Full Length Research Paper

ABSTRACT
In Mali, the economy is mainly based on agriculture. Rice contributes about 5% of the GDP and constitutes a strategic sector with enormous potential. Despite its strategic importance, rice cultivation faces multiple biotic constraints among which rice yellow mottle virus disease is the most important one with an incidence varying from 60 to 100%. To reduce yield loses due to this disease, we determined the mode of action of two rice yellow Mottle virus (RYMV) pathotypes previously isolated in rice fields in Mali, and identified by Reverse transcription polymerase chain reaction (RT-PCR). Both types significantly decrease rice height with a more pronounced effect with the type A, which showed more aggressivity even on the SK20-28 genotype recognized as RYMV disease-tolerant variety. Root dry weight, grain production and tiller number were the most affected by the disease. Except for tiller number, the virus type A was always more virulent than the type B. Significant positive correlation was found between disease severity and % reduction in tiller number, percentage reduction in plant height, and grain production. The data presented root inhibition as a mode of action of the two RYMV pathotypes isolated in Mali. This information should be used in order to improve the management strategies for these pathogens on field rice in Mali.
Key words: Rice yellow mottle virus, root inhibition, rice, Mali.
INTRODUCTION
Rice yellow mottle virus is a disease of rice causing high losses to rice production (Onwughalu et al., 2010; Traoré et al., 2009). Rice plants affected by this virus show yellow or orange leaves at the early stage of the crop, and leaves roll up and dry in severe cases (Kouassi et al., 2005; Odedera et al., 2016). Stunting, reduced tillering and poor panicle filling are other possible symptoms (Onwughalu et al., 2010; Suvi et al., 2018). The effects of the virus on rice plants result in low or no seed production if affected at booting stage and poor grain quality (Onwughalu et al., 2010). Rice yield losses caused by field infestation by rice yellow Mottle virus (RYMV) were estimated between 17 and 100% (Onwughalu et al., 2011) with significant socio-economic impacts on smallholder farmers (Abo et al., 1998; Onwughalu et al., 2010; Asante et al., 2013; Sereme et al., 2016).
Characterization of rice varieties using the lack of expression of symptoms and the un-detectability of the virus content in plant tissues by ELISA test as resistance factor has made it possible to identify rice (Oryza sativa) varieties (Gigante and Bekarosaka) as well as Oryza glaberrima species (Tog5681, Tog5672, Tog5674 and Tog7291) as very resistant to RYMV (Oludare et al., 2016). For that, the most widely strategy currently used for disease control is genetic resistance, through the use of resistant varieties against pathogens (Samaké et al., 2018; Suvi et al., 2018).
However, the ability of pathogens to overcome resistant genes in crops compromised the durability of these resistances (Ochola and Tusiime, 2011). West and Central Africa have a high pathogenic diversity (Abo et al., 1998; N’Guessan et al., 2001; Oludare et al., 2016; Odedara et al., 2016) and this genetic diversity occurring among RYMV populations seems to present a big challenge for breeding rice for durable resistance to plant virus (Munganyinka et al., 2016). An hypervirulent pathotype of Rice Yellow Mottle Virus was identified in West and Central Africa (Hebrad et al., 2018; Pidon et al., 2017). This pathotype named pathotype T, is able to overcome all type known sources of high resistance and appears to be more abundant than suspected (Hebrad et al., 2018).
Despite the production losses, socio-economic impacts are caused by the disease across Africa, and the ever-increasing resistance breakdown (Sereme et al., 2016); research activities continue mainly in the search for resistant varieties (Ndjiondjop et al., 1999; Albar et al., 2006; Thiémélé et al., 2010; Suvi et al., 2018), and few studies have been conducted to study how the virus affects the growth of the different parts of the plant to impact its production (Opalka et al., 1998; Abebrese et al., 2019).
The studies carried out in this direction have shown that partial digestion of pit membranes resulting from programmed cell death may permit virus migration through them, concomitant with autolysis (Opalka et al., 1998). In addition, the displacement of the Ca2+ from pit membranes to virus particles may contribute to the disruption of the pit membranes and facilitate systemic virus transport (Opalka et al., 1998). It was also shown that the virus negatively impacts plant water and mineral nutrition by initiating an imbalanced growth between roots and leaves (Guinagui et al., 2018). In Mali, these modes of action were not detected by the works of the researcher. This suggests that the rice yellow mottle virus
genotypes affecting rice varieties in Mali use different modes of action. To be able to efficiently control the RYMV disease in Mali, we must know how the virus cause damages to rice plants. In this study, we firstly identified the type of RYMV in rice fields in Mali (Ohasanya et al., 2006). Secondly, we determined the mode of action of the different types of the identified virus using different rice varieties cultivated in Mali (Mogga et al., 2012).
MATERIALS AND METHODS
Study area
To determine the pathogenicity of two types of Rice Yellow Mottle Virus newly isolated in Mali, an experiment was conducted in a greenhouse under artificial conditions at the Laboratory of Research in Microbiology and Microbial Biotechnology (LaboREM-Biotech), Faculty of Sciences and Techniques; University of Sciences, Techniques and Technologies of Bamako in Mali. This experiment was conducted between June and October 2017.
Rice varieties
In this study, nine rice varieties most cultivated in Mali (Table 1) were used. These rice varieties were obtained from the Institute of Rural Economy of Mali (IER).
Isolates of rice yellow mottle virus
Two types of the RYMV isolated from leaves of infected rice plants collected from farmers’ field at Niono, Baguineda and Selingué (Samaké et al., 2018) were used in this study. These two isolates were identified by molecular typing following the method described by Uke et al. (2016), and propagated in greenhouse using the highly susceptible cultivar Kogoni 91-1 by mechanical inoculation of two-week-old seedlings using finger-rub technique (Samake et al., 2018). Four weeks after inoculation, rice leaves showing typical yellow mottle symptoms were harvested to prepare the inoculum used for inoculating rice genotypes in the greenhouse experiment.
Inoculum preparation
The inoculum used was prepared according to the method by Munganyinka et al. (2016). Briefly, one gram of infected leaf tissue was first crushed in a drop of doubly-distilled water using sterile mortars and pestles until 80% of the leaf tissue material was macerated. The resultant leaf extract was diluted 10 times by adding10 ml of doubly-distilled water and kept in a refrigerator.
Experimental design and treatments
In this experiment, we used a complete randomized bloc design (CBRD). The RYMV isolates type A and type B, and the non-inoculated represent the three blocs studied. Rice varieties represent the twenty-one treatments studied. Each treatment was replicated four times. Five seeds of each tested rice variety were seeded directly in a plastic pot filled with 10 kg of soil. After one week the seedlings were thinned to two plants per pot. The cultivar Kogoni 91-1 was used as the highly susceptible check while Gigante was the highly resistant check. Each pot was constantly
supplied with tap water in the morning. To avoid confusion between the yellowing associated with disease development and that due to malnutrition, 2 g of NPK fertilizer was applied to the plants at 28 days after planting followed by application of 2 g urea at 45 days after planting and at early flowering stage of the plants (Onwughalu et al., 2010).
Inoculation of rice varieties
The inoculation which started 14 days after transplanting using a mixture of carborundum powder to aid the infection, was performed by rubbing the mixture onto the leaves from the base to the top using pieces of cotton wool (Munganyinka et al., 2016). Care was taken to ensure maximum wetting and formation of mild bruises that acted as infection passageways, according to Ochola and Tusiime (2011).
Data collection
Disease severity
Appearance of symptoms and disease progress were monitored for each inoculated genotype from 7 to 32 days post inoculation. Disease severity was scored with a version of the scale developed by John and Thottapilly (1987) and modified by Ochala et al. (2011), where 0 was equivalent to non- infection and 9 was 100% infection.
Components of the yield
Data on important agronomic traits were collected according to Munganyinka et al. (2016). The data include the plant height of inoculated and uninoculated plants (measured from the soil surface to the tip of the shoot in cm); root architecture and length were determined and measured after carefully washing the roots; the number of tillers per plant was recorded for each hill, and the seed weight was determined. These data were used to assess the impact of the disease on the growth of the rice (Munganyinka et al., 2016). In each case, the impact of the disease was assessed using the following formula:
Impact (%) = (Ni-I) × 100/Ni (Zouzou et al., 2008)
Where: Ni = mean values on the seedlings not inoculated and I= mean values on the seedlings inoculated.
Data analysis
Analysis of variance was performed for each parameter (Rice genotypes and Virus types) according to the General Linear Model (GLM) procedures using SAS software (version 9.2). Whenever the Fisher test indicates a significant effect at a probability of 0.05, Fisher's Protected Least Significant Difference (LSD) test was used to compare the means.
RESULTS AND DISCUSSION
Effect of different types of RYMV on rice aerial and root growth
Effect of two RYMV pathotypes isolated in Mali on rice aerial growth
The effect of two types RYMV on the growth of the aerial part of the rice is presented in Figure 1. No significant effect was observed on the growth of the aerial part of the two rice genotypes after inoculation with water in the absence or presence of carburundum (Figure 1). However, inoculation with RYMV shows a decrease in plant size with a stronger effect of the two types of RYMV in Kogoni 91-1 (Figure 1). Indeed, both types of RYMV significantly decrease the hard rice size with a more pronounced effect with the RYMV type A on Kogoni 91-1. RYMV type A was more aggressive even on the SK20-28 genotype, which is recognized as disease-tolerant, than type B (Figure 1).

Effect of RYMV on rice root growth
The effect of two RYMV types on rice root growth is shown in Figure 2. No significant effects of inocution with water were observed on root growth. However, inoculation with RYMV showed a decrease in root size in the Kogoni91-1 and BG 90-2 genotypes with a strong effect of both RYMV types in Kogoni 91-1 (Figure 2). The more root growth is inhibited, the less the aerial part of the rice grows (Figures 1 and 2).

Effect of the different types of RYMV isolated in Mali on rice yield components
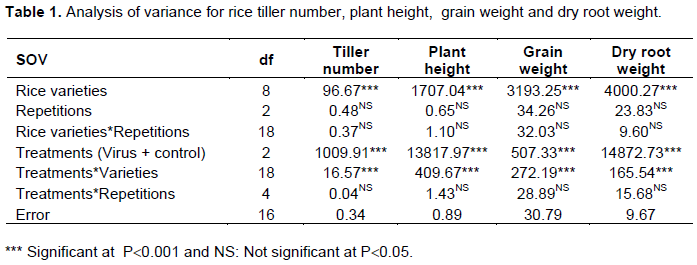
The mean squares for the effect of RYMV disease on rice yield components are presented in Table 1. Rice varieties, treatments (virus pathotypes and an uninoculated control) and their interactions significantly (P ≤ 0.001) affected rice tiller number, plant height, grain weight and dry root weight (Table 1). Contrariwise, no significant effect was observed with repetions and their interaction with rice varieties or treatments.
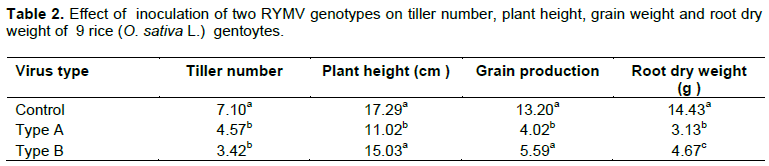
Inoculation with RYMV reduced the yield parameters studied as compared to the non-inoculated treatments (Table 2). Root dry weight, grain production and tiller number were the most affected by the disease. Except for tiller number, the virus type A was always more virulent than the type B (Table 2). Results presented in Table 3 show the disease effect on yield components.
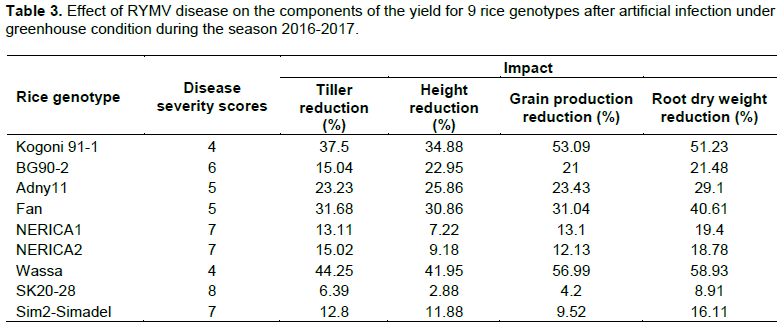
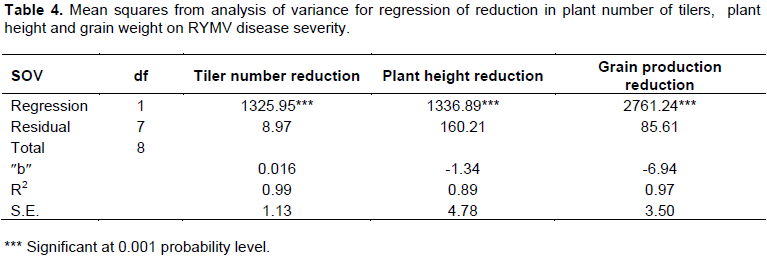
Results showed that RMYV infection reduced rice production between 4.2 to 56.99% depending on the genotypes whereas reduction in tiller number per plant varied from 6.39 to 44.25%, plant height from 2.88 to 41.95% and plant root dry weight from 8.91 to 51.23% (Table 3). The rice genotype Sim2-Simadel showed a small reduction in grain production compared to the tolerant check SK20-28. The susceptible check varieties, Kogogni 91-1 lost 53.09% of its production. The variety Wassa which appears to be more succeptible than Kogoni 91-1, lost 56.99% of its production in contrast to NERICA 2, NERICA 1 and BG90-2, which lost 12.13, 13.1 and 21% respectively (Table 3). Significant positive correlation was found between disease severity and % reduction in tiller number (R2=0.99), percentage reduction in plant height (R2=0.89), and grain production (R2=0.97) (Table 4).
DISCUSSION
Root growth and plant height of rice genotypes Kogoni 91-1 and BG90-2 (Oryza sativa L.) were strongly inhibited by the two types of RYMV isolated from Mali. These results confirmed that the intraspecific lines (Indica species) were highly susceptible (Munganyinka et al., 2016). Guinagui et al. (2018) showed that RYMV infected leaves contain less minerals (P, K, Ca and Mg) and chlorophyll than those of the non-infected rice plants. The early inhibition of rice root growth may affect water and mineral absorption and can explain the reduction in grain yield in suceptible rice varieties, mainly in O. sativa L. intraspecific lines. Contrary, to succeptible rice lines, in the presence of these two types of RYMV the rice genotypes SK20-28 and Sim-simadel showed no significant inhibition in root growth. Abebrese et al. (2019) indicated that some rice hybrids decrease the incidence of the RYMV disease and improve rice yield. So, they recommaded the introduction of these rice hybrids.
In this work the effect of RYMV disease on yield components differed among varieties and the type of virus. Results were confirmed by a highly significant regression of disease score on % reduction in tiller number, plant height, grain production and dry root yield. Results from this study confirm the findings of Munganyinka et al. (2016) in which they showed that RMYV infection reduced plant height between 2.0 to 20.3% depending on genotype; whereas tiller number per plant varied from 4.84 to 30.6%. Onwughalu et al. (2010) stated that the behavior of the rice varieties they tested varies by variety and the parameter selected, and revealed negative corrections with RYMV infection.
Likewase, Guinagui et al. (2018) obtained a high yield loss of 94.4% in Bouake 189 at seedling and booting infection stages. These results establish the fact that yield losses to RYMV are strongly influenced by host cultivars as well as time of virus infection, and revealed that the period from seedling and booting represents the most vulnerable phase to RYMV infection in rice growth stages (Onwughalu et al., 2010).
CONCLUSION
The rice varieties Kogoni91.1 and BG90-2 were very susceptible to rice yellow mottle virus. These rice varieties were the most appreciated by consummers and smallholder farmers in Mali. Two RYMVs were identified in rice fields in Mali. It appeared through this reseach results that: these two RYMV genotypes impact negatively the tested rice varieties’ growth and yield, by strongly inhibiting root sizes (length, width and volume). Thus, improving rice root growth in the presence of RYMV disease could improve growth and yield of susceptible rice varieties.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
The authors appreciate USAID (through the PEER program), Marketplace (through Africa-Brazil) and the World Bank (through WAAPP2a) for the financial support of this study.
REFERENCES
|
Abebrese SO, Yeboah A, Dogbe W, Dartey PKA, Akromah R, Gracen VE, Offei SK, Danquah EY (2019). Evaluation of Yield, Reaction to Diseases, and Grain Physical Attributes of Some Introduced Rice Hybrids in Ghana. International Journal of Agronomy, pp. 1-8. |
|
|
Abo ME, SY AA, Alegbejo MD (1998). Rice Yellow Mottle Virus (RYMV) in Africa: Evolution, Distribution, Economic Significance on Sustainable Rice Production and Management Strategies. Journal of Sustainable Agriculture 11:51-58. |
|
|
Albar L, Bangratz-Reyser M, Hébrard E, Ndiondjop M., Jones M, Ghesquière A (2006). Mutations in the eIF (iso) 4G translation initiation factor confer high resistance of rice to Rice yellow mottle virus. Plant Journal 47:417-426. |
|
|
Asante D, Asante BO, Acheampong G K (2013). Farmer and consumer preferences for rice in the Ashanti region of Ghana: Implications for rice breeding in West Africa. Journal of Plant Breeding and Crop Science 5(12):229-238. |
|
|
Guinagui NB, Sorho F, Sanogo S, Koné B, Koné D (2018). Effect of Rice yellow mottle virus, Sobemovirus on the Contents of N P K Ca and Mg in Leaves of Infected Rice. Annual Research and Review in Biology 30(3):1-10. |
|
|
Hebrad E, Pinel-Galzi A, Oludare , Poulicard N, Aribi J, Fabre S, Siluée D (2018). Identification of a hypervirulent pathotype of Rice yellow mottle virus: A threat to genetic resistance deployment in West-Central Africa. Phytopathology 108(2):299-307. |
|
|
Kouassi NK, N'Guessan P, Albar L, Fauquet CM, Brugidou C (2005). Distribution and Characterization of Rice yellow mottle virus: A Threat to African Farmers. Plant Disease 89(2):1-11. |
|
|
Mogga ML, Lamo J, Asea G, Gibbson P, Edema R (2012). Reaction of rice cultivars to a virulent rice yellow mottle virus strain in Uganda. African Crop Science Journal 20(s1): 51-57 |
|
|
Munganyinka E, Edema R, Lamo J, Gibson P (2016). The reaction of intraspecific and interspecific rice cultivars for resistance to rice yellow mottle virus disease. European Journal of Experimental Biology 6(3):13-18. |
|
|
N'Guessan P, Pinel A, Sy A, Ghesquière A, Fargette D (2001). Distribution, Distribution, pathogenecity and interactions of two strains of Rice yellow mottle virus in forested and savannah zones of West Africa. Plant Disease 85:59-64. |
|
|
Ndjiondjop MN, Albar L, Fargette D, Fauquet C, Ghesquière A (1999). The Genetic Basis of High Resistance to Rice yellow mottle virus (RYMV ) in cultivars of two cultivated Rice species. Plant Disease 83:931-935. |
|
|
Ochola D, Tusiime G (2011). Pathogenicity of Rice Yellow Mottle Virus and the potential sources of resistance against the disease in eastern Uganda. Asian Jornal of Plant Pathology 5(1):1-15 |
|
|
Odedara OO, Ademolu KO, Ayo-John EI (2016). Prevalence of Rice Yellow Mottle Virus (RYMV) on Rice Plants Grown in Selected Farms in Ogun State: Preliminary Results. Nigerian Journal of Biotechnology 31:96-102. |
|
|
Oludare A, Tossou HT, Kini K, Silue D (2016). Diversity of Rice yellow mottle virus in Benin and Togo and Screening for Resistant Accessions. Journal of Phytopathology 164:924-935 |
|
|
Onwughalu JT, Abo ME, Okoro JK, Onasanya A, Séré Y (2010). The Effect of Rice yellow mottle virus Infection on the Performance of Rice (Oryza sativa L.) Relative to Time of Infection under Screenhouse Condition. Journal of Applied Sciences 10:1341-1344. |
|
|
Onwughalu J, Abo ME, Okoro JK, Onasanya A, Séré Y (2011). Rice yellow mottle virus infection and reproductive losses in rice (Oryza sativa Linn). Trends in Applied Sciences Research 6(2):182-189. |
|
|
Opalka O, Brugidou C, Bonneau C, Nicole M, Beachy RN, Yeager M, Fauquet C (1998). Movement of rice yellow mottle virus between xylem cells through pit membranes. Proceedings of the National Academy of Sciences 95:3323-3328 |
|
|
Pidon H, Ghesquière A, Chéron S, Issaka S, Hébrard E, Sabot F (2017). Fine mapping of RYMV3: A new resistance gene to Rice yellow mottle virus from Oryza glaberrima. Theoretical and Applied Genetics. Springer Berlin Heidelberg; 0: 0. |
|
|
Samaké S, Traoré D, Goita O, Sarra S, Dao S, Doumbia B, Babana AH (2018). Genetic diversity of rice yellow mottle virus from Niger Office and Selingue Development Rural Office in Mali. Journal of General and Molecular Virology 8(1):1-7. |
|
|
Sereme D, Ouedraogo I, Wonni I, Yao N, Neya BJ, Konate G (2016). Assessment of yield losses due to Rice yellow mottle virus under field conditions in Burkina Faso. International Journal of Current Advanced Research 5(12):1522-1528. |
|
|
Suvi WT, Shimelis H, Laing M (2018). Breeding rice for rice yellow mottle virus resistance in Sub-Saharan Africa: A review. Acta Agriculturae Scandinavica, Section B - Soil and Plant Science 69(2):1-8. |
|
|
Thiémélé D, Boisnard A, Ndjiondjop MN, Chéron S, Séré Y, Aké S (2010). Identification of a second major resistance gene to Rice yellow mottle virus, RYMV2, in the African cultivated rice species. Theoretical and Applied Genetics 121:169-179. |
|
|
Traoré O, Pinel-Galzi A, Sorho F, Sarra S, Rakotomalala M, Sangu E, Kanyeka Z, Sérég Y, Konaté G, Fargette D (2009). A reassessment of the epidemiology of Rice yellow mottle virus following recent advances in field and molecular studies. Virus Research 141:258-267. |
|
|
Uke A, Asea G, Natsuak KT (2016). Phylogenetic analysis and serotyping of Rice Yellow Mottle Virus strains in Uganda. Tropical Agriculture and Development 60(2):81-88. |
|
|
Zouzou M, Kouakou TH, Koné M, Issaka S (2008). Screening rice (Oryza sativa L.) varieties for resistance to Rice yellow mottle virus. Scientific Research and Essays 3(9):416-424. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


