Full Length Research Paper

ABSTRACT
The response of maize to a composite inoculation of Arbuscular Mycorrhizal Fungi (AMF) in fields in Central Benin was evaluated. The 2000 SYNEE maize variety underwent 3 AMF groups involved in treatments namely: T1 = "Control (AMF and fertilizer free)"; T2 = "Glomeraceae"; T3 = Acau "Acaulosporacea"; T4 = Diversi " Diversisporaceae"; T5 = "50% NPK + Urea"; T6 = "Glomeraceae + 50% NPK + Urea"; T7 = "Acaulosporaceae + 50% NPK + Urea"; T8 = "Diversisporaceae + 50% NPK + Urea"; T9 = "100% NPK + Urea". The 2 factors were combined in a split-split plot design with a complete randomized block layout including 3 repetitions. The main and subplot factors were respectively the AMF type and use of fertilizers. The growth parameters (height, noose diameter and leaf area), grain yield and endomycorrhizal infection of plants were evaluated. T6 had induced the highest height, fresh above, underground biomass and grain yield of maize passing respectively from 42.38, 25.58, 58.33 and 27.18%, values recorded by the control plants. Moreover, T7 held the highest values of noose diameter and T8 those of mycorrhization intensities. However, T2 and T4 generated the highest values of mycorrhization frequencies and spore number. These results highlighted the advantages of endogenous AMF for sustainable agriculture in Benin.
Key words: Bio-fertilizers, mycorrhizal, composite, maize, sustainable agriculture, Benin.
INTRODUCTION
Arbuscular mycorrhizal fungi (AMF), beneficial microbes to agricultural and natural ecosystems (Pellegrino et al., 2015), improve the host plant's (P) nutrition, growth and resistance to drought and disease (Kumari et al., 2019).
Thus, AMF present in most soils and colonizing the roots of the majority (80%) of plants (Walder and van der Heijden, 2015) have received particular attention in relation to their application in sustainable agriculture (Chen et al., 2018). Under natural conditions, these obligatory symbionts, unlike ectomycorrhizal symbionts, do not have host specificity and in many cases, a differential effect in each species has been observed with different natures of mycorrhizal inoculum (Diallo et al., 2016; Kouadio et al., 2017). Indeed, Van der Heijden et al. (1998) and Abbas (2014) have shown that the benefits of a fungus or AMF complex are not identical within the same plant species (Anguiby et al., 2019). Mycorrhization also increases plants’ performance by improving growth (Pavithra et al., 2018), water status and nutrient accumulation (Kapoor et al., 2013). Consequently, increases in yields, particularly for maize and wheat, were highlighted (Treseder, 2013; Pellegrino et al., 2015). During this process, mycorrhizal colonization promotes the establishment of extensive hyphae networks and the secretion of glomaline, which in turn contributes to the absorption of water and nutrients (Pagano, 2014), thus; contributing to a better soil structure (Miller and Jastrow, 2010).
However, under intensive agricultural conditions, AMF have a less significant effect than under natural conditions. The reduction in the number of plant species found in a limited geographical area, such as wheat-corn rotation (Sasvári 2017), regular soil disturbance (Balogoun, et al., 2013), excessive use of mineral fertilizers and fungicide use (Jin et al., 2013) inexorably lead to a decline in the number and activity of AMF (Bakonyi and Csitári, 2018). In areas of constantly exploited land, the number of AMF propagules is seriously reduced (Gottshall et al., 2017). Also, the inadequacy of the fungus introduced under new edaphic conditions sometimes leads to its disappearance (Duponnois et al., 2013; Plenchette et al., 2000). However, Bakonyi and Csitári, (2018) noted that the effectiveness of AMF inoculation depended on soil nutrient availability, type of tillage, fertilizer applied, use of protective agents (especially fungicides) and climate. It therefore appears necessary to study the mycorrhizal crops status and identify the indigenous mycorrhizal complex associated with them (Tchinmegni et al., 2016) in order to integrate this technology into agricultural operations as long as it has been proven that the effects of mycorrhizal symbiosis in a controlled environment (greenhouse) differ from those obtained in a real environment (field) (Hoeksema et al., 2010). For example, Lekberg and Koide (2005) demonstrated that the AMF effects are negligible in real environments than in greenhouses. According to this observation, Bakonyi and Csitári (2018) insisted on the size of the experiment, which must go beyond that of experiment in a controlled environment for fear to be irrelevant.
Thus, this study proposes to evaluate the maize response to composite inoculation of AMF in real conditions. Its objective is to promote the use of mycorrhizal fungi in the technical itinerary of maize production. Moreover, it consists of: assessing the growth parameters of maize after inoculation of a few endogenous AMF and measuring their mycorrhization criteria to determine the best combination.
MATERIALS AND METHODS
The 2000 SYN EE-W maize variety was used during the experiment. These are 75-day early varieties, developed by the International Institute of Tropical Agriculture (IITA) and the National Institute of Agricultural Research of Benin (INRAB) (MAEP, 2017).
Determination of soil chemical parameters
Soil samples were collected at a depth of 0-20 cm (Adjanohoun et al., 2011). Sampling consisted of 500 g composite samples at the level of each plot before the test was set up just after ploughing and before any application of fertilizer amendments. These samples were sent to the Soil Sciences, Water and Environment Laboratory (LSSEE) of the National Institute of Agricultural Research of Benin (INRAB). These analyses consisted of the determination of pH (water) (by glass electrodes in a soil-to-water ratio of 1/2.5), organic carbon (Anne, 1945), assimilable P (Bray and Kurtz, 1945), total N (Kjeldahl, 1883), and exchangeable bases using the Metson (1956)’s method with ammonium acetate at a pH of 7.
Isolation of AMF
The groups of mycorrhizal fungi Glomeraceae (Funeliformis mosseae, Funeliformis geosporum, Glomus caledonius, Glomus ambiosporum, Rhizophagus intraradices and Septoglomus contrictum), Acaulosporaceae (Acaulospora capsicula, Acaulospora denticulata) and Diversisporaceae (Diversispora globifera) were identified from the maize rhizosphere of the 6 different municipalities in the department of Collines using the Davis- INVAM key. These different groups of fungi were isolated and multiplied over a period of twelve weeks using sorghum as a trap plant because of its high mycorrhizogenic potential and as a substrate clay + sterile soil at respective doses of 3∕4 and 1∕4 in a 250 ml pot.
Trapping and preparation of inoculums
Sorghum seeds were previously soaked for 2 min in a 0.024% sodium hypochlorite (NaClO) solution and rinsed 5 times with distilled water under vortex shaking (Gholami et al., 2009). Then, 10 sorghum seeds (Sorghum bicolor L.) were sown in each pot containing different levels of soil substrate dilution. The pots were then placed in a greenhouse at room temperature. The plants were watered daily with (tap) water to maintain soil capacity similar to that in the field for 6 weeks. The root biomass and substrate were therefore crushed to obtain the inoculums.
Seed coating
The inoculums obtained were used to coat the seeds in a 10:1 ratio (10 kg of seed per 1kg of bio-product) for each group of fungi. Each mixture was mixed with a quantity of water equivalent to 600 ml.kg-1 of fertilizers. The coated seeds were dried in ambient air for 12 h in accordance with the recommendations published by Fernández et al. (2000).
Experimental design
The trial was installed at the Miniffi Research and Development (RD) station in the municipality of Dassa-Zoumé in Benin. The choice was based on the decline of soil fertility. Flat ploughing was carried out the day before sowing on a not flooded plot with a slope of at most 2%. Each plot had an area of 12.8 m² and consisted of 4 lines of 4 m long with a spacing of 0.80 m. Seeding was done at a spacing of 0.80 m x 0.40 m with a density of 31,250 plants/ha (Yallou et al., 2010). The distances between the plots and the repetitions between them were 1.8 m and 2 m, respectively. The useful plot had an area of 6.4 m2, where data were collected on the 2 central lines. The different treatments are: T1 = "Control (no mycorrhizal fungus and no P addition)"; T2 = "Glomeraceae"; T3 = Acau "Inoculation with Acaulosporaceae"; T4 = Various "Diversisporaceae"; T5 = "50%NPK+Urea"; T6 = "Glomeraceae + 50%NPK+Urea"; T7 = "Acaulosporaceae +50%NPK+Urea"; T8 = "Diversisporaceae+50%NPK+Urea"; T9 = "100%NPK+Urea". Two factors were combined in a split-split plot system (split-split plot with a complete randomized block layout) with 3 repetitions; the large plot factor was the mycorrhizal fungus type and the subplot factor was the use of chemical fertilizer. Mineral fertilizer rate recommended by INRAB (1995) was 200 kg/ha NPK.
Sowing and maintenance of plots
Two coated or uncoated maize (Zea mays L.) seeds were sown 5 cm deep. The demarcation to one plant per hole was done at 7th DAS (Day after sowing). The various parameters were collected every 15 days from the 15th to the 60th DAS. Only the leaf area was measured at the 60th DAS.
Evaluation of growth parameters
The effects of the different treatments on maize growth were assessed by measuring the plant height, noose diameter, number of leaves and leaf area. The height of maize plant is the distance between the noose and the last ligule. It was measured with a measuring tape, while the noose diameter was measured with a caliper every 15 days, the 15th, 30th, 45th, 60th DAS. The area of the leaves with ligule was estimated by multiplying the product of leaves length (from the top of the sheath to the tip of the blade) and width (measured in the middle of the blade) by the coefficient 0.75 (Ruget et al., 1996). This measure concerned the last 2 leaves with ligule of each plant and was evaluated at the 60th DAS.
Evaluation of performance parameters
Biomass produced
The determination of above and underground biomass yield was determined as:
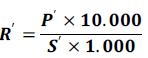
Where: R '= average yield of dry biomass of maize plants t. ha-1,P = weight of dry biomass of maize plants in kg, 10000 is the conversion of hectare in m²; 1000 converting tonne (t) into kg, S = crop area cultivated in​​ m².
Maize grain yield
The harvested maize cobs were weighed per treatment using a precision scale. The moisture content of the grains was determined using a moisture meter (LDS-1F). The average of maize grain yield was determined by Valdés et al. (2013):
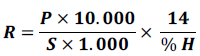
R = Average yield in maize grains (t. ha-1); P = weight of maize grains in kilograms (kg) ; 10000 is the conversion from ha to m2; 1000 is the conversion from tonne (t) to kg; S = harvest area in m²; % H = percentage of grain moisture in %.
Evaluation of endomycorrhizal infection of plant roots
At 40 DAS, maize root samples were collected. After staining the roots with trypan blue according to the method described by Phillips and Haymain (1970), root fragments from maize plants were observed in binoculars (XSP-BM-2CEA .2013). Estimation of mycorrhizal root infection was performed using the intersection method (Giovannetti and Mosse, 1980; Trouvelot et al., 1986). Two parameters of arbuscular mycorrhizal infections were used to calculate the mycorrhization rate: The frequency of mycorrhization (F) which reflects the degree of infection of the root system:
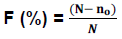
Where N corresponds to the number of fragments observed and no is the number of fragments without any trace of mycorrhization.
The mycorrhization intensity: m (absolute mycorrhization intensity) which expresses the portion of the colonized cortex relative to the entire root system:
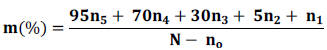
In this formula, n5, n4, n3, n2 and n1 are the numbers of fragments respectively noted in the five infection classes indicating the importance of mycorrhization namely: 5 = more than 95%, 4= from 50 to 95%, 3= 30 to 50%, 2 = 1 to 30%, 1= 1% of the cortex.
Data analysis
All analyses were performed with software R (3.5.3) (R Core Team 2018). A repeated measurement variance analysis was performed on the height, diameter and values related to leaves with the nlme (Linear and Nonlinear Mixed Effects) package (Pinheiro et al., 2019). For Fresh Above Biomass (FAB), Leaf Area (LA) and fresh underground biomass (FUB), we performed multiple variance analysis.
RESULTS
Soil chemical characteristics
The soil chemical properties of the site before the installation of the tests (Table 1) generally showed that the soil of Miniffi, the centre of Benin was slightly basic (pH = 7.8) at the horizon, 0-20 cm. The soil showed intermediate fertility and was richer in Ca2+ than the potassium (K+) exchangeable cations. The carbon/nitrogen (C/N) ratio was relatively low at the topsoil level. The P concentration in the topsoil (47.5 mg/kg soil) was low.
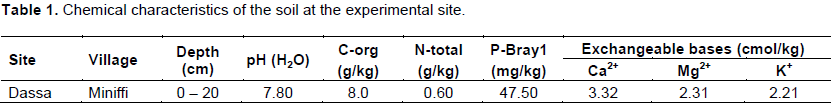
Influence of AMF on the height and noose diameter of maize plants
Height
Figure 1 shows the evolution of the height depending on strains and time. A significant effect (p<0.001) of both strains and treatments was observed. Glomeraceae-50NPK+Urea followed by Acaulosporaceae-50NPkUrea were the treatments that induced the highest plant values over the entire period of the experiment (Figure 1) with respective increases of 42.38, 36.59% compared to the control.
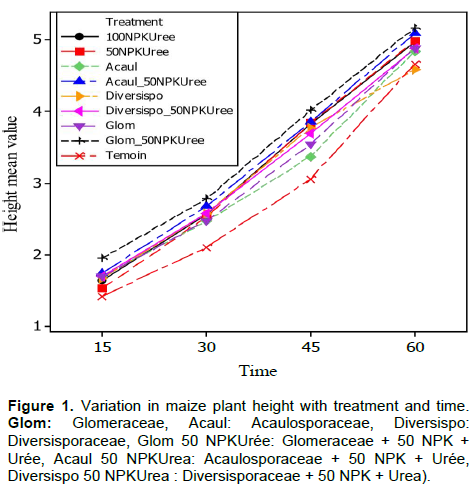
Noose diameter
The results of the plant analysis revealed that the plants grew in diameter gradually according to the strains and blocks over time with a significant effect (p<0.001). The largest diameters were consistently generated by Acaulosporaceae + 50% NPK + Urea followed by Glomeraceae + 50% NPK + Urea (Figure 2).
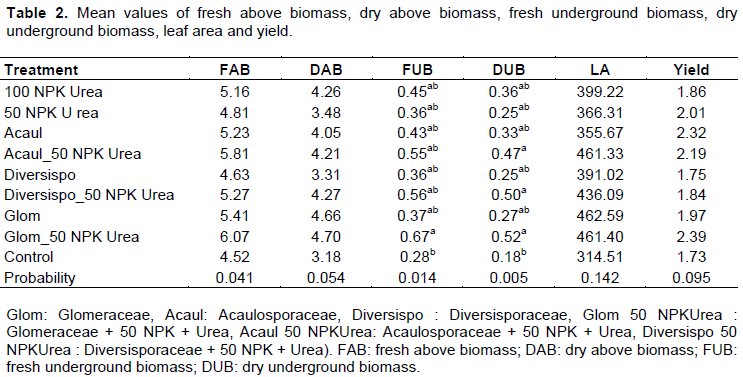
Influence of AMF on Biomass produced, leaf area of plants and maize grain yield
Analysis of the results in Table 1 revealed that Glomeraceae + 50% NPK Urea induced an important fresh above biomass (FAB) followed by Acaulosporaceae + 50% NPK Urea with respective increases of 25.58 and 22.32% over the control. The results of the multiple variance analyses revealed significant differences between treatments for fresh underground biomass (FUB) and dry underground biomass (DUB).
Glomeraceae + 50% NPK Urea generated high value of FUB with an increase of 58.33% compared to control.
Regarding DUB, Glomeraceae + 50% NPK Urea, Diversporaceae + 50% NPK+ Urea and Acaulosporaceae + 50 NPK Urea have induced a high and similar values. No significant differences were found between yield average and leaf area (LA) mean values. The best grain yield was obtained with Glomeraceae + 50% NPK Urea while the largest leaf area was obtained with Glomeraceae only, exceeding the control by 27.18 and 32.01%, respectively (Table 2).
Mycorrhization parameters
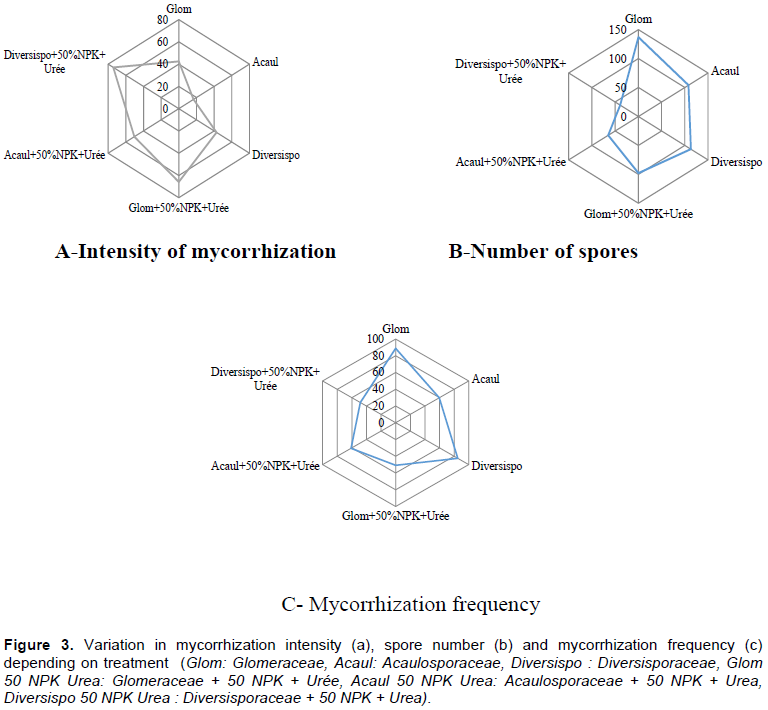
The analysis of the mycorrhization parameters presented in the radar diagram showed that Diversporaceae + 50% NPK+ Urea followed by Glomeraceae + 50% NPK + Urea induced the highest mycorrhization intensities unlike Acaulosporaceae which generated the lowest intensity (Figure 3a). With regard to mycorrhization frequency (Figure 3c) and spore number (Figure 3b), Glomeraceae and Diversisporaceae induced the highest values.
DISCUSSION
AMF are soil-borne fungi that can significantly improve plant nutrient uptake and resistance to several a biotic stress (Sun et al., 2018), enabling thereby the establishment of plants in land (Rouphael et al., 2015). However, use of appropriate AMF, either individually or in combination, still remains a big challenge as the benefits from the mutualism greatly vary depending on the AMF strains (Chen et al., 2017). This study was conducted to evaluate the effect of a composite AMF inoculation on maize in field conditions in Central Benin. The results of the soil samples taken from the site revealed that the soil prior to installation was slightly basic (pH = 7.8) at the horizon of 0-20 cm. The soils have intermediate fertility and are richer in Ca2+ than in K+ ions. The carbon nitrogen-C/N ratio was relatively low at the topsoil level. Those data are similar to those observed by Igue et al. (2015) and the weakness of soil characteristics suggested a good expression of AMF. Hence, the mycorrhization of maize with endogenous strains has significantly enhanced growth parameters such as height, noose diameter, while there is similar effect on leaf area. This improvement may be attributed to both a good mineral uptake of plant (Malonda et al., 2019; Gnamkoulamba et al., 2018) and AMF adaptability to environment traits (Benjelloun, 2014). Benjelloun (2014) proved better that plants of maize inoculated grew better than those not inoculated.
Growth in height, noose diameter at the experimental site was significantly different (p<0.01). Glomeraceae-50NPK+Urea had induced the greatest heights while the greatest diameters were obtained with Acaulosporaceae-50NPK-Urea with respective increases of 42.38, 34.89% compared to the control. Leaf areas of maize plants were not significantly different and the largest leaf area was obtained with only Glomeraceae, exceeding the control by 32.01% (Table 2). Balliu et al. (2015) supported these results by observing an improvement in leaf area, nitrogen, potassium, calcium and phosphorus levels, indicating an increase in growth of tomato plants. These rates of improvement in the height, noose diameter and leaf area of maize plants are believed to be due to the positive effect of AMF, which might facilitate the absorption of mineral elements by plants (Malonda et al., 2019) and decrease the uptake of Na and Cl (Evelin et al., 2012), leading to growth stimulation of mycorrhizal plants (Gnamkoulamba et al., 2018).
Glomeraceae+50%NPKUrea induced a large FAB and FUB with increases of 25.58 and 58.33% respectively compared to the control. The same trends were observed by Gnamkoulamba et al. (2018a, 2018b) who recorded a significant total dry rice biomass and indicated an increasing trend with the inoculation dose. These results would be due to the significant improvement in the water balance, the absorption of certain trace minerals and macronutrients from the plants by AMF; an increase in the production of photosynthates improves the accumulation of biomass (Nyembo et al., 2012; Mitra et al., 2019) interrelated to photosynthetic performance of a plant (Chen et al., 2017). Zhang et al. (2018) demonstrated the AMF involvement in amelioration of the biomass allocated to stems, panicles and grains with an increased redistribution of phosphorus and nitrogen to grains. The effects of AMF used in this study were clearly demonstrated through the improvement of all growth and yield parameters. The best grain yield was obtained with Glomeraceae+ 50%NPKUrea, exceeding the Control by 27.18% (Table 2). This should be explained by the production of glomalin-related soil protein containing 30–40% of carbon and its related compounds that safeguard soil from desiccation by enhancing the soil water holding capacity (Sharma et al., 2017). This rate of grain yield improvement was below the 51% obtained by Aboubacar et al. (2013) in cowpea after inoculation of mycorrhizal fungi and slightly above the 20% got by Ceballos et al. (2013) in cassava. These results support the high mycorrhization frequency and intensity and the large numbers of spores obtained. Therefore, Diagne et al. (2003) mentioned that host plants got many benefits when intensity of mycorrhisation is above12%. Beltrano et al. (2013) and Sarah and Ibrar (2016) suggested that spore density and host colonization are largely defined by the compatibility of AMF with host plants, environment and interactions between AMF and exudates produced by the host plant. In addition, the importance of AMF is also highlighted by the increase in mycorrhizal infection even under hostile conditions. Our study demonstrated that the combinations, Diversporaceae+50% NPK+ Urea followed by Glomeraceae+50%NPK+Urea had the highest mycorrhization intensities, showing a significant development of fungal mechanisms promoting nutrient exchange between the 2 symbiotic partners (Benjelloun et al., 2014).
CONCLUSION
This study compared the behaviour of different groups of AMF on maize growth and grain yield variables in field environment. Glomeraceae50NPK+Urea induced the highest heights, yield of fresh above-ground and underground fresh biomass with the highest grain yields of maize exceeding the values measured in the control plants by respectively 42.38, 25.58, 58.33% and 27.18%. Regarding the noose diameter, the largest values were measured with Acaulosporaceae+50%NPK. These results showed that the mycorrhization of maize with the different strains of used AMF could improve the productivity of this crop by halving the use of mineral fertilizers. Thus, it is therefore imperative to popularize this technology by developing an organic fertilizer based on AMF for sustainable agriculture in Benin.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
The authors appreciate the project of West Africa Agricultural Productivity Program (WAAPP) and the National Fund of the Scientific Research and the Innovation (FNRSIT) for their financial supports of this work.
REFERENCES
|
Abbas Y (2014). Microorganisms in the rhizosphere of Tetraclinaria: a tool to optimize the assisted regeneration of Tetraclinis articulates Vahl. M.Sc., Ph.D. Thesis, Mohammed V University, Faculty of Science Rabat, 157p. |
|
|
Aboubacar K, Zakari MO, Harouna IA, Seydou I, Alzouma M, Zoubeirou (2013). Effect of co-inoculation of rhizobium and mycorrhizae on the agronomic performance of cowpeas (Vigna unguiculata (L.) Walp.) in Niger. Journal of Applied Biosciences 72:5846-5854. |
|
|
Anguiby BLA, Ouattara G, Bomisso EL, N'goran B, Ouattara B, Coulibaly SA, Aké S (2019). Evaluation du statut mycorhizien d'arbres de Ceiba pentandra (L), Gaertn et Tieghemella heckelii (A.Chev), Pierre, du jardin Botanique de Bingerville en Côte d'Ivoire Journal of Applied Biosciences 138:14092-14105. |
|
|
Anne P (1945). Sur le dosage rapide du carbone organique dans les sold Ann: Agroni Avril, Mai, Juin, 1945, 5he année 2:161-172. |
|
|
Bakonyi I, Csitári G (2018). Response of winter wheat to arbuscular mycorrhizal fungal inoculation under farm conditions. Journal of Agriculture and Environmental Sciences 5:1. |
|
|
Balliu A, Sallaku G, Rewald B (2015). AMF Inoculation enhances growth and improves the nutrient uptake rates of transplanted, salt-stressed tomato seedlings. Sustainability 7:15967-15981. |
|
|
Balogoun I, Saïdou A, Ahoton LE, Adjanohoun A, Amadji GL, Ezui G, Youl S, Mando A, Igué AM, Sinsin BA (2013). Détermination des formules d'engrais minéraux et des périodes de semis pour une meilleure production du maïs (Zea mays L.) au Sud et au Centre Bénin. Bulletin de la Recherche Agronomique du Bénin, ISSN sur papier : 1025-2355, Numéro spécial Fertilité du maïs, pp.: 1-25 |
|
|
Beltrano J, Ruscittil M, Arango MC, Ronco M (2013). Effects of Arbuscular Mycorrhiza Inoculation on Plant Growth, Biological and Physiological Parameters and Mineral Nutrition in Pepper Grown under Different Salinity and P Levels. Journal of Soil Science and Plant Nutrition 13(1):123-141. |
|
|
Benjelloun S, El Harchli EH, Amrani JK, El Ghachtouli N, Fikri Benbrahim K, El Yamani J (2014). Etude De L'importance De la Mycorhization dans la Synthèse des composés phénoliques chez le Maïs (Zea mays L.) en condition de stress hydrique. Research Inventy: International Journal of Engineering and Science 4(12):43-49. |
|
|
Bray RH, Kurtz LT(1945). Determination of total organic and available forms of phosphorus in soils. Soil Science 59:39-45. |
|
|
Ceballos I, Ruiz M, Fernández C, Peña R, Rodriguez A, Sanders IR (2013). The in vitro mass-produced model mycorrhizal fungus, Rhizophagus irregularis, significantly increases yields of the globally important food security crop cassava. PLOS ONE 8(8):e70633. |
|
|
Chen S, Zhao H, Zou C, Li Y, Chen Y, Wang Z, Jiang Y, Liu A, Zhao P, Wang M, Ahammed GJ (2017). Combined Inoculation with Multiple Arbuscular Mycorrhizal Fungi Improves Growth, Nutrient Uptake and Photosynthesis in Cucumber Seedlings. Frontiers in Microbiology 8:2516. |
|
|
Chen MM, Arato L, Borghi E, Nouri, Reinhardt D (2018). Beneficial Services of Arbuscular Mycorrhizal Fungi - From Ecology to Application. Frontiers in Plant Science 9:1270. |
|
|
Diallo B, Samba SAN, Sane D (2016). Effects of MA fungi on the growth and development of castor seedlings grown under saline stress under semiconductor conditions. Revue des Energies Renouvelables19(1):59-68. |
|
|
Duponnois R, Ramanankierana H, Hafidi M, Baohanta R, Baudoin E, Thioulouse J, Lebrun M (2013). Endemic plant resources to sustainably optimize forest cover rehabilitation operations in Mediterranean and tropical environments: example of plants that facilitate the spread of mycorrhizal fungi. Biology Reports 336 (5-6):265-272. |
|
|
Evelin H, Giri B, Kapoor R (2012). Contribution of Glomus intraradices inoculation to nutrient acquisition and mitigation of ionic imbalance in NaCl-stressed Trigonella foenum-graecum. Mycorrhiza 22:203-217. |
|
|
Fernández F, Gómez R, Vanegas LF, de la Noval BM, Martínez MA (2000). Mycorrhizogenic inoculant product. National Office of Industrial Property. Cuba, Patent No. 22641. |
|
|
Gholami A, Shahsavani S, Nezarat S (2009). The Effect of Plant Growth Promoting Rhizobacteria (PGPR) on Germination, Seedling Growth and Yield of Maize. World Academy of Science, Engineering and Technology 49:19-24. |
|
|
Giovannetti M, Mosse B (1980). An evaluation of techniques for measuring vesicular- arbuscular infection in roots. New Phytologist 84:489-500. |
|
|
Gnamkoulamba A, Tounou AK, Tchao M, Tchabi A, Adjevi AKM, Batawila K (2018). Field assessment of the growth potential and production of rice (Oryza sativa L.) variety IR841 inoculated in the nursery by four strains of arbuscular mycorrhizal fungi. European Scientific Journal 14:12. |
|
|
Gottshall CB, Cooper M, Emery SM (2017). Activity, diversity and function of arbuscular mycorrhizae vary with changes in agricultural management intensity. Agriculture, Ecosystems and Environment 241:142-149. |
|
|
Hoeksema JD (2010). A meta-analysis of context-dependency in plant response to inoculation with mycorrhizal fungi. Ecology Letters 13:394-407. |
|
|
Igue MA, Oga AC, Balogoun I, Saidou A, Ezui GYS, Kpagbin G, Mando A, Sogbedji JM (2016). Détermination des formules d'engrais minéraux et organiques sur deux types de sols pour une meilleure productivité de maïs (Zea mays l.) dans la commune de Banikoara (Nord-Est Du Bénin). European Scientific Journal 12:30. |
|
|
Jin H, Germida JJ, Walley FL (2013). Suppressive effects of seed-applied fungicides on arbuscular mycorrhizal fungi (AMF) differ with fungicide mode of action and AMF species. Applied Soil Ecology 72:22-30. |
|
|
Kapoor R, Evelin H, Mathur P, Giri B (2013). Arbuscular mycorrhiza: Approaches for abiotic stress tolerance incropplantsforsustainableagriculture. In Plan tAcclimation to Environmental Stress; Tuteja, N., Gill, S.S., Eds.; Springer: New York, NY, USA,; pp. 359-401. |
|
|
Kjeldahl J (1883). Neue Methode zur Bestimmung des Stickstoffs in organischen Körpern. Zeitschrift für Analytische Chemie 22:366-382. |
|
|
Kouadio AN, Nandjui J, Zézé A (2017). An Indigenous and an Exotic AMF Strains Improve Dioscorea Alata growth and induce tolerance to Meloidogyne spp., Agricultural Science Research Journal 7(2):63-69. |
|
|
Kumari S, Merina P, Srimeena N (2019). Arbuscular Mycorrhizal Fungi (AMF) Induced Defense Factors against the Damping-off Disease Pathogen, Pythium aphanidermatum in Chilli (Capsicum annum). International Journal of Current Microbiology and Applied Sciences 8(6):2243-2248. |
|
|
Lekberg Y, Koide RT (2005): Is plant performance limited by abundance of arbuscular mycorrhizal fungi? A meta-analysis of studies published between 1988 and 2003. New Phytologist 168:189-204. |
|
|
MAEP (Ministère de l'Agriculture de l'Elevage et de la Pêche) (2017). Recueil des technologies agricoles prometteuses développées par le Système National de Recherche Agricole (SNRA) de 1996 à 2015 Document Technique et d'Informations. ISBN : 978-99919-2-985-9 Dépôt légal n° 9433 du 12 juin 2017 Bibliothèque Nationale du Bénin, 2èmetrimestre. 288p |
|
|
Malonda AN, Nzola-meso TM, Manga AM, Yandju MC (2019). Effet des champignons mycorhiziens Arbusculaires sur le phosphore des sols tropicaux et implication dans la biosynthèse du caroténoïde du manioc. Journal of Applied Biosciences 135:137. |
|
|
Miller RM, Jastrow JD (2010). Mycorrhizal fungi influence soil structure. In: Kapulnik, Y., Douds, D. (Eds.) Arbuscular mycorrhizas: Physiology and function. Chapter 1. Kluwer Academic Publishers. |
|
|
Mitra D, Navendra U, Panneerselvam U, Ansuman S, Ganeshamurthy AN, Divya J (2019). Role of mycorrhiza and its associated bacteria on plant growth promotion and nutrient management in sustainable agriculture. International Journal of Life Sciences and Applied Science 1:1-10. |
|
|
Pagano MC (2014). Drought stress and mycorrhizal plant. In Use of Microbes for the Alleviation of Soil Stresses; Miransari, M., Ed.; Springer: New York, NY, USA 1:97-110. |
|
|
Pavithra D, Yapa N (2018). Arbuscular mycorrhizal fungi inoculation enhances drought stress tolerance of plants. Groundwater for Sustainable Development 7:490-494. |
|
|
Pellegrino E, Opik M, Bonari E, Ercoli L (2015). Responses of wheat to arbuscular mycorrhizal fungi: A meta-analysis of field studies from 1975 to 2013. Soil Biology and Biochemistry 84:210-217 |
|
|
Phillip JM, Hayman DS (1970). Improved procedures for cleaning roots and staining parasitic and vesicular arbuscularmycorrhizal fungi for rapid assessment of infection. Transactions of the British Mycological Society 5:158-161. |
|
|
Pinheiro J, Bates D, DebRoy S, Sarkar D, R Core Team (2019). nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 31-141 https://CRAN.R-project.org/package=nlme. |
|
|
Plenchette C, Bois JF, Duponnois R, Cadet P (2000). Mycorrhization (Glomus aggregatum) of millet (Pennisetum glaucum). Soil Studies and Management 7(4):379-383. |
|
|
Rouphael Y, Franken P, Schneider C, Schwarz D, Giovannetti M, Agnolucci M (2015). Arbuscular mycorrhizal fungi act as biostimulants in horticultural crops. Scientia Horticulturae 196:91-108. |
|
|
Ruget F, Bonhomme R, Chartier M (1996). Estimation Simple de la Surface Foliaire de Plantes de Maïs en Croissance. Agronomien 16(9):553-562. |
|
|
Sarah S, Ibrar M (2016). Effects of arbuscular mycorrhizal fungi on spores density and root colonization of four hybrids of sunflower (Helianthus annuus L.) at different rock phosphate levels. Sarhad Journal of Agriculture 32(4):258-266. |
|
|
Sasvári Z (2017). Study of arbuscular mycorrhizal fungal diversity in long-term field experiments. PhD dissertation, Szent István University, GödöllÅ‘. |
|
|
Sharma S, Prasad R, Varma A, Sharma AK (2017). Glycoprotein associated with Funneliformis coronatum, Gigaspora margarita and Acaulospora scrobiculata suppress the plant pathogens in vitro. Asian Journal of Plant Pathology 11(4):192-202. |
|
|
Sun Z, Song J, Xin X, Xie X, Zhao B (2018). Arbuscular mycorrhizal fungal proteins 14-3-3- are involved in arbuscule formation and responses to a biotic stress during AM symbiosis. Frontiers in Microbiology 5:9-19. |
|
|
Tchinmegni FI, Tsobeng AC, Ngonkeu MEL, Tchoundjeu Z (2016). Evaluation du statut mycorhizien chez Allanblackia floribunda en vue de sa domestication en zones forestières humides du Cameroun, Revue Scientifque et Technique Forêt et Environnement du Bassin du Congo 6:81-83. |
|
|
Treseder KK (2013). The extent of mycorrhizal colonization of roots and its influence on plant growth and phosphorus content. Plant and Soil 371:1-13. |
|
|
Trouvelot A, Kough JL, Gianinazzi-Pearson V (1986). Measurement of the VA mycorrhization rate of a root system. Research and estimation methods of functional significance. In: Physiological and genetic aspects of mycorrhizae, Dijon, 1985. INRA (ed.), pp. 217-221. |
|
|
Van der Heijden MG, Boller T, Wiemken A, Sanders IR (1998). Different arbuscular mycorrhizal fungal species are potential determinants of plant community structure. Ecology 79(6):20822091. |
|
|
Walder F, van der Heijden MGA (2015). Regulation of resource exchange in the arbuscular mycorrhizal symbiosis. Nature Plants 1:15159 |
|
|
Zhang F, Jia-Dong HE, Qiu-Dan NI, Qiang-Sheng WU, Zou YN (2018). Enhancement of drought tolerance in trifoliate orange by mycorrhiza: changes in root sucrose and proline metabolisms. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 46:270. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


