Full Length Research Paper

ABSTRACT
The crop-livestock-forest integration systems are a form of sustainable production, creating a consortium between the cultivation of crops and forest production and the creation of pastures for livestock breeding, seeking a synergy between the system components. Therefore, integrating agricultural crops with tree species provides countless benefits to the components, such as the maintenance and increase in nutrient cycling. Therefore, the objective of this study was to evaluate the biomass and nutritional stock of Brachiaria pasture and accumulated litter in a silvopastoral system with tree species of the Cerrado. Three silvopastoral systems, formed with the forage species Urochloa decumbens (Brachiaria) integrated with three tree species, namely Dipteryx alata (baru tree), Caryocar brasiliense (pequi) and Eugenia dysenterica (cagaita), were evaluated. Litter and pasture were collected in each system at different sample distances from the tree. The amount of biomass and the nutritional contents of both components were evaluated by chemical analysis. The largest amount Brachiaria pasture biomass and largest amount of litter was found in the silvopastoral system with baru trees. The pasture differed nutritionally. The pastures associated with baru trees and to pequis have higher amounts of macronutrients. There are nutritional differences with respect to the sampling positions.
Key words: Brachiaria, Baru, Pequi, Cagaita, CLFi, Cerrado, Brazil.
INTRODUCTION
The expansion of agriculture has contributed to the growth of Brazil as a supplier of agricultural products (Townsend et al., 2009). However, the unrestrained expansion of agriculture has caused profound changes in natural resources, in addition to changes in natural vegetation and soil properties (Lago et al., 2012; Santana et al., 2016). The aggressiveness of agriculture can be controlled in the way the activities are practiced and in how the environment is managed. Within this context, the crop-livestock-forest integration (CLFi) or agro-sylvopastoral system, is a sustainable production strategy that integrates forestry, agricultural and livestock activities, in the same area, either in intercropping, succession or rotation (Machado et al., 2011; Lana et al., 2016).
Integrated systems can be classified into four types (Balbino et al., 2011): crop-livestock-forest integration (CLFi/agrosilvopastoral), crop-livestock integration (CLi/agropastoral), livestock-forest integration (LFi/silvopastoral) and crop-forest integration (CFi). Soil fertility improvement, increased carbon sequestration and the supply of forage and more favorable environmental conditions for animals are highlighted amongst the benefits (Costa et al., 2002; Abreu et al., 2016). The tree component in agroforestry systems helps maintain nutrient cycling (Montagnini, 1992; Upson et al., 2016) by decomposing litter. Decomposition of litter is considered the primary means of transfer of nutrients to the soil, enabling its reabsorption by the living plant (Schumacher et al., 2004; Santana et al., 2016). The shade of trees and the presence of litter reduce the high soil temperatures and the trees also attenuate wind speed (Monteith et al., 1991; Ong et al., 1991; Assis et al., 2015; Brito et al., 2015). These two factors affect evaporation rate, soil water balance and evapotranspiration, as it increases the humidity available to plants, influencing the yield of agricultural crops and pastures (Duboc, 2007; Lemos-Junior et al., 2016).
Some LFi studies have shown native or exotic species being used in association with animals, as well as reviews of agronomic crops (Lemos-Junior et al., 2016). Duboc (2005) stated that to which extent, there is greater disclosure and encouragement for using native Cerrado species in an interaction system, an increase in the interest in the rational cultivation of these species may occur. At this juncture and with the need for a production that is increasingly sustainable and compatible with livestock and forestry production, the objective of this study was to evaluate biomass and nutritional stock of Brachiaria pasture and litter in silvopastoral system with tree native species of Cerrado, in Brazil.
MATERIALS AND METHODS
This study was carried out at the experimental area of the School of Agronomy, of the Federal University of Goias (Universidade Federal de Goiás; SA/UFG), at Goiânia, Goiás. In the area, the soil is a red oxisol and the plantations near one another. According to Köppen, the climate is predominantly tropical (Aw), with the marked division of two well defined seasons during the year: humid summer, from December to March, and dry prevailing winter from June to August. The average temperature varies between 18 and 26°C, as the average annual rainfall is about 1300 -1700 mm, concentrating in the months from October to March (in the spring and summer seasons). Between May and September is the dry season, a period when the rains are rare and may occur drought. Between the months of July to August, the humidity drops too (dry weather) and may be between 15 and 30%. Three silvopastoral systems formed with the forage species Urochloa decumbens (Stapf) R.D.Webster (Brachiaria) integrated with three tree species, Dipteryx alata Vog. (Baru tree), Caryocar brasiliense Camb. (Pequi) and Eugenia dysenterica D.C (Cagaita) were evaluated. Each system has a total area of 7590, 5725 and 15525 m2, respectively.The plantations were carried out in January 1998, spaced 6 x 6 m, and the plants come from a germplasm collection of the SA/UFG (Access to the map of the Germplasm bank).
Litter and above-ground biomass were collected following an experimental design of randomized blocks with six replicates, for each system and the sampling distances from the tree were recorded. A total of five above-ground biomass and three litter samples were collected, where each block was located 50 m from each other. All the material was stored in properly labeled Kraft paper bags.
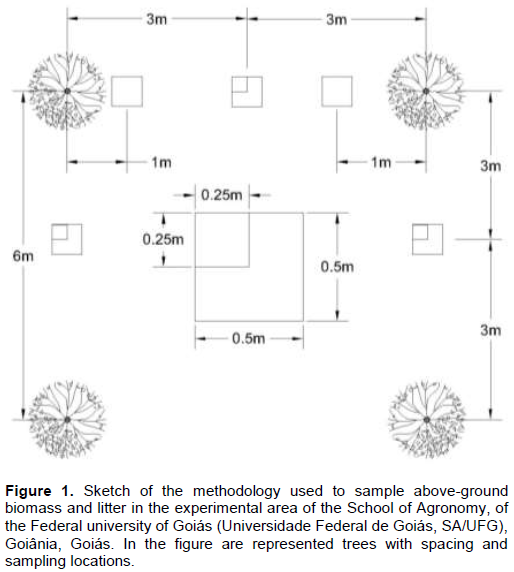
Litter was collected following the methodology determined by Lima et al. (2015), using a template of 0.25 x 0.25 cm (0.0625 m2), in the same place in which the above-ground was harvested, removing all the material until the soil was exposed. Roots still attached to the soil were avoided. Each sample was packed into labeled paper bags and taken to the Forest Ecology Laboratory.
The above-ground and litter of Brachiaria samples were dried in the laboratory, in a circulating and air renewal chamber at 70°C for approximately 72 h until reaching a constant weight. The final weighing of the Brachiaria samples were carried out individually. In addition, the total weight of each Brachiaria sample was obtained, and then each sample was divided into leaves and mixture (root, flowers and seeds). Each fraction was weighed once again.
The nutrition stock was evaluated using two samples for each tree per component (above-ground biomass and litter) in each different position. Three of the samples collected were mixed, and two sub-samples were made from the mixture, that is, two composite samples. At the end, a total of six samples of pasture and four samples of litter were analyzed per system of tree species, adding up to 30 samples.
The material of the composite samples was milled (Lippel crusher and in a Wiley type mill), and finally sieved in a sieve with mesh opening of 1.0 mm (20 mesh). Then, the material was directed to a laboratory for chemical analyses to determine micronutrient and macronutrient quantities in the plant tissue, following the methodology described by Tedesco et al. (1995) and Miyazawa et al. (1999).
Statistical analyses
A principal components analysis (PCA) was carried out with a covariance matrix to assess the nutritional contents in pasture and litter, and for the following parameters nutrition stock with regards to the different tree species: nitrogen (N), phosphorous (P), potassium (K), calcium (Ca), magnesium (Mg), sulfur (S), sodium (Na), copper (Cu), iron (Fe), manganese (Mn), zinc (Zn), cobalt (Co) and molybdenum (Mo) (Monteiro et al., 2016). The different sampling locations (between lines, between plants and one meter from the plant) were also considered in the analyses (ANOVA two way). All analyses were conducted considering a significance level of 95%.
RESULTS
The highest Brachiaria above-ground biomass was observed in the silvopastoral system with Baru trees (2.23 ton/ha; F=8.51; p=0.001) (Figure 2A). Above-ground biomass in this system was approximately 23% higher than in Cagaita system (1.81 ton/ha) and 32% higher than in Pequi system (1.69 ton/ha) (Figure 2A). Above-ground Brachiaria biomass within the systems with Cagaita and Pequi trees did not differ statistically.

The Baru system had a higher above-ground biomass in the center of the sampling area, between the tree lines (F=2.68; p=0.03). Still, above-ground biomass of Brachiaria in the Baru system was similar to that observed for the other systems in the position within the same row and 1 m from the tree (Figure 2B). Above-ground of Brachiaria was similar in all sampling distances for the Cagaita and Pequi systems.
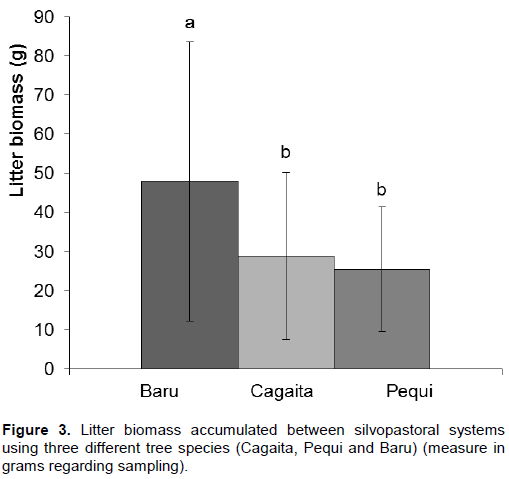
The Baru system had a higher amount of litter biomass (7.67 ton/ha), of about 66% more litter than the area with Cagaita trees (4.63 ton/ha) and 88% more than the area with Pequi trees (4.07 ton/ha) (F= 675.46; p=0.000; Figure 3). There were no significant differences between the system with Cagaita and Pequi trees (Figure 3). In addition, there was no difference in the production of accumulated litter between areas at the center, between tree lines or between plants for either system with the different tree species.
The pasture showed significant differences in its nutritional balance in the presence of the tree species, as well as among sampling positions (Table 1). The stocks of N had a significant difference in the central sampling position associated with the cagaita tree. The Brachiaria had 12% more N when associated with the Baru tree and 8% when associated with the pequi tree. Nitrogen stocks were also significantly affected when the Brachiaria was collected between plants in the system integrated with the baru tree. Therefore, pasture integrated with Baru trees exhibited a 13% increase in N when analyzed with regards to the same sampling position for Cagaita trees.

The stocks of K in the Brachiaria were statistically different when samples were taken from the position of 1 m in the cagaita system. Despite being significant, K stocks were 10% smaller in a same sampling position for systems with the pequi and 4% smaller when for systems with Baru trees. Ca stocks in the pasture also exhibited significant differences in the central sampling position associated with the Pequi, and were 3% smaller in the Cagaita system. The amount of the element Mg in Brachiaria differed among systems in the central position of sampling, was 21% higher for the pequi than for the Baru tree and 17% lower for the Brachiaria in the Cagaita system, for the same sampling position. The Na stock in the pasture had significant differences for the “1 m” collection point, where it was 6% higher for Baru systems than for the cagaita system and 13% higher than the Pequi system. Differences in Na stock were also observed for the position “between plants”, where the pasture from the cagaita system had 14% more Na than the pequi and 8% more than the baru system.
The amount of Cu differed significantly for the “1 m” collection point, where 170% more Cu was recorded in the pasture for the Cagaita than in the pequi system and 91% more than in the Baru. The amount of Fe in the pasture differed between sampling positions, being greater at 1 m away from the plant for the Pequi, Baru and Cagaita systems. However, the Cagaita system had 15% more Fe than the Pequi system. The element Mn was also recorded in higher amounts for samples collected 1 m from the Cagaita trees, and was 142% lower for the Pequi trees and 79% lower for the Baru trees.
The nutrition stocks of the litter also showed some distinctions. The element N had a significant difference with regards to the collection position for the three native species. The litter of Baru trees had 3% more N at the central sampling position and 136% more N than the Pequi. The litter of Baru trees had 44% more S than when associated with Cagaita trees and 13% more than when associated with Pequi, for samples collected between plants.
The Na stocks in the litter of Baru systems was 20% higher than the recorded for systems with Cagaita and 22% more than for pequi, for samples collected between plants. The litter of the system integrated with Baru trees had 3% more Na in the central sampling area than the Cagaita system and 5% more than the system integrated with Pequi. The litter of Baru systems had 83% more Fe than the Cagaita system, and 12% more than the litter of the Pequi system, in the central sampling area. The stock of Fe in the litter of the baru system was 131% higher than the stock recorded for the litter of the Cagaita system and 140% more than the observed for the Pequi system in the sampling position “between plants”.
The elements recorded in larger quantities in the litter were Ca, Fe and Mn. In addition, these elements were directly and positively related, where the higher the calcium, the higher the iron and manganese contents. On the other hand, the relationship observed for these three elements opposes that observed for N, P and K. The elements N, P and K occur in lower quantities in the litter and larger amounts were observed in the pastures integrated with the three native Cerrado species, especially in those integrated with baru and pequi trees (Figure 4). The pastures integrated with Baru and Pequi trees have higher amounts of macronutrients with regards to the nutritional characteristic of the grass. Other mineral elements such as Zn, Mo, Co, S, Cu and Na were recorded in higher concentrations in pastures associated with Cagaita trees, and were all positively related with each other.

DISCUSSION
The largest amount of Brachiaria pasture biomass and largest amount of litter was found in the silvopastoral system with Baru trees. The pasture differed nutritionally, being that pastures associated with Baru trees and Pequi have higher amounts of macronutrients. The livestock farming forest integration system seems to be an efficient system in agroforestry production and in improving the quality of pastures of the Cerrado. The use of trees native to the biome in the pastures make it more similar to natural areas of Cerrado sensu strictu and “campo sujo”, in addition to the benefits also inherent in the natural areas such as increasing organic matter deposition, nutrient cycling and improving soil cycling and microclimate characteristics.
The tree canopies break down wind and change the microclimate of the understory, consequently reducing the evaporation rate of herbaceous plants (Castro et al., 2008). The areas beneath the canopy and litter production retain soil humidity in dry periods, improving the production of grass. Shading caused by trees may impair the productivity of grasses, due to competition for light, in addition to the natural competition for water and nutrients. Therefore, spacing is essential for a good development of the tree-grass system, reducing the competition imposed by the shadows of the canopies (Galzerano and Morgado, 2008; Venturoli et al., 2015). The spacing for planting vary depending on the species and are important for a good soil cover (Gonçalves et al., 2015). The most commonly used spacing range from 4 to 16 m between lines and 50 cm to 2 m between plants (within the lines) (Gonçalves et al., 2015).
Sousa et al (2007) studied the productivity of Brachiaria brizantha cv. Marandu in a silvopastoral system and noted that despite dry matter (DM) reduced due to shadowing, this reduction is associated with larger Brachiaria leaves instead of stems. They also reported an increase in crude protein content in relation to the Brachiaria grown under full sun.
Andrade et al. (2003), assessed the development of six forage grasses (B. brizantha cv. Marandu, B. brizantha cv. MG-4, B. decumbens cv. Basilisk, Panicum maximum cv. Mombaça, Melinis minutiflora and Hyparrhenia rufa) in a silvopastoral system in Paracatu, Minas Gerais, a Cerrado region, associated or not with the leguminous plants Stylosanthes guianensis cv. Mineirão and Eucalyptus sp. Grasses were established between Eucalyptus lines, in plots of 12 x 10 m, and the highest biomass production capacity was recorded for the species B. brizantha cv. Marandu and B. decumbens.
The production litter by falling leaves, twigs, flowers and fruits contribute to the transfer of nutrients from the canopy to the ground, and the leaves account for approximately 60% of this transfer (Poggiani, 2012). The tree species used in this forest and livestock integration system are semi-deciduous, and produce leaves from July to September (Silva-Júnior, 2005). The plants partially lose their leaves at the same time of year, increasing the deposit of nutrients in litter and soil (Caldeira et al., 2007). The low concentration of potassium (K) in the litter may be related to the high mobility of the K within plants, which transfer the element from senescent leaves for younger parts. Potassium is also movable in the soil, so much of it is lost by leaching, especially in the rainy season (Caldeira et al., 2007).
Calcium (Ca) showed higher values in the litter because it is immobilized in the leaves, and is rarely redistributed to other parts of the plant when it is in senescence. Calcium is a movable element in soil, which causes a loss of Ca by erosion and leaching to deeper layers of the soil. The roots of the trees are pivotable, and so reach greater soil depths, facilitating the reabsorption of the elements that were leached. Grasses have fasciculated roots, therefore occupy mostly the surface of the soil (Vitti, et al., 2006; Silva-Neto et al., 2015).
Micronutrient levels in the litter followed a descending order of Fe, Mn, Zn, Cu. The same result was reported in a research conducted by Caldeira et al. (2007), studying the production of accumulated litter and nutrient contents in a Mixed Rain Forest, at General Carneiro, state of Paraná. Iron (Fe) is considered an immobile element in the plant. Therefore, iron deficiency symptoms are observed faster than is the deficiency of other mobile elements. Because of the limited redistribution in the plant, it is possible to find young leaves with Fe deficiency while old leaves and roots have accumulated Fe (Dechen and Nachtigall, 2006). The high iron content found in the litter may be associated with the high contents found in some species or the contamination of litter samples with soil (Caldeira et al., 2007).
Oxides and sulfides (Mn) are the forms of manganese found most frequently in the soil, and are commonly found associated with Fe. The Mn concentration varies between plant parts and growth period. Older leaves accumulate higher concentrations of manganese and a small portion is translocated to young leaves (Dechen and Nachtigall, 2006). Thus, the accumulated litter showed higher amounts of manganese due to the accumulation in the leaves.
Nitrogen (N) is one of the elements found in larger quantities in the leaves due to their participation in the metabolic reactions of the plant, especially photosynthesis (Malavolta, 1985). Phosphorus (P) is one of the primary macronutrients, along with N and K. Phosphorus is crucial for the process of photosynthesis. Still, it is not required in large amounts in plant, leading to the low values recorded. In addition, the need of the element decreases when the plant is already established, that is, adult (Raij, 1991)
Pastures grown under the baru and pequi trees showed better nutritional aspects with respect to NPK. Oliveira et al. (2005) obtained similar results and state that the increased levels of these minerals must be directly connected to the greater accumulation of organic matter in soil arising from the litter deposition under the canopies.
Trees exploit nutrients found in deep soil layers through the range of their root system. However, forage species only access the most superficial nutrients. Therefore, trees assist in the gradual incorporation of nutrients into the soil/pasture system by depositing biomass onto the soil (Nair, 1999). Carvalho and Xavier (1999) also found higher mineral contents in soils under B. decumbens grassland that were intercropped with baru and pequi trees. The trees in silvopastoral systems affect the development of the herbaceous forage increasing soil nutrient and changing the microclimate under the canopies (Carvalho and Xavier, 1999).
Cavalcante et al. (2007) evaluated the spatial variability of MO, P, K and CTC under different managements, recording higher values in Cerrado and no-till areas and a reduction in organic matter contents in the other cultivated areas. This fact can be explained by litter accumulation in the larger forest than in other systems. In addition, P, K and CTC values were higher in the no-till and conventional farming systems. The P and K levels were higher in surface, decreasing the deeper layers.
The benefits of the integrated system in the Cerrado directly improves productivity and quality of the pastures, and enables the production of alternative products, such as fruits of the Cerrado and even wood from sustainable plantations as the CLFi (Abreu et al., 2016, in press). The Cerrado has a richness of native trees with potential of being used in commercial production and in the integrated systems. However, the lack of studies on the behavior of these tree species, forages and crops in a mixed system is a limiting factor.
CONCLUSION
Systems integrated with Baru trees had higher productivity, that is, higher biomass, for both Brachiaria pasture and litter production in this study. The production of biomass and litter was higher in line among the trees.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
The research group is grateful to the Postgraduate Program in Agronomy of the Federal University of Goiás (UFG), the Foundation of the State of Goiás Research (FAPEG) for project financing
REFERENCES
|
Abreu SAH, Arruda EM, Barros LR, Almeida RF, Maranhão DDC, Silva VL, Silva-Neto CM, Flores RA, Calil FN, Collier LS (2016). Chemical attributes of the soil in agroforestry systems subjected to organic fertilizations. African J. Agricult. Res. 11(27):2378-2388 |
|
|
Andrade CMS, Garcia R, Couto L, Pereira OG, Souza AL (2003). Desempenho de Seis Gramíneas Solteiras ou Consorciadas com o Stylosanthes guianensis cv. Mineirão e Eucalipto em Sistema Silvipastoril. Rev. Bras. Zoot. 32(6):1845-1850. |
|
|
Assis PCR, Stone LF, Medeiros JC, Madari BE, Oliveira JM, Wruck FJ (2015). Atributos físicos do solo em sistemas de integração lavoura-pecuária-floresta. Rev. Bras. Eng. Agrí. Amb. 19(4):309-316. |
|
|
Balbino LC, Cordeiro LAM, Porfírio-da-Silva V, Moraes A, Martine GB, Alvarenga RC, Kichel NA, Fontaneli RS, Santos HP, Franchini JC, Galerani PR (2011). Evolução tecnológica e arranjos produtivos de sistemas de integração lavoura-pecuária-floresta no Brasil. Pesq. Agrop. Bras. 46:10. |
|
|
Brito TR, Soares LC, Carmo TD, Fernandes LA (2015). Nutrição e produtividade de Brachiaria decumbens em sistema silvipastoril e monocultivo. Cad. Ciên. Agrár. 7(1):22-26. |
|
|
Caldeira MVW, Marques R, Soares RV, Balbinot R (2007). Quantificação de serapilheira e de nutrientes - Floresta Ombrófila Mista, Montana- Paraná. R Acad. Curitiba. 5(2):101-116. |
|
|
Carvalho MM, Xavier AF (1999). Sistemas silvipastoris para recuperação e desenvolvimento de pastagens. Faculdade de Ciências Ágrarias e Veterinárias. UNESP. |
|
|
Castro AC, Junior JBL, Santos NFA, Monteiro EMM, Aviz MAB, Garcia AR (2008). Sistema silvipastoril na Amazônia: ferramenta para elevar o desempenho produtivo de búfalos. Ciên. RuraL 38(8):2395-2402. |
|
|
Cavalcante EGS, Alves MC, Pereira GT, Souza ZM (2007).Variabilidade espacial de MO, P, K e CTC do solo sob diferentes usos e manejos. Ciên. Rur. 37:2. |
|
|
Gonçalves KG, Duarte GSD, Filho AAT (2015). Espécies frutíferas do cerrado e seu potencial para os SAFs. FLOVET – Boletim do Grupo de Pesquisa da Flora, Vegetação e Etnobotânica 1(7):64-79. |
|
|
Costa RB; Arruda EJ, Oliveira LCS (2002). Sistemas agrossilvipatoris como alternativa sustentável para a agricultura sustentável para a agricultura familiar. Rev. Inter. Desenv. Local. 3(5):25-32. |
|
|
Dechen AR, Nachtigall GR (2006). Micronutrientes. In: Fernandes, Manlio Silvestre. (editor). Nutrição mineral de plantas. Sociedade Brasileira de Ciência do Solo. Cap. 13:327-354. |
|
|
Duboc E (2005). Desenvolvimento inicial e nutrição de espécies arbóreas nativas sob fertilização, em plantios de recuperação de áreas de Cerrado degradado. Tese Doutorado. Universidade Estadual Paulista, Faculdade de Ciências Agronômicas, Botucatu 151 p. |
|
|
Duboc E (2007). Sistemas agroflorestais e o Cerrado. In: Faleiro F, Farias-Neto, AL (Ed.). Savanas: desafios e estratégias para o equilíbrio entre sociedade, agronegócio e recursos naturais. Planaltina, DF: Embrapa Cerrados, 37 p. (Embrapa Cerrados. Documentos, 197). |
|
|
Freitas ECS, Neto SNO, Fonseca DM, Santos MV, Leite HG, Machado VD (2013). Deposição de serapilheira e de nutrientes no solo em sistema agrossilvipastoril com eucalipto e acácia. R. Árvore. 37(3):409-417. |
|
|
Galzerano L, Morgado E (2008). Eucalipto em Sistemas Agrossilvipastoris. Rev. Elect. Vet. 93 p. |
|
|
Lana RMQ, Lana ÂMQ, Reis GL, Miranda E (2016). Productivity and nutritive value of brachiaria forage intercropping with eucalyptus in a silvopastoral system in the Brazilian Cerrado biome. Aust. J. Crop Sci. 10(05):654-659. |
|
|
Lago WNM, Lacerda MPC, Neumann MRB (2012). Indicadores de qualidade dos solos na microbacia do Ribeirão Extrema, Distrito Federal: parte II. Rev. Bras. Eng. Agríc. Amb. 16(7):721-729. |
|
|
Lima NL, Silva-Neto CMS, Calil FN, Souza LR, Moraes DC (2015). Acúmulo de serapilheira em quatro tipos de vegetação no Estado de Goiás. Enciclopédia Biosfera 11(22): 39-46. |
|
|
Lemos-Junior JM, Souza, KR, Guimarães LE, Oliveiro FD, Monteiro MM, Goncalves RA, Silva-Neto CM, Lima NL, Venturoli, F, Calil FN (2016). Volumetric models for Eucalyptus x urograndis in a crop-livestock-forest integration (CLFI) system in Brazilian Cerrado. African J. Agric. Res. 11:1337. |
|
|
Machado LAZ, Balbino LC, Ceccon G (2011). Integração Lavoura-Pecuária-Floresta. 1. Estruturação dos Sistemas de Integração Lavoura-Pecuária. Dourado, MS: Embrapa Agropecuária Oeste, Série Embrapa, Documentos P. 110. |
|
|
Malavolta E, Ferri MG (1985). Nutrição de plantas. Fisiologia vegetal. São Paulo: EDUSP 1:400-408. |
|
|
Miyazawa M, Pavan M, Muraoka T, Carmo CAFS, Mello WJ (1999). Análises químicas de tecido vegetal. In: Silva FC (Ed.). Manual de análises químicas de solos, plantas e fertilizantes. Brasília: EMBRAPA Solos pp. 171-223. |
|
|
MontagninI F (1992). Sistemas agrofloretales: princípios y aplicaciones em los trópicos. 2 ed. ver. aum. San José: Organización para Estúdios Tropicales 622 p. |
|
|
Monteith JL, Ong CK, Corlett JE (1991). Microclimatic interactions in agroforestry systems. Forest Ecol. Manage. 45:31-44. |
|
|
Monteiro MM, Vieira DA, Silva-Neto CM, Gatto A, Venturoli F (2016). Abordagem multivariada do uso do hidrogel em espécies nativas do cerrado em área degradada. TreeDim. 1:1-13. |
|
|
Nair PKR (1999). Biogeochemical processes in tropical agroflorestry systems: nutriente cycling. In: Congresso Brasileiro em Sistemas Agroflorestais no Contexto da Qualidade Ambiental e Competitividade, 2. |
|
|
Oliveira ME, Leite LL, Franco AC, Castro LHR (2005). Efeito de duas espécies nativas de árvores sobre as propriedades do solo e forragem de Brachiaria decumbensStapf. Past. Tropicales. 27:1. |
|
|
Ong CK, Corlett JE, Singh RP, Black CR (1991). Above and below ground interactions in agroforestry systems. For. Ecol. Manage. Amsterdam. |
|
|
Poggiani F (2012). Ciclagem de Nutrientes em Florestas do Brasil. In: MARTINS, Sebastião Venâncio. (Editor). Ecologia de florestas Tropicais do Brasil. 2ªed. Universidade Federal de Viçosa, Cap. 6:175-251. |
|
|
Raij BV (1991). Geoquímica de micronutrientes. In: Simpósio sobre micronutrientes na agricultura, 1991, Jaboticabal. Anais. Piracicaba: POTAFOS/CNPq pp. 99-112. |
|
|
Santana GRS, Sant Ana, CER, Silva-Neto CM, Gonçalves BB, Sant'ana LR, Monteiro MM, Ribeiro ACC, Goncalves RA, Castro SS (2016). Microbiological features of dystroferric and dystrophic red oxisols under sugar cane crops subject to different management procedures. Afr. J. Agricult. Res. 11:941-950. |
|
|
Schumacher MV, Brun EJ, Hernandes JI, Konig FG (2004). Produção de serapilheira em uma floresta de Araucaria angustifolia (Bertol.) Kuntze no município de Pinhal Grande-RS. Rev. Árvore. 28(1):29-37. |
|
|
Silva A, Agust S, Costa CA, Carvalho EJM (2015). Avaliação das propriedades físicas de um latossolo amarelo em um cultivo de cumaru em sistema de integração lavoura-pecuária-floresta. Rev. Cient. Elet. Eng. Flor. 26:1. |
|
|
Silva-Júnior MC (2005). 100 árvores do cerrado: guia de campo. Rede de Sementes do Cerrado. Brasília 278 p. |
|
|
Silva-Neto CM, Carneiro VA, Ribeiro ACCM, Oliveira TM, Gonçalves BB (2015). Utilização de resídos de gesso da construção civil para incremento no desenvolvimento de Crotalaria retusa. Brazilian Geographical J.: Geosciences and humanities research medium 6:140-155. |
|
|
Sousa LF, Maurício RM, Gonçalves LC, Sabila EOS, Moreira GR (2007). Produtividade e valor nutritivo da Brachiaria brizantha cv. Marandu em um sistema silvipastoril. Arq. Bras. Med. Vet. Zoot. 59(4):1029-1037. |
|
|
Tedesco MJ, Gianello C, Bissani CA, Bohnen H, Volkweiss SJ (1995). Análise de solos, plantas e outros materiais. Porto Alegre: UFRG, Departamento de Solos/Faculdade de Agronomia. 174 p. Boletim Técnico; 5. |
|
|
Townsend CR, Costa NL, Pereira RGA (2009). Considerações sobre sistemas de integração lavoura-pecuária na Amazônia. Porto Velho, RO: Embrapa Rondônia, 2009 Série Embrapa, Documentos, 130. |
|
|
Upson MA, Burgess PJ, Morison JIL (2016). Soil carbon changes after establishing woodland and agroforestry trees in a grazed pasture. Geoderma 283:10-20. |
|
|
Venturoli F, Carvalho FA, Silva-Neto CM, Oliveira TM, Moraes DC, Souza DM (2015). Manejo Florestal no bioma Cerrado: uma opção para conservar e lucrar. Sci. For. P 43. |
|
|
Vitti GC, Lima E, Cicarone F (2006). Cálcio, Magnésio e Enxofre. In: Fernandes, Manlio Silvestre. (editor). Nutrição mineral de plantas. Sociedade Brasileira de Ciência do Solo. Cap. 12:299-235. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


