Full Length Research Paper

ABSTRACT
The aim of this study was to establish the attributes of kernel type based on kernel hardness and its interrelationship with maize endosperm chemical components, which are essential during storage. Three local varieties and one hybrid variety commonly grown by smallholder farmers in central Malawi were used. For storage experiment, the maize samples were infested with Prostephanus truncatus, larger grain borer (LGB) for eight weeks. Due to the high propensity of maize to cross-pollinate, the kernels were classified into flint and dent kernels to establish the physicochemical properties of the kernel type and variety independently. Local variety (L-3) exhibited the lowest number and weight of damaged grains and the highest adult LGB cadavers. Local variety, L-1 and L-3 had significantly the highest proportion of flint kernels type. Moreover, flint kernels of local varieties showed significantly the highest hardness, and the highest content of protein, total zein, α-zein and zinc (p < 0.05). Furthermore, hardness was significantly and positively correlated with total zein and 14 kDa β-zein content (p < 0.05). Zein in the endosperm, particularly 19 and 22 kDa α-zein mainly contributed to distinct hardness of the local varieties hence may contribute to physical barrier of the kernels against storage pests.
Key words: Maize, local variety, kernel type, hardness, chemical properties, larger grain borer.
INTRODUCTION
Globally, maize (Zea mays L.) is the most produced crop (FAO, 2016). Most developing countries rely on maize as the main staple food and primary source of calories (Smale et al., 2011; Suleiman et al., 2015). Maize is consumed by more than 50% of the population in sub-Saharan Africa (SSA), with an average per capita annual consumption in Malawi being 129 kg, Lesotho at 158 kg, Kenya at 76 kg, Zambia at 118 kg and Zimbabwe at 93 kg (FAO, 2013; Smale and Jayne, 2003). Smallholder farmers rely on rain-fed maize farming; hence, production of the single crop in one season, which necessitates long storage period of about eight months (Maliro and Kandiwa, 2015). Furthermore, they prefer flinty varieties commonly referred to as ‘local’, (Smale and Heisey, 1997) which are locally adapted and traditional maize varieties inherited from the ancestors. In addition, these local varieties are associated with superior performance under abiotic stress conditions and higher grain to flour conversion ratio, than dent, white hybrids (Gilbert et al., 1994).
Maize hardness is an important quality factor to smallholder farmers. Indeed, hard kernels are less susceptible to damage during traditional storage by pests (Smale et al., 2011; Suleiman et al., 2015). Endosperm, the main component of maize kernels, significantly contributes to hardness (Mestres and Matencio, 1993; Blandino et al., 2010) with the most abundant components being starch and protein (Fox and Manley, 2009). Protein plays a significant role in influencing mechanical strength of the endosperm, (Fox and Manley, 2009; Williams et al., 2009) owing to zein, the main maize storage protein (Shewry and Halford, 2002). α-zein, which is composed of 19 and 22 kDa has been associated with vitreous kernels whereas γ-zein, which comprised 16 and 27-kDa has been associated with the floury kernels (Chandrashekar and Mazhar, 1999; Gayral et al., 2016).
Generally, normal starch consists of about 25% amylose (Sandhu and Singh, 2007; Hu et al., 2010). Although amylose is a minor compound of maize endosperm, it plays a role in the formation of a compact starch-protein matrix (Dombrink-Kurtzman and Knutson, 1997) as shown by Gayral et al. (2015) that flint maize have a higher amylose content than dent maize as well as their corresponding vitreous and floury endosperm fractions.
In our previous study, we have shown that the Malawian local varieties, which are mainly orange in color, have superiority in nutrients, cookability, and storability than hybrid varieties (Murayama et al., 2017). The local varieties showed more resistance to Prostephanus truncatus, larger grain borer (LGB) during a 12-week storage experiment than hybrid varieties. Furthermore, the Malawian maize varieties and particularly flinty ‘local’ varieties have been contaminated with dent hybrids due to natural open-pollination over time (Smale and Heisey, 1997) and have different chemical composition 1997) and have different chemical composition (Letchworth and Lambert, 1998).
In light of the aforementioned, the extent of cross-pollination and its effect on maize hardness and endosperm chemical properties, which are crucial in determining maize resistance to storage pest damage, are not well established. In the current study, we elucidated the level of natural cross-pollination among farm-stored maize varieties by smallholder farmers in central Malawi and its effect on the interrelationship of kernel hardness, storability, and endosperm chemical components.
MATERIALS AND METHODS
Maize varieties and experimental field design
Four maize varieties were used in this study. Three local varieties namely white local variety (ACC OU 549), yellow local variety (ACC OU 546-2), and yellow local variety (ACC OU 552-4) obtained from Malawi Plant Genetic Resource Center in Chitedze Agricultural Research Station (CARS) and one hybrid variety (DKC-9089) obtained from DeKalb Genetics Corp., USA (Table 1). To obtain the maize samples, eight plots were prepared as a single factor experiment of the maize varieties without randomization. The trial experiment was conducted in an experimental field in CARS, Lilongwe, Malawi during the 2014/2015 cultivation season. Each plot size was 10 m × 15 m and maize seed spacing between hills was 25 and 75 cm between the rows. A compound fertilizer of 23% N, 21% P and 0% K with urea of 46% N were applied in the ratio 2:3 to attain a standard fertilizer of 92 kg-N/ha as recommended by the government of Malawi (Mutegi et al., 2015). The actual total rainfall in 2014/2015 season was 560.4 mm as compared to the normal expected rainfall of 859.0 mm. Daily minimum and maximum temperatures averaged between 14.0 and 38.2°C, respectively (Messina et al., 2017).

Sample preparation
The maize grains were disinfected by freezing at -30°C for three weeks and thawed at 4°C for one week. Moisture standardization was done in a hot air oven (WFO-700, Tokyo Rikakikai Co., Ltd, Tokyo, Japan) at 40°C to achieve approximately 12.0 ± 1.0% moisture content, wet weight basis. Dried samples were kept at 4°C in an airtight container for one week.
LGB resistance
LGB resistance was evaluated as described by Tefera et al. (2011) and Abebe et al. (2009) with some modifications. Two hundred grams of each maize variety were placed in glass jars (500 mL) with steel screens at the lids (60 mesh), then 30 adult P. truncatus of 0 to 7 days old were introduced into each jar and kept in an incubator (MIR-254, Sanyo Co., Ltd) at 28.0 ± 1.0°C. The number of live adult LGB, adult LGB cadavers and number, and weight of damaged grains were recorded after the fourth and eighth week of incubation. All LGB were separated and removed from the maize by use of sieves with 1.00 and 4.00 mm mesh sizes (IIDA Testing Sieve, IIDA Manufacturing Co. Ltd, Osaka, Japan). The number of damaged grains was determined by randomly selecting 100 grains from the jar and counting all grains tunneled by LGB and their weight was determined in triplicate.
Kernels classification
Classification of maize kernels into flint and dent types was done based on endosperm phenotypic characteristics (Suleiman et al., 2015). Maize flour was prepared by milling with a blender (New Power Mill PM-2005, Osaka Chemical Co., Ltd., Osaka, Japan) and sieved through a 355 µm mesh sieve for chemical analysis.
Physical properties
Kernel hardness was determined using the method described by Blandino et al. (2010). Fifteen maize grains were punctured on the upper lateral surface using a texture analyzer (TA-XT2, Stable Micro System, Godalming, UK) with a flat probe P/2 (diameter 2 mm) and 25 kg load cell. Puncture test was performed at 1 mm/s of puncture speed and kernel punctured at the lateral center side, opposite to the embryo to a depth of 2 mm. Kernel hardness was expressed as the break force calculated in Newton (N).
Chemical properties
Moisture content and crude protein were determined using the standard AOAC methods (AOAC, 2005). Total starch and amylose contents were determined using Megazyme K-TSTA and K-AMYL Assay procedures, respectively (Megazyme International, Bray, Ireland). Total zein was extracted as described by Wallace et al. (1990) and the supernatant representing the total zein fraction was dried at 80°C in Kjeldahl tubes for 4 h and analyzed for nitrogen content by the Kjeldahl method (AOAC, 2005). Zein composition analysis was done by extracting zein using a method described by Wu et al. (2012) with some modifications. Twenty-five mg of flour was transferred to a 2 mL micro-centrifuge tube, then mixed, vortexed with 2 mL of 70% ethanol with 2% 2-mercaptoethanol (v/v), and allowed to stand at room temperature overnight. The mixture was centrifuged at 13,000 rpm for 10 min; 100 µl of the supernatant was transferred into a new tube and mixed with 10 µl of 10% sodium dodecyl sulfate (SDS). The mixture was dried in a rotary vacuum (DNA Speed Vac DNA110, Savant Instruments, INC. Farmingdale, NY), and resuspended in 50 µl of water by vortexing. Further 50 µl of sample reducing buffer, composed of 4% SDS, 10% 2-mercapethanol, 10% sucrose, 0.05% Coomassie blue G-250 and 1.5 M Tris-HCl pH 6.5 was added, and vortexed. Tubes were sealed and boiled for 5 min and frozen overnight. The extracted zein was separated by SDS-PAGE (49.5% T, 6% C mixture) as described by Schägger (2006), the gel scanned at 312 nm (BioInstruments ATTO, AE-6933FXES) and quantification of the bands done using ImageJ 1.50i (National Institutes of Health, Bethesda, Maryland). Mineral contents of magnesium (Mg), phosphorus (P), potassium (K), calcium (Ca) and zinc (Zn) were determined by wet-ashing 100 mg with concentrated sulfuric acid and hydrogen peroxide (2:1 v/v), the resulting solution was diluted 25-fold with deionized water and quantified by an inductively coupled plasma mass spectrometry (ICP; ICPS-8100, Shimadzu Co. Ltd., Japan) in triplicate.
Statistical analysis
Results were subjected to statistical analysis using SPSS for Windows (ver.17.0). A t-test and ANOVA complemented with Tukey’s multiple-range test were used to determine significant differences of LGB resistance parameters for each variety, physicochemical properties, and kernel type. Significant level was established at p ≤ 0.05. Hardness and chemical properties were correlated using Pearson’s correlation.
RESULTS
LGB resistance
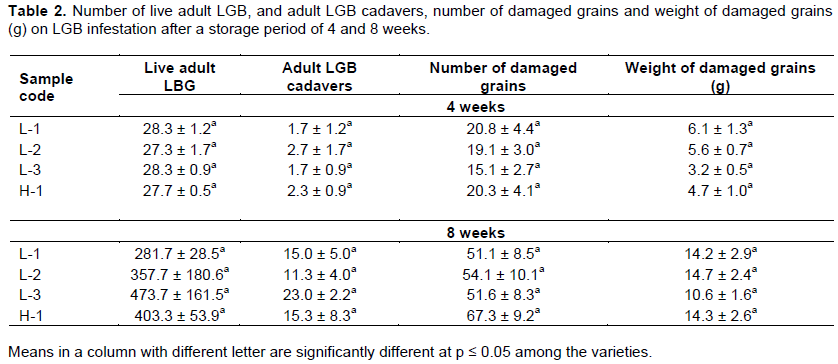
Live and dead insects, damaged grains, and their weight are commonly used to determine maize resistance to LGB in laboratory storage experiments. As shown in Table 2, there was an exponential increase in live adult LGB from the fourth to the eighth week of infestation. In addition, adult LGB cadavers and number, and weight of damaged grains increased with storage period. In the fourth and eighth week of the storage periods, there were no significant differences of the LGB resistance parameters among the varieties (Table 2). However, in the fourth week, L-3 showed the minimum number and weight of damaged grains at 15.1 and 3.2 g, respectively. L-1 and H-1 were observed to have a higher number and weight of damaged grains than L-3. After the eighth week of infestation, L-3 steadily exhibited the minimum number and weight of damaged grains and the highest number of adult LGB cadavers. LGB is known to cause damage to maize grains by tunneling at pre-ingestion phase for food and oviposition during storage period (Li, 1988). The tendency of L-3 to show resistance to damage by LGB during storage due to the less feeding observed, indicates a higher tolerance to insect damage, probably attributed to its physicochemical properties and/or genetic ability. The tendency of H-1 to exhibit the highest number of damaged grains and lower weight damaged grains depicts susceptibility to attack by LGB over the storage period.
Classification
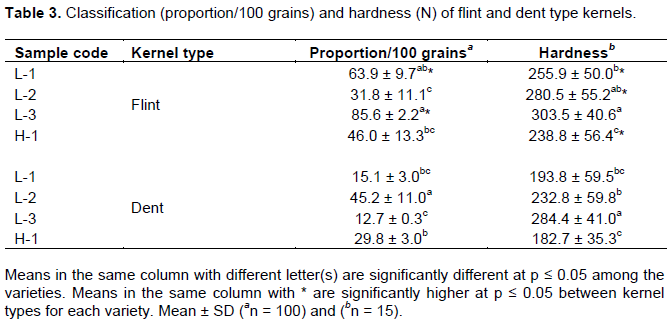
Cross-pollination significantly affects both physical and chemical properties of maize grains (Letchworth and Lambert, 1998). Thus, the maize varieties was classified into flint and dent type kernels based on endosperm phenotypic characteristics (Suleiman et al., 2015)to highlight the interconnection between hardness and chemical composition and their effect on kernel resistance. In every 100 g of maize grains, L-3 and L-1 had significantly the highest proportion of flint type kernels at 85.6 and 63.9%, respectively. Dent type kernel proportion of L-1, L-2, L-3 and H-1 was 15.1, 45.2, 12.7 and 29.8%, respectively (Table 3), while the remaining proportion was classified as semi-flint. Local maize varieties are generally ‘flinty’, which was in agreement with our results. However, the proportion of dent type kernel in local varieties might continue to increase in the near future due to the high degree of maize to cross-pollinate and predominance of hybrid dent varieties among smallholder farmers as well as in the market outlets (Smale and Heisey, 1997). Therefore, there is a need to conserve the ‘local’ flinty maize diversity among the smallholder farmers.
Hardness
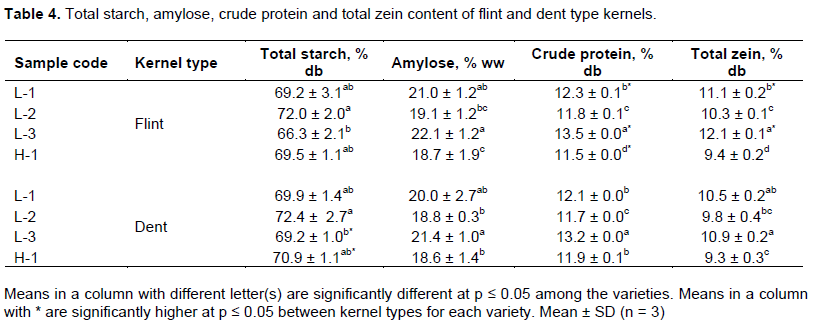
Hardness among the varieties fluctuated significantly between 238.8 and 303.5 N for flint type kernels and 182.7 and 284.4 N for dent type kernels as shown in Table 3. Blandino et al. (2010) reported results within the same range, from 145 to 300 N for 13 commercial maize hybrids. Local varieties had a significantly higher hardness (p < 0.05) compared to Hybrid DKC9089 (H-1), with L-3 showing significantly the highest hardness. For the type of kernels, flint had the significantly higher hardness as compared to dent counterparts apart from L-3, although its flint type kernels had a higher hardness than the dent type kernels. These findings suggest that flint type kernels had a high ratio of vitreous compact endosperm fraction compared with the dent type kernel counterparts. Consequently, this contributed to higher physical resistance of flint type kernels during penetration and shearing by the probe.
Chemical properties
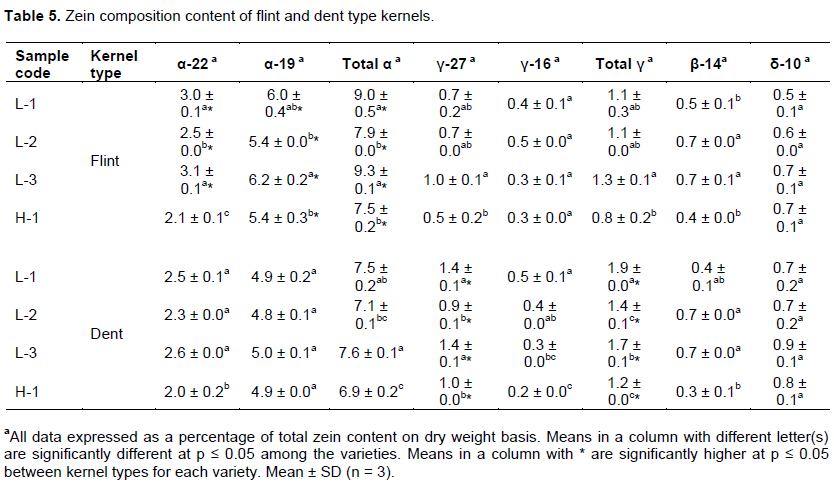
Chemical properties evaluated for flint and dent type kernels of each variety are shown in Table 4. Total starch content among the varieties ranged from 66.3 to 72.0% and 69.2 to 72.4% for flint and dent type kernels, respectively. Essentially, dent type kernels were observed to have a higher content of total starch as compared to flint counterparts (Table 4). Dent type kernels of L-3 and L-1 had significantly the lowest starch content and significantly the highest hardness whereas H-1 had significantly higher starch content and significantly the lowest hardness. These findings are in agreement with other studies that total starch content has a negative correlation with hardness (Pineda-Hidalgo et al., 2015).
Amylose content varied from 18.6 to 22.1% (Table 4) as reported in other studies (Gayral et al., 2015). Local varieties were observed to have significantly higher amylose content than Hybrid variety H-1 (p < 0.05) whereas flint type kernels exhibited the higher content of amylose than dent type kernels (Table 4), which was in agreement with Juárez-García et al. (2013) that vitreous kernels have a higher content of amylose than floury kernels. Amylose gradient exists in the endosperm with a gradual decrease from vitreous to the floury fraction in flint and dent maize (Chandrashekar and Mazhar, 1999; Gayral et al., 2016), thus, the higher amylose content may be responsible for the compactness of starch granules at the periphery of the endosperm as proposed by Dombrink-Kurtzman and Knutson (1997).
The average crude protein for kernel types of each variety are shown in Table 4. Flint type kernels were observed to have higher protein content as compared to their corresponding dent type kernels for all varieties with L-1, L-3 and H-1 having significantly higher protein content (p < 0.05). Despite the low abundance of protein in maize endosperm, its role in constituting the physical resistance in maize kernels cannot be ignored as exhibited by L-3, due to the formation of starch-protein matrix. Indeed, the compactness of the endosperm depends on the architecture of starch (Narvaez-Gonzalez et al., 2006)which is influenced by the presence of proteins leading to the formation of a starch-protein matrix (Wallace et al., 1990).
Further studies with a key interest of zein’s role in kernel hardness were conducted. Total zein accounted for an average of 87.4 and 82.7% of protein content for flint and dent type kernels, respectively. Total zein content among the varieties varied from 9.4 to 12.1% and 9.3 to 10.9% for flint and dent type kernels, respectively as shown in Table 4. Similar to protein content, flint type kernels had higher total zein content compared to dent counterparts with L-1 and L-3 having significantly higher total zein content (p < 0.05). It is important to note that zein synthesis takes place in the endoplasmic reticulum, assembled as protein bodies, and deposited in the endosperm, (Holding and Larkins, 2006; Shewry and Halford, 2002)while Pereira et al. (2008)established that hard-endosperm maize cultivars had abundant and well-organized protein bodies. This may augment the adhesion of protein bodies on the surface of the starch granules hence increasing the rigidity of the starch-protein matrix, which explains the significantly higher hardness of the flint type kernels. It is worth noting that non-zein protein content such as albumins, glutelins, and globulins in dent type kernels of H-1 was the highest at around 22.0% of crude protein, which explains H-1 having significantly the lowest total zein content, yet a higher crude protein content. Furthermore, dent type kernels of the H-1 had significantly the lowest hardness.
To evaluate the composition of zein sub-classes in connection with kernel hardness, extracted zein was separated by SDS-PAGE and quantified as a percentage of total zein as shown in Table 5. Distinct bands observed in the gels were similar to previous studies of maize kernels (Wu et al., 2012; Paiva et al., 1991). α-zein was the most abundant zein class with an average of 78.7% and 75.4% of total zein content for flint and dent type kernels, respectively. Notably, 19 kDa was higher than 22 kDa for α-zein content, while 27 kDa content was higher than 16 kDa for γ-zein content. Flint type kernels had significantly higher total α-zein and 19 kDa α-zein compared to dent type kernel for each variety (p < 0.05), whereas dent type kernels had a significantly higher total γ-zein and 27 kDa γ-zein content (p < 0.05). This is in agreement with Gayral et al. (2016)who established that γ-zein was significantly higher in floury endosperm and augments our earlier findings that dent kernel had a higher content of total starch. γ-zein and the most expressed 27 kDa γ-zein (Holding and Larkins, 2006)are synthesized during early stages of kernel development and as a result of programmed cell death (PCD) (Young and Gallie, 2000)are associated with the inner core floury fraction of endosperm. In addition, more mature protein bodies, mainly composed of α-zein are located at the periphery of endosperm due to PCD (Gayral et al., 2016; Holding and Larkins, 2006)which explains the high content of α-zein in flint kernels.
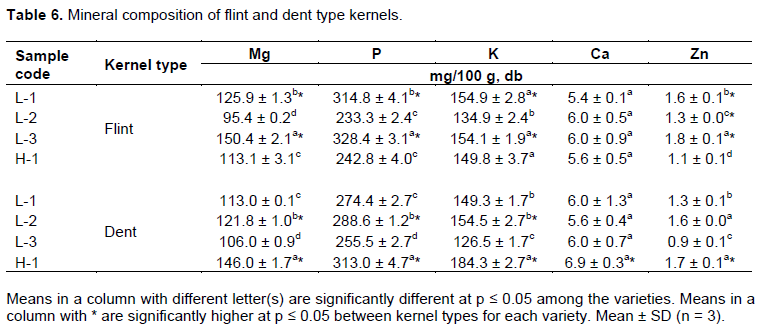
The main minerals in maize kernels such as Mg, K, P, Ca and Zn are shown in Table 6. The most abundant mineral was P, ranging from 242.8 to 328.4 mg/100 g dry weight (dw) while Zn was the least ranging from 1.8 to 0.9 mg/100 g dw in both kernel types. Mineral content was similar with other studies following P, K, Mg, Ca, and Zn descending order (Chen et al., 2016; Nuss and Tanumihardjo, 2010). Maize endosperm has a negligible <1% of minerals in the grain, while 30% of zinc is localized in the endosperm (Nuss and Tanumihardjo, 2010). The concentration of zinc at a high level may be toxic to grain storage insects by impairing catalase mechanism (Lazzari and Lazzari, 2012). Moreover, for normal growth and a healthy life, smallholder farmers require maize rich in microelements such as zinc (Ortiz-Monasterio et al., 2007)considering the high prevalence of zinc deficiency in Malawi (Siyame et al., 2013). In this study, it was observed that flint type kernels had significantly higher content of zinc, which can help improve the zinc deficiency problem as well as act as an intrinsic deterrent of LGB during storage of maize.
Correlation between hardness and chemical composition
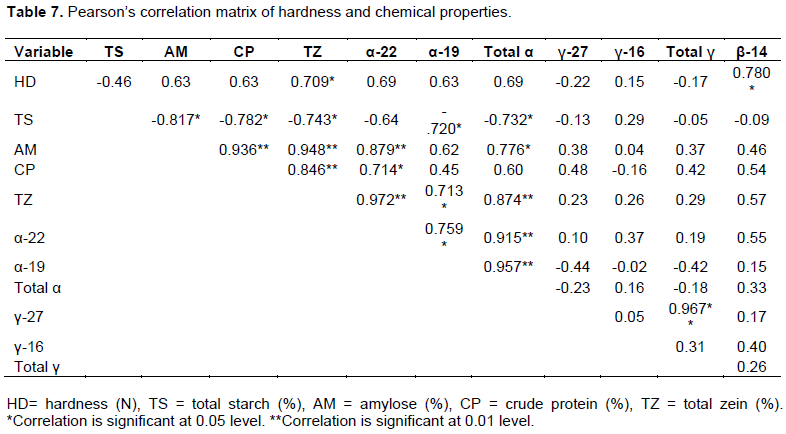
Pearson’s correlation results of hardness, chemical properties and LGB resistance parameters at the eighth week of infestation are shown in Table 7. Total zein content, the abundant protein in maize endosperm was positively correlated with hardness at r = 0.71 (p < 0.05). This finding shows that total zein is strongly associated with hardness and directly influences the physical resistance of maize kernels. It is worthwhile noting that, greater than 70% of zein is composed of α-zein (Wu et al., 2012)and actually, flint type kernels had significantly higher hardness than their dent counterparts, and therefore α-zein and particularly 19 kDa α-zein certainly plays a major role in kernel hardness as shown by our ANOVA results. Furthermore, 14 kDa β-zein was positively correlated with hardness at r = 0.78 (p < 0.05). β-zein is located in the periphery of the protein bodies (Shewry and Halford, 2002) and contains methionine and cysteine hence rich in sulfur (Wu et al., 2012). L-3 showed significantly the highest 14 kDa β-zein content, whereas H-1 had significantly the lowest content. Indeed, it can be hypothesized that, probably due to the presence of methionine and cysteine, β-zein could contain a higher disulfide bridges hence increasing the rigidity of the protein bodies at the periphery, which might impart better filling of the starch intergranular spaces, therefore higher compactness and vitreousness of the endosperm.
CONCLUSION
Based on this study, the tendency of local varieties to have resitance to LGB, especially L-3 was due to its physicochemical properties despite of natural transgene effect of cross-pollination, further depicting the preference of local varieties among smallholder farmers in Malawi. The hardness of maize kernel was highly influenced by both the composition of starch and protein in the endosperm. In this study, it has been shown that zein profile was more associated with hardness, and therefore regulating its synthesis may increase endosperm vitreousness. Consequently, L-3 exhibited highest proportion of flint type kernels and superiority in its hardness, which was closely associated with its chemical properties. These chemical properties depend on genetic traits as observed in variety and kernel type variables, although abiotic factors may also have an influence. L-3 may be proposed as a suitable farm-stored maize variety for smallholder farmers in Malawi, and highlights the importance of conserving the local flint maize varieties. Currently, breeding programs endeavour to develop insect-resistant varieties to minimize postharvest losses due to storage pests. Plant breeders may utilize local varieties like L-3 as a genetic resource to develop naturally superior pest resistant farm-stored maize cultivars for smallholder farmers in Malawi and sub-Saharan Africa (SSA) reducing the current high post-harvest losses caused by LGB during storage.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
The authors thank Mr. Arsenio Chimphamba and Ms. Mary Chatambalala in the Chitedze Agricultural Research Station for field management during cultivation of the maize samples. They also thank Chitdze Agricultural Research Station (CARS), Lilongwe, Malawi for supporting the field experiment. This work was supported by Japan Society for the Promotion of Science (JSPS) KAKEN Grant Number 26304022.
REFERENCES
|
Abebe F, Tefera T, Mugo S, Beyene Y, Vidal S (2009). Resistance of maize varieties to the maize weevil Sitophilus zeamais (Motsch.) (Coleoptera: Curculionidae). African Journal of Biotechnology 8(21):5937-5943. |
|
|
AOAC (2005). Official methods of analysis of AOAC International, 18th ed. AOAC International, Gaithersburg, MD, USA. |
|
|
Blandino M, Mancini M, Peila A, Rolle L, Vanara F, Reyneri A (2010). Determination of maize kernel hardness: Comparison of different laboratory tests to predict dry-milling performance. Journal of the Science of Food and Agriculture 90(11):1870-1878. |
|
|
Chandrashekar A, Mazhar H (1999). The biochemical basis and implications of grain strength in sorghum and maize. Journal of Cereal Science 30(3):193-207. |
|
|
Chen Q, Mu X, Chen F, Yuan L, Mi G (2016). Dynamic change of mineral nutrient content in different plant organs during the grain filling stage in maize grown under contrasting nitrogen supply. European Journal of Agronomy 80:137-153. |
|
|
Dombrink-Kurtzman M, Knutson C (1997). A study of maize endosperm hardness in relation to amylose content and susceptibility to damage. Cereal Chemistry 74(6):776-780. |
|
|
Food and Agriculture Organization (FAO) (2013). FAOSTAT The Food and Agriculture Organization of the United Nations. |
|
|
Food and Agriculture Organization (FAO) (2016). FAOSTAT The Food and Agriculture Organization of the United Nations. |
|
|
Fox G, Manley M (2009). Hardness methods for testing maize kernels. Journal of Agricultural and Food Chemistry 57(13):5647-5657. |
|
|
Gayral M, Bakan B, Dalgalarrondo M, Elmorjani K, Delluc C, Brunet S, Linossier L, Morel MH, Marion D (2015). Lipid partitioning in maize (Zea mays L.) endosperm highlights relationships among starch lipids, amylose, and vitreousness. Journal of Agricultural and Food Chemistry 63(13):3551-3558. |
|
|
Gayral M, Gaillard C, Bakan B, Dalgalarrondo M, Elmorjani K, Delluc C, Brunet S, Linossier L, Morel M, Marion D (2016). Transition from vitreous to floury endosperm in maize (Zea mays L.) kernels is related to protein and starch gradients. Journal of Cereal Science 68:148-154. |
|
|
Gilbert E, Phillips L, Roberts W, Sarch MT, Smale M, Stround A, Hunting E (1994). Maize research impact in Africa: The obscured revolution. |
|
|
Holding D, Larkins B (2006). The development and importance of zein protein bodies in maize endosperm. Maydica 51(2):243-254. |
|
|
Juárez-García E, Agama-Acevedo E, Gómez-Montiel N, Pando-Robles V, Bello-Pérez L (2013). Proteomic analysis of the enzymes involved in the starch biosynthesis of maize with different endosperm type and characterization of the starch. Journal of the Science of Food and Agriculture 93(11):2660-2668. |
|
|
Lazzari S, Lazzari F (2012). Insects pests in stored grain. In Panizzi R, Paria R (Eds.), Insect Bioecology and Nutrition for Intergrated Pest Management. Florida, FL: CRC Press pp. 471-450 |
|
|
Letchworth M, Lambert R (1998). Pollen parent effects on oil, protein, and starch concentration in maize kernels. Crop Science 38(2):363-367. |
|
|
Li L (1988). Behavioural ecology and life history evolution of the Larger Grain Borer, Prostephanus truncatus (Horn). Ph.D. Thesis, University of Reading, Reading, UK. |
|
|
Maliro D, Kandiwa V (2015). Gender analysis of maize post-harvest management in Malawi: A case study of Lilongwe and Mchinji districts. |
|
|
Messina J, Snapp S, Peter B (2017). Malawi weather station data for the 2014-2015 growing season. Mendeley, . |
|
|
Mestres C, Matencio F (1993). Biochemical basis of kernel milling characteristics and endosperm vitreousness of maize. Journal of Cereal Science 24:283-290. |
|
|
Murayama D, Tomoka Y, Chandiona M, Ephantus NB, Rodney LG, Jiwan PP, Masayuki T, Hiroshi K, Daigo A (2017). Superiority of Malawian orange local maize variety in nutrients, cookability and storability. African Journal of Agricultural Research 12(19):1618-1628. |
|
|
Mutegi J, Kabambe V, Zingore S, Harawa R, Wairegi L (2015). The status of fertilizer recommendation in Malawi: Gaps, challenges, opportunities and guidelines: Soil Health Consortium of Malawi. |
|
|
Narvaez-Gonzalez E, Figueroa-Cardenas J, Taba S, Tostado E, Peniche R, Sanchez F (2006). Relationships between the microstructure, physical features, and chemical composition of different maize accessions from Latin America. Cereal Chemistry 83(6):595-604. |
|
|
Nuss E, Tanumihardjo S (2010). Maize: A paramount staple crop in the context of global nutrition. Comprehensive Reviews in Food Science and Food Safety 9(4):417-436. |
|
|
Ortiz-Monasterio JI, Palacios-Rojas N, Meng E, Pixley K, Trethowan R, Peña RJ (2007). Enhancing the mineral and vitamin content of wheat and maize through plant breeding. Journal of Cereal Science 46(3):293-307. |
|
|
Paiva E, Kriz A, Peixoto M, Wallace J, Larkins B (1991). Quantitation and distribution of y-zein in the endosperm of maize kernels. Cereal Chemistry 68(3):276-79. |
|
|
Pereira R, Davide L, Pedrozo C, Carneiro N, Souza I, Paiva E (2008). Relationship between structural and biochemical characteristics and texture of corn grains. Genetics and Molecular Research 7(2):498-508. |
|
|
Pineda-Hidalgo K, Vega-Alvarez E, Calderon-Zamora L, Salazar-Salas N, Gutierrez-Dorado R, Reyes-Moreno C, Bello-Perez L, Lopez-Valenzuela J (2015). Physicochemical, structural, and proteomic analysis of starch granules from maize landraces of Northwest Mexico. Cereal Chemistry 92(3):320-326. |
|
|
Sandhu KS, Singh N (2007). Some properties of corn starches II: Physicochemical, gelatinization, retrogradation, pasting and gel textural properties. Food Chemistry 101(4):1499-1507. |
|
|
Schägger H (2006). Tricine - SDS-PAGE 1(1):16-23. |
|
|
Shewry P, Halford N (2002). Cereal seed storage proteins: structures, properties and role in grain utilization. Journal of Experimental Botany 53(370):947-958. |
|
|
Siyame E, Hurst R, Anna W, Young S, Broadley M, Chilimba A, Ander L, Watta M, Chilima B, Gondwe J, Dalitso K, Kalimbira A, Fairweather-Tait S, Bailey K, Gibson R (2013). A high prevalence of zinc but not iron deficiency among women in rural Malawi: a cross-sectional study. International Journal for Vitamin and Nutrition Research 83(3):176-187. |
|
|
Smale M, Byerlee D, Jayne T (2011). Maize Revolutions in Sub-Saharan Africa. Policy Research Working Paper 5659, Agriculture and Rural Development Team, World Bank. |
|
|
Smale M, Heisey P (1997). Maize technology and productivity in Malawi. In: Byerlee D, Eicher C (Eds.), Africa's Emerging Maize Revolution. Colorado, CO: Lynne Rienner Publishers. pp. 63-81 |
|
|
Smale M, Jayne T (2003). Maize in Eastern and Southern Africa: "Seeds" of Success in Retrospect. EPID Discussion Paper No. 97, International Food Policy Research Institute. |
|
|
Suleiman R, Williams D, Nissen A, Bern C, Rosentrater K (2015). Is flint corn naturally resistant to Sitophilus zeamais infestation? Journal of Stored Products Research 60:19-24. |
|
|
Tefera T, Mugo S, Likhayo P (2011). Effects of insect population density and storage time on grain damage and weight loss in maize due to the maize weevil Sitophilus zeamais and the larger grain. African Journal of Agricultural Research, 6(10):2249-2254. |
|
|
Wallace J, Lopes M, Paiva E, Larkins B (1990). New methods for extraction and quantitation of zeins reveal a high content of y-zein in modified opaque-2 maize. Plant Physiology 92:191-196. |
|
|
Williams P, Geladi P, Fox G, Manley M (2009). Maize kernel hardness classification by near infrared (NIR) hyperspectral imaging and multivariate data analysis. Analytica Chimica Acta 653(2):121-130. |
|
|
Wu Y, Wang W, Messing J (2012). Balancing of sulfur storage in maize seed. BMC Plant Biology 12(77):1471-2229. |
|
|
Young T, Gallie D (2000). Programmed cell death during endosperm development. Plant Molecular Biology 44(3):283-301. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


