to 31%, with Desi varieties having the highest CP (Sharma et al., 2013). However, high CP content in Chania III could be attributed to differences in genetic traits that enhanced efficient translocation of N from biomass to seed. Similar results have previously, been noted by Gaur et al. (2016) who found high protein content of 29.2% in Desi compared to 20.5% CP in Kabuli. Other outcomes are of Zhao et al. (2008). Nevertheless, differences of varieties in protein percentage showed that the superior nodulating variety (Mwanza 2) did not translate to higher percent CP. Further, some findings also suggest non-functional nodules can be formed translating to low CP in grains (Kalefeto?lu et al., 2017).
While spacing posted statistically similar results of percent CP, Low spacing resulted in higher CP than low spacing by 2%. Increase of CP at low spacing could be due to decreased carbohydrates to protein ratio, because of interplant competition for growth factors that resulted in high nodulation and high N-fixing activity. The results are in agreement with Farjam et al. (2014) who found highest protein content of 24.1% in chickpea grains at low spacing of 20 cm compared to highest spacing of 30 cm (17%). Nonetheless, results of similarities in the CP content was ratified by Ashrafi et al. (2015) who found insignificant differences in CP in two cultivars, Bivanij and Azad at 25cm x 10cm of chickpeas. Statistical similarity indicates that findings of CP might not be majorly sensitive to management practices like spacing.
It is noteworthy that the crops’ ability to convert inorganic N into CP in the seed is more efficient when N application is enhanced. This however, is only possible if there is sufficient moisture before physiological maturity to increase nutrient uptake that leads to high concentrations of mineral N in seed grains. High CP at N90 approves findings of Bellaloui et al. (2020) who found high protein in soybeans when he used 168kg N ha-1 compared to control. Of importance to note is that while, all interactions VS, VN, SN and VSN were highly significant for CP, VS is the more feasible interaction due to its low error margins.
CONCLUSION
This study demonstrated that interactions of Variety, Spacing and Nitrogen rates have significant implications on nodulation and percent CP content of chickpea varieties, grown in Mbeere. Proper identification of the Variety, optimum Spacing and N-rates are feasible in improving nodulation and grain quality. Moreover, response of chickpea to N was significant suggesting low levels of soil N in the experimental site. Nodule numbers, nodule fresh weight and nodule colour showed Mwanza 2; 50x20 cm spacing; N30 to be superior. For protein content, Chania III, 50x10 cm and N30 was more yielding indicating that high nodulation does not translate to high protein content. Additionally, Sizes of root nodules agreed with the fresh weights in determining CP. Although Desi varieties have higher CP, farmer preference in the Kenyan context is Kabuli varieties due to their palatability.
RECOMMENDATIONS
Favourable interaction for variety, spacing and nitrogen is Mwanza 2, spacing 50x10 or 20 cm and N30 for enhanced nodulation and pink nodule colours. In terms of protein content, chania III, 50x10 cm and N30 will be more desirable. However, since majority of farmers in Kenya do not value quality but volumes, therefore, Mwanza 2 would be recommendable. Selection of varieties with high protein content and moderate seed size can be used in developing chickpea cultivars with high protein content and high grain yield.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interest.
REFERENCES
|
Abebe G, Debebe S (2020). Factors Affecting Chickpea Market Chain: The Case of Dembia District, North Gondar Zone, Ethiopia. Transportation P 66. |
|
|
Achakzai AK (2007). Effect of various levels of nitrogen fertilizer on nodulation of pea cultivars. Pakistan Journal of Botany 39(5):1673-1680. |
|
|
Asante M, Ahiabor BD, Atakora WK (2020). Growth, Nodulation, and Yield Responses of groundnut (Arachis hypogaea L.) as influenced by combined application of rhizobium inoculant and phosphorus in the Guinea Savanna zone of Ghana. International Journal of Agronomy 7 p. |
|
|
Ashrafi V, Pourbozorg H, Kor NM, Ajirloo AR, Shamsizadeh M, Shaaban M (2015). Study on Seed Protein and Protein Profile Pattern of Chickpea (Cice arietinum L.) by SDS-PAGE under Drought Stress and Fertilization. International Journal of Life Sciences 9(5):87-90. |
|
|
Bejandi TK, Sharifii RS, Sedghi M, Namvar A (2012). Effects of plant density, Rhizobium inoculation and microelements on nodulation, chlorophyll content and yield of chickpea (Cicer arietinum L.). Annals of Biological Research 3(2):951-958. |
|
|
Bellaloui N, McClure AM, Mengistu A, Abbas HK (2020). The Influence of Agricultural Practices, the Environment, and Cultivar Differences on Soybean Seed Protein, Oil, Sugars, and Amino Acids. Plants 9(3):378. |
|
|
Bergersen FJ (1982). Root nodules of legumes: Structure and functions. Root nodules of legumes. |
|
|
Beyene S, Worku W, Abate B, Endalkachew A, W/Meskel E, Assefa M, Hidoto L, Tena W, Dedefo T, Ayana R, Fikre A (2015). Chickpea (Cicer arietinum L.) production in the Southern Nations, Nationalities, and Peoples' Region of Ethiopia pp. 1-26. |
|
|
Bicsak RC, Boles R, Cathey R, Collins V, Hannasious K, Haselhorst J, Henderson L, Jann L, Meschi L, Molloy R, Stillions M, Swanson K, Tate D, Webb J, Wilkins G (1993). Comparison of Kjeldahl method for determination of crude protein in cereal grains and oilseeds with generic combustion method: collaborative study. Journal of AOAC international 76(4):780-786. |
|
|
Bremner JM (1960). Determination of nitrogen in soil by the Kjeldahl method. The Journal of Agricultural Science 55(1):11-33. |
|
|
Chemining'wa GN, Mwangi PW, Mburu MW, Mureithi JG (2013). Nitrogen fixation potential and residual effects of selected grain legumes in a Kenyan soil. International Journal of Agronomy and Agricultural Research 3(2):14-20. |
|
|
Chianu JN, Nkonya EM, Mairura FS, Chianu JN, Akinnifesi FK (2011). Biological nitrogen fixation and socioeconomic factors for legume production in Sub-Saharan Africa. In Sustainable Agriculture Volume 2. Springer, Dordrecht pp. 309-329. |
|
|
Choudhry M (2012). Growth, yield and nitrogen content of lentil (Lens culinaris medic) as affected by nitrogen and diquat application. Doctoral dissertation, University of Saskatchewan. |
|
|
David J, Khan KS (2001). Effect of nitrogen application on nodulation in inoculated chickpea (Cicer arietinum L.). Journal of Biological Sciences 1(3):87-89. |
|
|
De Camargo AC, Favero BT, Morzelle MC, Franchin M, Alvarez-Parrilla E, de la Rosa LA, Geraldi MV, Marostica Junior MR, Shahidi F, Schwember AR (2019). Is chickpea a potential substitute for soybean? Phenolic bioactives and potential health benefits. International Journal of Molecular Sciences 20(11):2644. |
|
|
Egamberdieva D, Wirth SJ, Shurigin VV, Hashem A, Abd_Allah EF (2017). Endophytic bacteria improve plant growth, symbiotic performance of chickpea (Cicer arietinum L.) and induce suppression of root rot caused by Fusarium solani under salt stress. Frontiers in Microbiology 8:1887. |
|
|
Erdemci I, Aktas H, Nadeem MA (2017). Effect of fertilization and seed size on nodulation, yield and yield components of Chickpea (Cicer Arietinum L.). Applied Ecology and Environmental Research 15(1):563-571. |
|
|
Fagodiya RK, Kumar A, Kumari S, Medhi K, Shabnam AA (2020). Role of nitrogen and its agricultural management in changing environment. In Contaminants in Agriculture. Springer, Cham., pp. 247-270. |
|
|
Food and Agriculture Organization (FAO) (2015). Status of the world's soil resources (SWSR)-main report. Food and Agriculture Organization of the United Nations and Intergovernmental technical panel on soils, Rome, Italy, 650. |
|
|
Food and Agriculture Organization (FAO) (2019). Food and Agriculture Organization of the United Nations statistic division. |
|
|
FAO, Unicef (2015). The state of food insecurity in the world 2012. Economic growth is necessary but not sufficient to accelerate reduction of hunger and malnutrition. Rome, FAO. P 46. |
|
|
Farjam S, Kenarsari MJ, Rokhzadi A, Yousefi B (2014). Effects of inter-row spacing and superabsorbent polymer application on yield and productivity of rainfed chickpea. Journal of Biodiversity and Environmental Sciences 5(3):316-20. |
|
|
Flowers TJ, Gaur PM, Gowda CL, Krishnamurthy L, Samineni S, Siddique KH, Turner NC, Vadez V, Varshney RK, Colmer TD (2010). Salt sensitivity in chickpea. Plant, Cell and Environment 33(4):490-509. |
|
|
Gaur PM, Singh MK, Samineni S, Sajja SB, Jukanti AK, Kamatam S, Varshney RK (2016). Inheritance of protein content and its relationships with seed size, grain yield and other traits in chickpea. Euphytica 209(1):253-260. |
|
|
Gaur PM, Tripathi S, Gowda CLL, Ranga Rao GV, Sharma HC, Pande S, Sharma M (2010). Chickpea Seed Production Manual. Patancheru 502 324, Andhra Pradesh, India: International Crops Research. Institute for the Semi-Arid Tropics. |
|
|
Gan Y, Johnston AM, Knight JD, McDonald C, Stevenson C (2010). Nitrogen dynamics of chickpea: Effects of cultivar choice, N fertilization, Rhizobium inoculation, and cropping systems. Canadian Journal of Plant Science 90(5):655-666. |
|
|
Giller KE, Schilt C, Franke AC (2013). N2Africa: putting nitrogen fixation to work for smallholder farmers in Africa. N2Africa project. No. 18. |
|
|
Girma A (2015). Yield, Yield Attributes, Nodulation and Protein Content of Chickpea as Influenced by Variety and Inoculation with Rhizobium Strains. Journal of Biology, Agriculture and Healthcare 5(17):115-120. |
|
|
Hazra KK, Nath CP, Singh SS, Swain DK, Kumar N, Das K, Lamichaney A (2020). Categorization of Chickpea Nodules and Their Relation with Plant Growth. National Academy Science Letters pp. 1-5. |
|
|
Hossain Z, Wang X, Hamel C, Knight JD, Morrison MJ, Gan Y (2016). Biological nitrogen fixation by pulse crops on semiarid Canadian prairies. Canadian Journal of Plant Science 97(1):119-131. |
|
|
Hungria M, Vargas MA (2000). Environmental factors affecting N2 fixation in grain legumes in the tropics, with an emphasis on Brazil. Field Crops Research 65(2-3):151-164. |
|
|
Ibewiro B, Onuh M, Sanginga N, Vanlauwe B, Merckx R (2001). Symbiotic performance of herbaceous legumes in tropical cover cropping systems. The Scientific World Journal 1:17-21. |
|
|
Isoi T, Yoshida S (1991). Low nitrogen fixation of common bean (Phaseolus vulgaris L.). Soil Science and Plant Nutrition 37(3):559-563. |
|
|
Jaetzold R, Schmidt H, Hornetz B, Shisanya C (2006). Farm management handbook of Kenya-Eastern Province. Ministry of Agriculture, Kenya/Deutsche Gesellschaft für Internationale Zusammenarbeit: Nairobi, Kenya. |
|
|
Jukanti AK, Gaur PM, Gowda CL, Chibbar RN (2012). Nutritional quality and health benefits of chickpea (Cicer arietinum L.): a review. British Journal of Nutrition 108(S1):11-26. |
|
|
Kalefeto?lu T, Macar O, Dürdane ?N (2017). Variability in some biochemical and nutritional characteristics in desi and Turkish kabuli chickpea (Cicer arietinum L.) types. Celal Bayar Üniversitesi Fen Bilimleri Dergisi, 13(3): 677-680. |
|
|
Karkanis A, Ntatsi G, Kontopoulou CK, Pristeri A, Bilalis D, Savvas D (2016). Field pea in European cropping systems: adaptability, biological nitrogen fixation and cultivation practices. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 44(2):325-336. |
|
|
Karugia J, Massawe S, Guthiga, P, Ogada M, Wambua, J (2012). Technologies for enhancing the productivity of cereals, pulses, roots and tubers in the arid and semiarid lands of Kenya. ReSAKSS-ECA Annual Trends and Outlook Report. |
|
|
Kenya Meteorological Department (KMD) (2017). KMD (2017/2018). Kenya Meteorological department. Weather forecast report |
|
|
Kimutai WJ (2017). Effect of selected bradyrhizobia and nutrients on cowpea biomass, biological nitrogen fixation and yield in Kilifi and Mbeere regions of Kenya. Doctoral dissertation, Kenyatta University. |
|
|
Kimurto PK, Towett B, Mulwa RS, Njogu N, Kosgei A, Songok S, Oyier M, Rutto D, Korir P, Rao G, Silim S (2014). An overview of chickpea breeding programs in Kenya. Legume Perspectives (3):55-57. |
|
|
King CA, Purcell LC (2001). Soybean nodule size and relationship to nitrogen fixation response to water deficit. Crop Science 41(4):1099- 1107. |
|
|
Kouyaté Z, Krasova-Wade T, Yattara II, Neyra M (2012). Effects of cropping system and cowpea variety on symbiotic potential and yields of cowpea (Vigna unguiculata L. Walp) and pearl millet (Pennisetum glaucum L.) in the Sudano-Sahelian Zone of Mali. International Journal of Agronomy, 2012. |
|
|
Kumar S, Meena RS, Datta R, Verma SK, Yadav GS, Pradhan G, Molaei A, Rahman GM, Mashuk HA (2020). Legumes for carbon and nitrogen cycling: an organic approach. In carbon and nitrogen cycling in soil. Springer, Singapore pp. 337-375. |
|
|
Kumar M, Panwar GS, Kushwaha S (2016). Effect of planting date, seed rate and row spacing on nodulation efficiency of bold seeded spring mungbean [Vigna radiata (L.) Wilczek] in Bundelkhand region of India. Legume Research-An International Journal 39(2):293-296. |
|
|
Lemma W, Wassie H, Sheleme B (2013). Response of chickpea (Cicer Arietinum L.) to nitrogen and phosphorus fertilizer S in Halaba and Taba, Southern Ethiopia. Ethiopian Journal of Natural Resources 13:115-128. |
|
|
Liu Y, Wu L, Baddeley JA, Watson CA (2011). Models of biological nitrogen fixation of legumes. Sustainable Agriculture 2:883-905. |
|
|
Lori M, Symanczik S, Mäder P, Efosa N, Jaenicke S, Buegger F, Tresch S, Goesmann A, Gattinger A (2018). Distinct nitrogen provisioning from organic amendments in soil as influenced by farming system and water regime. Frontiers in Environmental Science 6:40. |
|
|
Mallu TS (2015). Evaluation of Chickpea Genotypes for Yield and Selected Agronomic Traits in Kenya. Doctoral dissertation. |
|
|
Mallu TS, Mwangi SG, Nyende AB, Rao NVPRG, Odeny DA, Rathore A, Kumar A (2014). Assessment of genetic variation and heritability of agronomic traits in chickpea (Cicer arietinum L). International Journal of Agronomy and Agricultural Research 5(4):76-88. |
|
|
Mandal D, Sinharoy S (2019). A toolbox for nodule development studies in chickpea: a hairy-root transformation protocol and an efficient laboratory strain of Mesorhizobium sp. Molecular Plant-Microbe Interactions 32(4):367-378. |
|
|
Maya M, Maphosa M (2020). Current status of chickpea production: Opportunities for promoting, adoption and adapting the crop in Zimbabwe: A review. Journal of Dryland Agriculture 6(1):1-9. |
|
|
McVay KA, Crutcher F (2016). Chickpea Production. Environment 2011(4):5. |
|
|
Merga JT (2020). Evaluation of common bean varieties (Phaseolus vulgaris L.) to different row-spacing in Jimma, South Western Ethiopia. Heliyon 6(8):04822. |
|
|
Merga B, Haji J (2019). Economic importance of chickpea: Production, value, and world trade. Cogent Food and Agriculture 5(1):1615718. |
|
|
Ministry of Agriculture (MOA) (2013). Crops extension pocket handbook, Vol. 1: Field crops, Revised Edition pp. 93-117. |
|
|
Monyo E, Mponda O, Ganesamurthy K, Upadhyaya HD, Janila P (2015). Bulletin of Tropical Legumes. Tropical Legumes III, 2:01-06. |
|
|
Muehlbauer FJ, Sarker A (2017). Economic importance of chickpea: production, value, and world trade. In The chickpea genome. Springer, Cham. pp. 5-12. |
|
|
Namvar A, Sharifi RS, Sedghi M, Zakaria RA, Khandan T, Eskandarpour B (2011). Study on the effects of organic and inorganic nitrogen fertilizer on yield, yield components, and nodulation state of chickpea (Cicer arietinum L.). Communications in Soil Science and Plant Analysis 42(9):1097-1109. |
|
|
Niste M, Vidican R, Pop R, Rotar I (2013). Stress factors affecting symbiosis activity and nitrogen fixation by Rhizobium cultured in vitro. ProEnvironment 6(2013):42-45. |
|
|
Okalebo JR, Gathua KW, Woomer PL (2002). Laboratory methods of soil and plant analysis: a working manual second edition. Sacred Africa, Nairobi, 21. |
|
|
Porkka M, Kummu M, Siebert S, Varis O (2013). From food insufficiency towards trade dependency: a historical analysis of global food availability. PloS One 8(12):82714. |
|
|
Reen RA, Thompson JP, Clewett TG, Sheedy JG, Bell KL (2014). Yield response in chickpea varieties and wheat following crop rotations affecting population densities of Pratylenchus thornei and arbuscular mycorrhizal fungi. Crop and Pasture Science 65(5):428-441. |
|
|
Rao NV, Silim SN, Simtowe F, Gaur PM, Gowda CL, Monyo ES (2012). Enhancing Chickpea Productivity and Production in Eastern and Southern Africa. |
|
|
Rao AN, Wani SP, Ladha JK (2014). Weed management research in India-an analysis of past and outlook for future. |
|
|
Rondon MA, Lehmann J, Ramírez J, Hurtado M (2007). Biological nitrogen fixation by common beans (Phaseolus vulgaris L.) increases with bio-char additions. Biology and Fertility of Soils 43(6):699-708. |
|
|
Roy A, Ghosh S, Kundagrami S (2019). Nodulation pattern and its association with seed yield in chickpea (Cicer arietinum L.) germplasms. Indian Journal of Agricultural Research 53(2):172-177. |
|
|
Rutherford A (2011). ANOVA and ANCOVA: a GLM approach. John Wiley and Sons. |
|
|
SAS Institute (2015). Base SAS 9.4 procedures guide. |
|
|
Sethi IB, Sewhag M, Kumar R, Kumar P, Jajoria M (2015). Studies on nodulation of chickpea cultivars as influenced by sowing time and seed rate. Ecology, Environment and Conservation 22:363-365. |
|
|
Sharma S, Yadav N, Singh A, Kumar R (2013). Nutritional and antinutritional profile of newly developed chickpea (Cicer arietinum L) varieties. International Food Research Journal 20(2). |
|
|
Singh B, Singh J, Singh G, Kaur G (2015). Effects of long term application of inorganic and organic fertilizers on soil organic carbon and physical properties in maize-wheat rotation. Agronomy 5(2):220-238. |
|
|
Sreerama YN, Sashikala VB, Pratape VM, Singh V (2012). Nutrients and antinutrients in cowpea and horse gram flours in comparison to chickpea flour: Evaluation of their flour functionality. Food Chemistry 131(2):462-468. |
|
|
Suzaki T, Yoro E, Kawaguchi M (2015). Leguminous plants: inventors of root nodules to accommodate symbiotic bacteria. International Review of Cell and Molecular Biology 316:111-158. |
|
|
Tajer A (2016). What's the function of nitrogen (N) in plants. |
|
|
Tajima R, Lee ON, Abe J, Lux A, Morita S (2007). Nitrogen-fixing activity of root nodules in relation to their size in peanut (Arachis hypogaea L.). Plant Production Science 10(4):423-429. |
|
|
Tena W, Wolde-Meskel E, Walley F (2016). Response of chickpea (Cicer arietinum L.) to inoculation with native and exotic Mesorhizobium strains in Southern Ethiopia. African Journal of Biotechnology 15(35):1920-1929. |
|
|
Thalji T (2006). Impact of row spacing on faba bean L. Growth under Mediterranean Rainfed Conditions. Journal of Agronomy 5(3):527-532. |
|
|
Wallace TC, Murray R, Zelman KM (2016). The nutritional value and health benefits of chickpeas and hummus. Nutrients 8(12):766. |
|
|
Weisany W, Raei Y, Allahverdipoor KH (2013). Role of some of mineral nutrients in biological nitrogen fixation. Bulletin of Environment, Pharmacology and Life Sciences 2(4):77-84. |
|
|
Worku M, Astatkie T (2015). Effects of row spacing on productivity and nodulation of two soybean varieties under hot sub-moist tropical conditions in south-western Ethiopia. Journal of Agriculture and Rural Development in the Tropics and Subtropics 116(2):99-106. |
|
|
Xia X, Ma C, Dong S, Xu Y, Gong Z (2017). Effects of nitrogen concentrations on nodulation and nitrogenase activity in dual root systems of soybean plants. Soil Science and Plant Nutrition 63(5):470-482. |
|
|
Zhao Y, Xiao X, Bi D, Hu F (2008). Effects of Sulfur Fertilization on Soybean Root and Leaf Traits, and Soil Microbial Activity. Journal of Plant Nutrition 31(3):473-483. |
|
|
Zhou X, Liu X, Rui Y, Chen C, Wu H, Xu Z (2011). Symbiotic nitrogen fixation and soil N availability under legume crops in an arid environment. Journal of Soils and Sediments 11(5):762-770. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


