The aim of this work was to find a natural product and efficient dose to control early blight in tomato crop in agro-ecological or conventional agriculture system. The treatments were essential oils from Cymbopogon citratus, Eucalyptus citriodora, Piper hispidinervum, Cymbopogon martini, Rosmarinus officinalis, Syzygium aromaticum, Cinnamomum zeylanicum, Mentha piperita, Citrus sinensis var. dulcis, and Melaleuca alternifolia used against Alternaria solani in Solanum lycopersicum under greenhouse conditions. In vitro, mycelial growth and conidia germination were completely inhibited by C. zeylanicum, C. martini, C. citratus, and S. aromaticum oils of 750 μL L-1, E. citriodora and M. alternifolia at 2000 μL L-1, and Mentha piperita in 5000 μL L-1. In tomato, plants under greenhouse conditions, early blight in leaf was inhibited by C. zeylanicum, C. martini, C. citratus, and S. aromaticum at 750 μL L-1, and E. citriodora oil of 5000 μL L-1. The best results for MIC 50 and 90 with minor dose to control was essential oil from C. citratus at 341.32 and 1.822.10 µL L-1 rich in geranial (46.91%), neral (34.34%), and geranyl acetate (6.30%). Therefore, it is possible to use this essential oils or manufacture a new and efficient product to control Alternaria solani in tomato plants.
Plant diseases are caused by fungi, nematodes, bacteria, and viruses, among which fungi are the main pathogens, causing great yield losses in numerous important crops (Huang et al., 2010). Natural sources from plants play a significant role in the prevention and control of these diseases. In addition, products of higher plants may lead to the discovery of the source of antimicrobial agents with possible novel mechanisms of action (Hada, 2014). Tomato (Solanun lycopersicon Mill.) is one of the most economically important vegetables in the world (Kurozawa and Pavan, 2011, 2005; Filgueira, 2000). It is the host for a wide spectrum of pests and diseases and thus requires specific ecological, nutritional and phytosanitary conditions in cropping and/or field (Leite et al., 2003). At post harvesting conditions, it is a highly perishable vegetable with a short shelf-life and high susceptibility to fungal disease during prolonged storage (Ibrahim, 2014).
Early blight, which is caused by Alternaria solani (Ell. and Martin) Jones and Grout, is one of the most important diseases affecting tomato plants, since it causes direct (fruits) and indirect (branches and leaves) damages (Yanar, 2011; Leite et al., 2003). Fungicides including mancozeb, chlorothalonil, thiophanate-methyl and copper are used to control this disease.
Losses of up to 75% of agrochemicals during application have been recorded for tomato crops. Nowadays, producers use 45 agrochemical applications during the cycle of this species. The indiscriminate use of agrochemicals in tomato plants, associated with the lack of knowledge by producers, results in the development and intensification of occupational diseases and environmental contamination besides exposure of consumers to such risk (Araújo at al., 2000; Kishore and Pande, 2007; Fawzi et al., 2009).
Natural products have great potential for the management of pests and diseases, among other utilizations from extracts and essential oils (Zanella, 2015). Historically, the origin of pyrethroids and carbamates is an example of the potential of natural products for phytosanitary uses.
In this context, essential oils from plants have great potential due to their intrinsic characteristics such as high bioactivity, great molecule diversity and volatility, and brief environmental persistence (Knaak and Fiuza, 2010; Lee et al., 2008). The mechanism of action of essential oils in live cells is related to cell membrane permeability alteration, enzyme synthesis inhibition and/or inactivation (Souza et al., 2005). Cruz et al. (2015) had significant results in essential oils from Plectranthus amboinicus leaves against Fusarium solani at 10 μL. In Pyricularia grisea, Cymbopogon winterianus has total mycelia growth inhibition with 10 μL (Perini et al., 2013).
According to Soylu et al. (2010) and Copping and Duke (2007), based on the activity and mechanism of action concerning natural products, they may be directly applied on plants for protection and have potential applicability in the establishment of programs for the development of new products as well as synthesis and/or semi-synthesis processes. Also, Pawar and Thaker (2007) studied the antifungal effect of essential oils from 75 plant species against Alternaria porri and Fusarium oxysporum f.sp. ciceris. Yanar et al. (2011) studied the antifungal effect of essential oils from 27 plant extracts, and Sallam and Kamal (2012) studied six plants. These authors emphasized the potentiality and importance of researching these compounds as an alternative for synthetic phytosanitary products, considering their economic and environmental viability.
Several authors have reported the efficiency and wide spectrum of action of essential oils against insects, fungi and pathogenic and phytopathogenic bacteria, both in vitro and in vivo (Paes, 2012,). Thus, the present work aimed at advancing applied research by evaluating, through preventive applications, the effect of different doses of essential oils from 10 plant species on conidium germination and mycelial growth in vitro as well as their controlling action, in vivo, on the severity of early blight caused by A. solani in the leaves of S. lycopersicum Mill. cv. Sta. Clara cultivated under greenhouse conditions.
Since the present paper is composed of three different linked works, the result was adopted as eliminatory parameters of treatment efficiency for the subsequent studies.
The employed essential oils are of commercial origin and obtained through hydrodistillation. In all experiments, essential oils were homogenized with a 1:1 mixture of water and Tween 80 detergent. The used concentrations were calculated based on the essential oil.
The essential oils analysis
Analysis of essential oil components was analyzed through Gas Chromatography, ubMass Molecular Biology and Phytochemistry, Agronomical Institute of Campinas, São Paulo State, Brazil.
A 2 mg oil sample was diluted in 1 ml ethyl acetate (HPLC grade), from which a 1 μL aliquot was injected. The analysis was carried out in a GC/MS - Shimadzu/QP-5000, equipped with DB-5 column (30 m × 0.25 mm × 0.25 μm) electron ionization (70 eV) and mass scan range from 30 to 300 Da. Helium was used as carrier gas at 1.0 ml/min flow, and injector at 240ËšC. The following program was used: 50ËšC (5’) to 160ËšC, 3ËšC per minute, and 160 to 220ËšC, 10ËšC per minute. Using a split ratio of 35, the temperatures of the ion source and GC-MS interface were 200 and 230°C, respectively. Compounds were identified by comparing their mass spectra with the GC/MS spectral library. Kovats retention index was calculated and determined by comparing the data with those found in literature (Adams, 1995).
Effect of essential oils at different doses on A. solani mycelial growth in vitro (Experiment 1)
Mycelia were obtained in the Mycology and Forest Phytopathology Laboratory, Department of Plant Production, College of Agronomical Sciences ,SãoPaulo Stat eUniversity-UNESP, Botucatu, São Paulo State, Brazil.
The experiment was carried out in Petri plates. The plates with each treatment were organized in a completely randomized design of 10 × 6 factorial arrangement, with five replicates. The treatments were essential oils from the following species: Cymbopogon citratus (lemon grass), Eucalyptus citriodora (lemon eucalyptus), Piper hispidinervum (long pepper), Cymbopogon martini (palmarosa), Rosmarinus officinalis (rosemary), Syzygium aromaticum (clove), Cinnamomum zeylanicum (cinnamon), Mentha piperita (peppermint), Citrus sinensis var. dulcis (orange), and Melaleuca alternifolia (tea tree), at 5 different concentrations (0, 250, 500, 750, 1000 and 5000 μL L-1), except for M. alternifolia, M. piperita and Eucaliptus. citriodora oils, which were used at 0, 1000, 2000, 3000, 4000 and 5000 μL L-1. After autoclaving, treatments were added to PDA culture medium. Plates were inoculated with 0.5 cm diameter disks and kept in BOD chamber at 25ºC in the dark.
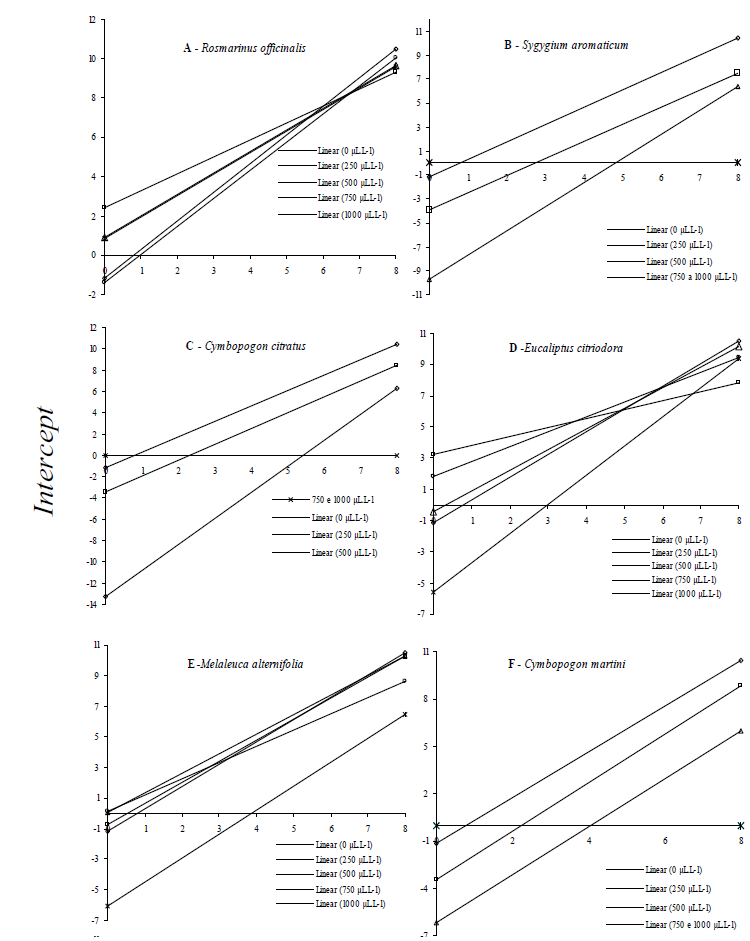
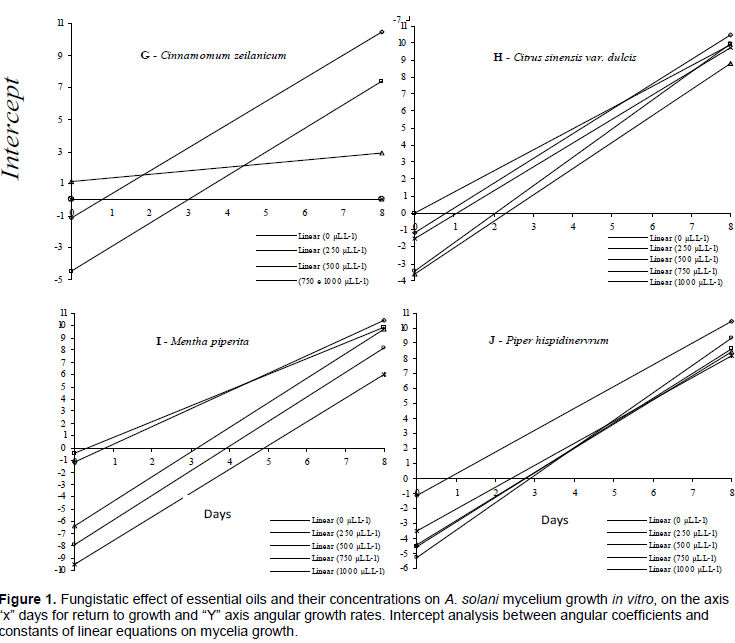
The colony diameter (mm) was daily evaluated until the eighth day. The mean of the ratio between measures in two perpendicular directions considered the growth value. The growth rate over time was assessed through parallelism test among doses of each oil, in which angular coefficients and constants from the growth linear regression equations were calculated. The results were plotted and the fungi static effect was analyzed through intercept (angular coefficient from the progress equation by each oil and dose), in which, the lower value, the smaller will be the angular coefficient of the disease symptom progression rate, mycelial growth and conidia germination at each treatment. Thus, each angular coefficient from the linear regression analysis contrasted the Scott-Knott method (p ≤ 0.0001).
To obtain the maximum inhibitory concentrations (50 and 90%), differential and integral calculus was applied. Regression statistical test was performed using "F" test.
Effect of essential oils at different doses on A. solani conidium germination in vitro (Experiment 2)
Conidia were obtained at Laboratory of Mycology and Forest Phytopathology, College of Agronomic Sciences, UNESP, Botucatu, São Paulo State, Brazil.
The experiment was carried out in 2% Agar medium in water. The design is completely randomized in 5 × 5 factorial arrangement, with three replicates. Treatments consisted of five essential oils selected among those from Experiment 1 (C. citratus, C. martini, S. aromaticum, C. zeylanicum, and E. citriodora). Each oil was tested at five concentrations (0, 250, 500, 750, and 1000 μL L-1). The culture medium containing the treatments was applied onto glass slides placed in Petri plates and inoculated with 10 mL of a suspension containing 103 μL-1 A. solani conidia. The plates were kept in a Biological organism development chamber (BOD) for 24 h at 25oC in the absence of light.
Conidium germination was evaluated with the aid of an optical microscope. The conidium that presented germ tube extension higher than the analyzed conidium length was considered germinated (Neely, 1978), and the germination percentage was calculated. Due to the pattern of conidium germination in response to treatments, only analysis of variance and mean contrasts were done through the Scott-Knott method (p ≤ 0.0001), to the detriment of regression analysis.
Effect of essential oils at different doses on the control of early blight caused by A. solani in the leaves of S. lycopersicon Mill. cv. Sta. Clara under greenhouse cropping (Experiment 3)
S. lycopersicon Mill. cv. Santa Clara seedlings (30 days old) was transplanted to 13-L pots. The substrate consisted of a mixture of earth, sand and commercial organic substrate (1:1:1), in addition to 10 kg fertilizer formula 4-14-08 per m3 of the mixture. Plants were kept for 57 days after transplanting (DAT) in a high tunnel under greenhouse condition; they were covered with 150 µ “Agrofilme” and subjected to daily irrigation through micro-aspersion (5 mm flow). The experimental design was in randomized blocks in 5 × 4 factorial arrangement, with 6 replicates; 5 essential oils (C. citratus, E. citriodora, C. martini, S. aromaticum, and C. zeylanicum) were used at 4 concentrations (0, 500, 750, and 1000 μL L-1), except for E. citriodora (0, 750, 1000 and 5000 μL L-1).
The pathogen inoculation consisted of the application of a 104 A. solani conidia/mL suspension twice on consecutive days after the first application of treatments, which was repeated at 3-day intervals until 57 DAT. It was done with the aid of a handheld backpack sprayer and plastic blanket for isolation of plots. At 57 DAT, the symptom severity was evaluated in the leaf tissue. The evaluator received a visual accuracy training with the software DISTRAN (public domain) of diagrammatic scale of damaged leaf area symptoms (Grade 0 = 0; 1 = 1-25%; 2 = 26-50%; 3 = 51-75%; and 4 = 76-100% injured leaf area) (Azevedo, 1997). Nine leaves from basal, middle and apical part of the canopy in ten plants were evaluated; from each replicate we can calculate the index control of relative efficiency (EFCR). The EFCR was calculated in relation to each standard treatment per treatments and replications (Ming et al., 2012)
Data were subjected to regression analysis (p ≤ 0.0001) with square root equations and Minimal Inhibitory Concentration (MIC) of 50 and 90% of symptoms of plant disease was determined. We do this study only in greenhouse field experiment with better oils after in vitro conidium and mycelial experiments.
Chemical composition of the essential oils and their potential use
The results of GC/MS analysis from the essential oils tested and the activity reported on literature for each one and/or the most important compound detected are presented. The majority of antimicrobial compounds found in essential oils are terpenoids and phenylpropenes with the most active being phenols, although some aldehydes and non-phenolic substances also present promising antimicrobial activity. The target site and mode of action of most essential oil components still under is not well understood, especially in yeast (Hyldgaard, 2012).
The GC/MS results from the essential oils used in this work indicated that: The C. citratus (lemon grass) essential oil has the major constituents of geranial (46.91%), neral (34.34%), geranyl acetate (6.30%), camphene (1.02%), 6-methyl-5-hepten-2-one (1.28%), linalool (0.82%), citronellal (0.25%), isomenthol (1.81%), nerol (0.45%), geraniol (3.52%), and α-trans-bergamotene (0.85%).
C. martini (palmarosa) essential oil had major component of geraniol (86.98%), followed by geranyl acetate (9.03%), trans-ocimene (1.45%), trans-caryophyllene (0.63%), and geranial (0.41%). Geranial and neral aldehyde mixture constitutes citral, a potent bioactive substance against phytopathogens (GlamoÄlija et al., 2011).
E. citriodora (eucalyptus) essential oil main compound was citronellal (74.74%), followed by citronellol (6.26%), isopulegol (5.18%), iso-isopulegol (4.33%), geraniol (2.96%), α-pinene (0.45%), β-pinene (0.68%), limonene (0.33%), 1,8-cineole (1.51%), citronellyl acetate (1.24%), geranyl acetate (0.28%), and trans-caryophyllene (0.72%).
Both S. aromaticum (clove) and C. zeylanicum (cinnamon) had the major compound, eugenol: it is a major constituent in clove essential oil, and its antimicrobial activity is linked to its ability to permeabilize the cell membrane and interact with proteins; a phenylpropanoid of high antibiotic efficiency, specially against fungi; it can be used in crop phytopathogen control, as well as in food conservation. The antifungal mode of action of eugenol needs further investigation, but it is known to depend on cell proliferation. Eugenol treatment altered cell membrane and cell wall structures of proliferating S. cerevisiae cells resulting in the release of cellular content (Hyldgaard et al., 2012; Faria et al., 2006; Souza et al., 2005).
S. aromaticum had higher eugenol content (82.55%), relative to that of C. zeylanicum (73.45%). It had less diversity of substances - trans-caryophyllene (11.98%), eugenyl acetate (3.13%), α-humulene (1.74), and α-copaene (0.59%) compared with those present in C. zeylanicum essential oil: Cis-caryophyllene (4.67%), linalool (3.58%), benzyl benzoate (2.92%), cis-cinnamyl acetate (2.11%), safrole (1.25%), α-pinene (1.24%), α-phellandrene (1.32%), ortho-cymene (1.11%), β-phellandrene (1.10%), camphene (0.40%), β-pinene (0.47%), α-terpineol (0.36%), trans-cinnamaldehyde (0.77%), and α-copaene (0.75%). However, Pawar and Thaker (2007) attributed the antifungal activity of C. zeylanicum essential oil to cinnamaldehydes.
M. alternifolia (tea tree) had 4-terpineol (46.74%) and gamma-terpinene (18.16%) as the main components; it also presented α-terpinene (7.5%), 1,8-cineole (5.25%), ortho-cymene (4.38%), α-terpineol (3.43%), terpinolene (2.84%), α-pinene (2.32%, limonene (1.69%), α-selinene (1.27%), aromadendrene (1.00%), gamma-cadinene (0.99%), myrcene (0.82%), β-pinene (0.80%), sabinene (0.41%), trans-caryophyllene (0.37%), and delta-3-carene (0.34%).
P. hispidinervum (long pepper) presented Safrole with major constituent (93.85%) in the essential oil; it had α -pinene (0.35%), myrcene (0.18%), delta-3-carene (0.41%), ortho-cymene (0.14%), limonene (0.21%), cis-ocimene (0.35%), trans-ocimene (0.88%), terpinolene (2.37%), trans-caryophyllene (0.30%), bicyclogermacrene (0.62%), and pentadecane (0.25%). In this oil, the major compound, Safrole detected has high relevance to industrial and agriculture chemical use. It has approximately 90 to 94% of the essential oil compound produced by the plant; it is applied as synergistic element in the composition of insecticides and herbicides (Maia et al., 1987).
For R. officinalis (rosemary), the essential oil is mainly composed of 1,8-cineole (25.51%), α -pinene (22.23%), camphor (18.64%), camphene (10.63%), limonene (5.51%), β -pinene (4.76%), borneol (2.48%), myrcene (2.39%), 3-octanone (0.44%), ortho-cymene (1.87%), bornyl acetate (1.80%), α -terpineol (1.40%), trans-caryophyllene (1.15%), and linalool (0.92%). Rosemary oil, until, has no agriculture use. M. piperita (peppermint) essential oil main constituent was menthol (45.18%), followed by menthone (22.54%), 1,8-cineole (7.03%), meta-cresol acetate (6.39%), menthyl acetate (5.97%), neo-menthol (2.88%), limonene (1.79%), pulegone (1.55%), trans-caryophyllene (1.21%), β-pinene (1.12%), non-identified compounds (0.72%), gamma-terpinene (0.70%), α-pinene (0.60%), gamma-muurolene (0.59%), isomenthol (0.47%), sabinene (0.38%), 3-octanol (0.25%), ortho-cymene (0.25%), trans- β -ocimene (0.25%), and myrcene (0.12%).
For Citrus sinensis var. dulcis (orange) essential oil used in this work is limonene (96.86%); it also presents small quantities of myrcene (2.27%) and α-pinene (0.52%).
Considering the effect attributed to the essential oil major compound, this relation is almost direct when the oil has little diversity of molecules. It is important to point out the major compounds of essential oils, since they may be responsible, at least in part, for the oil antifungal activity (Salamci et al., 2007). However, is not true for all the essential oils and/ or microorganisms.
For essential oils with great diversity of molecules that is well distributed, the synergistic effect hypothesis is probably the most suitable. In some cases, large proportions of oxygenated monoterpenes may guarantee the essential oil antifungal activity (Santana, 2015).
The mechanism of action of essential oils in live cells is related to cell membrane permeability alteration, and enzyme synthesis inhibition and/or inactivation (Souza et al., According to the parallelism test for the growth curves of the analyzed doses of essential oils, R. officinalis, C. sinensis v. dulcis, and P. hispidinervum did not significantly reduce mycelial growth rate, presenting thus the highest angular coefficients (Table 1).

However, it can be seen that there are oils at different concentrations where the growth remains stagnant when the intercept remains at the abscissas (Table 1 and Figure 1). In this case, there is only fungistatic action and not fungicide effect. Aside oil type or dose, regular application is needed. However, such characteristics concerning monoterpenes constitute a dubious quality for the applicability of essential oils in crop protection, since they are highly volatile and instable. Commercial applications of essential oils would benefit from deeper insight into the mode of action behind individual compounds, as this could facilitate the exploitation of, e.g., synergistic combinations with more powerful antimicrobial properties (Hyldgaard, 2012).

The antifungal activity of a given essential oil may be isolated effects or synergic action by two or more compounds (Cruz et al., 2015). However, increasing amounts of evidence indicate that the inherent activity of essential oils may not rely exclusively on the ratio in which the main active constituents are present, but also interactions between these and minor constituents in the oils. Various synergistic antimicrobial activities have been reported for constituents or fractions of essential oils when tested in binary or ternary combinations (García-García et al., 2011). However, if we can use this technology at this moment, there is need to think about environment and economic sustainability questions under agroecology system prerogatives. In this case, the direct use of the essential oil on the cropping process to control insects and pathogenic microorganisms is possible. To the resources, free input given by farmers is possible in planting economic or alimentary plants with phitossanitary plants for essential oil production or other environmental services.
Effect of essential oils at different doses on A. solani mycelial growth in vitro (Experiment 1)
A. solani mycelial growth was differently influenced, when the studied essential oils and their concentrations was compared (
Table 1 and Figure 1).
Action of the mycelial growth control can be possible because it has a high variation on the essential oils compounds. There are two hypotheses for the complex action: First, synergic action among the compounds against A. solani; second, horizontal action with multiples attack points in the membranes and biochemical process in the mycelial tissue.
In addition, Daferera et al. (2003) reported that R. officinalis oil was inefficient against B. cinerea and F. solani mycelial growth, corroborating the present results.
As such, essential oils did not show any action against A. solani under the conditions of the present work. They were eliminated at this stage. The opposite was observed for E. citriodora, M. alternifolia, M. piperita, S. aromaticum, C. citratus, C. martini, and C. zeylanicum oils, which not only reduced the colony growth rate but also completely inhibited it at the highest doses (Table 1).
For the remaining oils, the fungistatic effect for five days on colony stagnation was confirmed, and mycelial growth was completely inhibited at doses higher than 750 and 1000 μL L–1 (Figure 1).
Similar to the results obtained in the present work, several authors have reported the fungicide and fungistatic effects of different doses of S. aromaticum, C. zeylanicum, C. citratus, C. martini, E. citriodora, M. alternifolia and M. piperita essential oils against in vitro pathogens development: Aspergillus parasiticus, Aspergillus niger, Aspergillus flavus, Aspergillus fumigatus, Rhizopus sp, Penicillium sp, Europium repens, Rhizoctonia solani, Rhizoctonia bataticola, Fusarium moniliforme, F. oxysporum, F. solani, Bipolaris sorokiniana, Botrytis cinerea, Helminthosporium oryzae, Alternaria sp., Alternaria porri, Myrothecium verrucaria, and Curvalaria lunata (Pawar and Thaker, 2007; Negrelle and Gomes, 2007; Faria et al., 2006; Fonseca et al., 2006).
Besides, Faria et al. (2006) reported Alternaria sp. was susceptible to the essential oil of Ocimum gratissimum chemotype-eugenol as well as to isolated eugenol. Pawar and Thaker (2007) also observed that essential oils of other plant species rich in eugenol were efficient against Alternaria porri. Salgado et al. (2003) studied the bioactivity of several Eucalyptus sp. essential oils; E. urophylla oil was the most efficient; E. citriodora oil was against F. oxysporum Schlecht., Botrytis sorokiniana Shoemaker, and Botrytis cinerea Pers, with dose-dependent inhibition point.
The antifungal effect observed for S. aromaticum and C. zeylanicum is probably due to eugenol, the main compound in both species: 82.55 and 73.45%, respectively, of their composition. In addition, there are reports of the antifungal effect of isolated eugenol against A. solani (Faria et al., 2006), Fusarium spp., Rhizopus spp., and others (Souza et al., 2005).
Campaniello et al. (2010) found that eugenol (from 100 to 150 μg mL−1) is an effective antifungal compound against Aspergillus, Penicillium, Emericella and Fusarium spp., suggesting that this activity could be attributed, in part, to the presence of a phenolic group.
C. citratus oil antifungal effect may be related to its major compounds, neral (34.34%) and geranial (46.91%), or to their associate action with the remaining oil components. This hypothesis is confirmed by Negrelle and Gomes (2007), who reported that the mixture of these aldehydes constitutes citral, to which important antifungal activity is attributed. Souza et al. (2005) also detected potent antifungal activity by citral against some phytopathogens like Fusarium spp. and Rhizopus spp. The activity of E. citriodora oil was probably due to its main compound, citronellal (74%). This hypothesis supports the results obtained by Salamci et al. (2007), who stated that the antifungal activity of essential oils is generally attributed to their major compound. It must be considered, however, that in certain cases the antifungal action, as well as its intensity, may be a result of the synergistic effect among the oil constituents. For example, neral and/or geranial activities are potentiated when they are associated with myrcene (Onawunmi et al., 1984).
Effect of essential oils at different doses on A. solani conidium germination in vitro (Experiment 2)
The effects observed for conidium germination was similar to those observed for A. solani mycelial growth (Table 2).
Corroborating the present results, Caccioni and Guizzardi (1994) also detected an inhibitory effect by the same concentrations of C. zeylanicum and Cymbopogon sp essential oils on Monilinia laxa, Mucor piriformis and Rhizopus stolonifer mycelial growth and spore germination. S. aromaticum, C. citratus, C. martini, and C. zeylanicum essential oils similarly inhibited conidium germination at 750 μL L –1, whereas inhibition by E. citriodora oil occurred only at 5000 μL L-1 (Table 2).
Mishra and Dubey (1994) observed conidia germination and mycelial growth inhibition in F. moniliforme, A. flavus and A. fumigatus and other 47 species under C. citratus essential oil. On the other hand, these authors explain the importance of the cropping seasons of C. citratus on the essential oil compounds and their effect on fungi control.
Effect of essential oils at different doses on the control of early blight caused by A. solani in the leaves of S. lycopersicum (L. esculentum) Mill. cv. Sta. Clara cultivated under greenhouse conditions (Experiment 3)
Supporting the results obtained in the in vitro experiments for mycelial growth (Table 1 and Figure 1) and conidium germination (Table 2), in vivo studies indicated a significant reduction in early blight severity in the leaves of S. lycopersicun cv. Sta. Clara cultivated under field greenhouse conditions (Figures 2 and 3). However, we can identify the significant dose effect difference by the MIC 50, 90% and Ymax estimated by biological and mathematical model to reduce symptoms by each oil (Table 3).
C. zeylanicum essential oil reached its maximum response (78, 2%) lower than the remaining oils, indicating that even a ten-fold increase in its dose is inefficient (Figure 2, Table 3).
Only E. citriodora essential oil at 4.802,64 µL L-1 and C. citratus oil at 3.172,80 µL L-1 was 100% efficient in controlling early blight severity, relative to the remaining species. S. aromaticum and C. martini oils at 2.998,39 µL L-1 had 92% and 2898.642 µL L-1 90% control, respectively (Figures 2 and 3; Table 3).
The chemical compounds observed at this treatment was geranial (46.91%), neral (34.34%), geranyl acetate (6.30%), camphene (1.02%), 6-methyl-5-hepten-2-one (1.28%), linalool (0.82%), citronellal (0.25%), isomenthol (1.81%), nerol (0.45%), geraniol (3.52%), and α-
trans-bergamotene (0.85%) (
Figure 4).
Nashwa et al. (2012) found that different concentrations of six plant extracts, O. basilicum, A. indica, E. chamadulonsis, D. stramonium, N. oleander, and A. sativum, significantly reduced the early blight disease. The best results for MIC 50 and 90 with minor dose necessary to control was essential oil from C. citratus at 341, 32 and 1.822,10 µL L-1 respectively (Table 3).
These results are corroborated by GlamoÄlija et al. (2011) and Geromini et al. (2015) on the effectiveness of Geranial and Neral aldehyde mixture that constitutes citral, a potent bioactive substance that fights against phytopathogens, especially fungi and post-harvest and storage diseases.
Farming under agroecological system is possible with the use of these essential oils. If the farmer has this species in their land, they can proceed to the oil extraction by using steam distillation, and MIC 90 with a spreader-sticker. However, two applications are needed per week. Moreover, it is now also possible to develop a new product based on natural compound with more studies around formulation and chemical stability on field.