Full Length Research Paper

ABSTRACT
In recent years, essential oils (EOs) as alternatives to synthetic pesticides in managing pests have been assessed. The use of bioinsecticide in pest management is encouraged in agroecology for a sustainable agricultural system. Essential oils of Lantana camara L. leaves from different climatic zones of Kenya were extracted by steam distillation and analyzed through GC-MS to identify the compounds. The contact toxicity and repellent activity of EOs against the invasive tomato pest, Tuta absoluta, were tested. The toxicological assays were performed following the leaf-dip bioassay protocol, while the repellency activity was performed using the repellent response method for phytophagous pests and the data analyzed using the ANOVA test. It was found that L. camara EO has a good insecticidal activity with higher mortality (89%) on the 2nd instar larvae with a higher dosage (0.01 µl/µl). The repellence test also showed a higher average repellence (93.44%) effect with a higher dosage (0.01 µl/µl) of the EOs. According to these results, the EO of L. camara may well be a sustainable, eco-friendly alternative for synthetic insecticide in the T. absoluta management program.
Key words: Bioinsecticide; essential oil, integrated pest management, invasive species, secondary metabolites.
INTRODUCTION
The leaf miner, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae), is a pest of vital global economic importance. It limits tomato production (Lycopersicum esculentum L.) worldwide (Campos et al., 2017; Desneux et al., 2011; Guedes et al., 2019). As a result, this pest has gained notoriety as a pest species that can cause destruction and losses of up to 100% when there is no intervention (Rwomushana et al., 2019).
Current management of T. absoluta typically relies solely on synthetic insecticides (Silva et al., 2019). However, this management strategy has not provided a total solution to the problem due to insecticide resistance and pest resurgence (Guedes et al., 2019; Roditakis et al., 2018). Furthermore, these products are already proving to be harmful to the environment (Damalas and Koutroubas, 2018; Gill and Harsh, 2014), causing the development and progression of several health issues in humans (Özkara et al., 2016). To cut back the excessive use of synthetic insecticides, exploring environmentally friendly and sustainable alternative strategies, such as natural products from wild plants, remains a viable option. Natural products directly utilized as pest control agents offer a more sustainable pest management solution than synthetic pesticides (Nuruzzaman et al., 2019).
Essential oils (EOs) from pesticidal plants used as pesticides play a crucial role in controlling pests sustainably (Campolo et al., 2017)by reducing the pest population while minimizing the environmental effect. Lantana camara is a wild plant that produces Eos, extensively studied for its bioactive properties and reported as having insecticidal (Javier et al., 2017; Murugesan et al., 2012, 2016), acaricidal (Adehan et al., 2016), larvicidal (Costa et al., 2010; Zandi-Sohani et al., 2012), fumigant (Gotyal et al., 2016), acetylcholine inhibition, repellent (Yuan and Hu, 2012), and antifeedant (Chau et al., 2019; Yuan and Hu, 2012)action among other features against a wide range of pests. These bioactive properties exhibited by L. camara EO make it a novel candidate for use as a pesticide with multiple actions. For example, Murugesan et al. (2016)tested the L. camara EO on Hyblaea puera and Ahevidae fabriciella (Lepidoptera) at a concentration of 10000 ppm, reported a 62% larval mortality and concluded that the EO expressed insecticidal and antifeedant properties. Besides, Javier et al. (2017) tested the EO of L. camara for bioactivity against Spodoptera litura (Lepidoptera), showing remarkable insect growth regulatory activities and direct toxicity. This corroborates Deshmukhe et al.(2011)that it has the potential to be exploited as a botanical insecticide for cutworm management. In their study, Costa et al. (2010) tested for larvicidal activity against A. aegypti larvae using the EOs from the leaves of L. camara and showed that it has larvicidal potential. However, all these studies show the potential this plant has in the management of pests. There is no scientific investigation of its EO bioactivity on T. absoluta.
MATERIALS AND METHODS
Plant materials
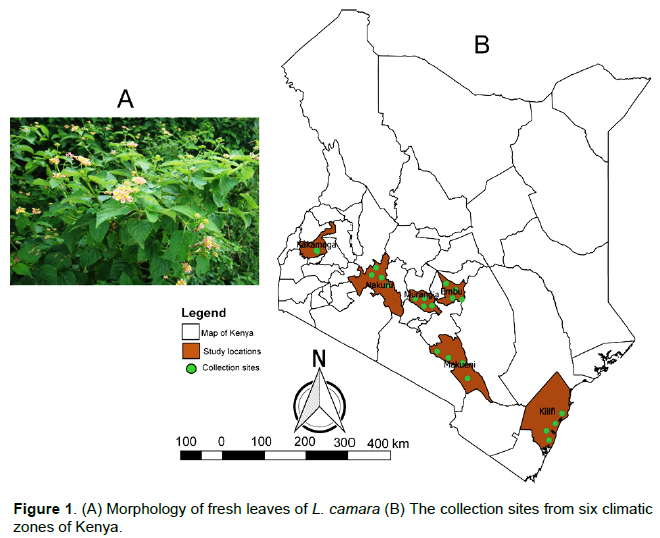
Fresh, healthy first four leaves of the stem from the top of L. camara plants (Figure 1A) were collected from six different climatic zones of Kenya, namely; Lower Highland-Nakuru (LH-NJ), Upper Midland 1-Kakamega (UM1-KK), Upper Midland 2-Kandara, Muranga (UM2-KA), Upper Midland 3-Embu (UM3-EM), Lower Midland-Kiboko, Makueni (LM-KI) and Coastal Lowland-Mtwapa, Kilifi (CL-MT) in July 2018. The exact location of the collection sites are marked with dots in Figure 1B. The leaves of L. camara from each sampling site were harvested from the first four leaves of the stem from the top of plants and mixed to make a composite sample. The samples were transferred to the lab within two days of collection in ventilated nylon gunny bags. The samples were washed and then air-dried immediately under room temperature (23–26°C) in a well-ventilated room for two weeks until crisp. Subsequently, they were grounded into powder and stored in khaki bags until it was time for oil extraction.
Extraction of the essential oils
The L. camara EO was isolated by steam distillation using a steam distiller apparatus (Deschem Science supply, China). First, 200 g of dried leaves from each sample was steam distilled separately using 2000 mL distilled water for 3 h. Next, the condensing oils were separated with a separating funnel, and the oily sample was treated with anhydrous sodium sulphate (Na2SO4) to remove the remaining trace of water and collected in amber-coloured vials, labelled, and stored at 4°C until use. The experiment was performed in triplicates for each sample, and the yield was averaged over triplicates. The percentage yields of EOs were determined based on dry matter and calculated as the weight of the extracted oil (g) /200 g of the dry weight of the leaf sample (% w/w).
Gas chromatography-mass spectrometry (GC-MS) analysis
The EO samples were diluted in n-hexane and transferred to the auto-sampler vials for GC-MS analysis. GC-MS-QP2010 SE (Shimadzu Corporation) coupled with an autosampler was used for this study. Ultrapure He was used as the carrier gas at a flow rate of 1 ml/min. A BPX5 non-polar column, 30 m; 0.25 mm ID; 0.25 µm film thickness, was used for separation. The GC was programmed as follows: 60°C (1 min); 10°C/min to 250°C (25 min). The total run-time was 45 min. Only 1 µL of the sample was injected. The injection was done at 200°C in split mode, with a divided ratio set to 10:1 and the interface temperature set at 250°C. The Electron Ionization (EI) ion source was set at 200°C. Simultaneously, the mass analysis was done in full scan mode, in a range of 40 – 550 amu (Shimadzu GC-MS QP2010 SE solution software (Tokyo, Japan)). The raw mass spectra obtained matched against the NIST 2017 Library of Mass Spectra for possible identification of compounds. Samples were analyzed at the Jomo Kenyatta University of Agriculture and Technology Analytical Chemistry Laboratory.
Rearing of test insects and plants
T. absoluta insects for the experiment originated from infested tomato leaves collected at the International Centre of Insect Physiology and Ecology (ICIPE) (S01°13.140’; E036°53.440’) Insectary Laboratory, Nairobi, Kenya. The adults were transferred to the laboratory within 2 h. Upon arrival in the laboratory, they were released in the insect-proof rearing cages (50×60×80 cm) and provided with four insect-free four potted tomato plants (height: 25 cm) and reared to the first filial (F1) generation as described in Roditakis et al. (2013). Newly-emerged adults of T. absoluta (F1 generation) were released inside another net cage to obtain homogeneous T. absoluta larvae (same age, nutritional and general health) (IRAC method No. 022). The adults of T. absoluta were provided with water and an energy source (commercial honey 1:1 dilution). The insects were allowed to oviposit for 24–48 h. The oviposition level was evaluated visually. If an adequate number of eggs were observed (that is, more than 150–200 eggs), then the plant material was carefully removed, and new plants were placed in the oviposition area to allow the continuation of the oviposition (IRAC method No. 022). The plant material infested with T. absoluta eggs were placed in an insect?proof rearing cage to allow larval development to the second instar.
Tomato (Rio Grande VF) plants (L. esculentum) used for rearing T. absoluta and for performing the bioassays experiments were grown in the greenhouse conditions in 2-L pots, inside large insect?proof cages and maintained pest-free under (30±3°C) temperature, (75-80%) relative humidity and (12:12) light: dark conditions. The plants were screened for the presence of pests every second day; in the rare event of detected infestations, these were manually removed by cutting the infested leaves or removing and destroying the plant (Roditakis et al., 2013). Therefore, no insecticides were used during the plant development phase.
Bioassay experiments
The experiments were conducted at the Department of Horticulture and Food Security at Jomo Kenyatta University of Agriculture and Technology (Kenya) under controlled environmental conditions in growth chambers, maintaining a temperature of 25 ±2°C, 65-70% relative humidity, and 12:12 light: dark photoperiod regime.
Contact toxicity
The second instar larvae were collected from the rearing cages, and accordingly, the toxicological assays were performed following the leaf-dip bioassay protocol of the 199 Insecticide Resistance Action Committee (IRAC) test method 022 (www.irac-online.org), with minor modifications. The EOs extracted from plants from the six climatic zones were prepared in three different dosages (0.01, 0.001, and 0.0001 µl/µl) mixed with 0.1% Tween® 20 (Sigma-Aldrich, Germany) and used as treatments. A commercial formulation of Flubendiamide (BELT® 480 SC, Bayer AG, Germany) insecticide was used as a positive control, whereas sterile distilled water containing 0.1% Tween® 20 (Sigma-Aldrich, Germany) as a non?ionic wetting agent was used as a negative control with three replications. The insecticide was used as per the manufacturer’s recommendations (0.1 µl/µl).
Sufficient non-infested, untreated tender young whole tomato leaflets of uniform size were collected and kept in sealed plastic bags to prevent them from wilting. Complete tomato leaflets were dipped for 5 s in the EO concentrations with gentle agitation to ensure the entire surface is covered equally. The treated leaflets were dried on a wire net with an upper leaf surface (abaxial surface) facing skywards and placed in a labelled petri dish (Ø- 90 mm) with slightly moistened filter paper covering the bottom. Around 0.2 ml distilled water was used; which was sufficient to wet the filter paper keeping the leaf material turgid throughout the bioassay period. Second?instar T. absoluta larvae were carefully removed from the galleries in infested tomato leaves under a light?bed (transparent bench with fluorescent illumination underneath). In each petri dish, ten larvae (2nd instar) (4-5 mm) were released carefully using a subtle soft brush to avoid damaging the very fragile larvae. Subsequently, all the Petri dishes sealed using a ventilated muslin cloth.
Larval mortality was assessed after 24, 48, 72, and 96 h of exposure whereas death was recorded under a magnifying glass (Osho® 10× magnification, Kenya). The larvae were scored as dead if they could not make coordinated movement from a gentle stimulus with a fine brush to the posterior body segment. The experiment was repeated three times simultaneously, and the average mortality obtained.
Repellent activity
In testing the repellent activity of the EOs, Whatman No.1 filter paper was cut to fit the size of the plastic dish and divided into two equal parts. Half of the filter papers was treated with 0.1% Tween® 20 as control and the other half with EO concentrations mixed with 0.1% Tween® 20 of 0.01, 0.001, and 0.0001 μl/μl. Twenty larvas (2nd instar) were placed in the middle of each Petri dish. The plastic dishes were closed and tightened with Parafilm. The experiment was carried out in five replications and at the same environmental conditions with insect rearing. After four hours of exposure, the number of insects in each half of the filter paper was recorded and the percentage repellency (PR) calculated using the following formula:
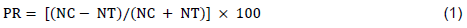
PR = percentage repellency, NC = number of larvas in the control area and NT = number of larvas in the treatment area.The mean repellency value of each extract was calculated and assigned to repellency classes from 0 to V: class 0 (PR ≤ 0.1%), class I (PR = 0.1-20%), class II (PR = 20.1-40%), class III (40.1-60%), class IV (60.1-80%), and class V (80.1-100%).
Statistical analysis
The mean number of live larvae per leaf was tested for per cent mortality. The per cent mortalities were corrected for control (that is, natural) mortality using Abbott’s formula (Abbott, 1925). The two-way analysis was conducted with the EOs from different climatic zones as the main effect, concentration rate as the covariate, and larvae mortality registered at different time intervals (24, 48, 72, and 96 h) as the response variable. In addition, the toxicity effect of the different EOs concentration on the second instar larvae was compared using the analysis of variance (two-way ANOVA) and the means compared by LSD test at 5% level (SAS®, On-Demand for Academics).
Concentration–mortality data (obtained from the bioassay) was computed using Probit procedure (PROC PROBIT LOG10; SAS®, On-Demand for Academics.) to estimate the median lethal concentrations (LC50 values), their 95% fiducial limits (FL) along with their respective standard errors, as well as slopes of the curves. The LC50 values were significantly different when their 95% fiducial limits did not show similarity.
Per cent of larval repellency was analyzed using ANOVA (Analysis of variance) (SAS®, On-Demand for Academics). The negative values were treated as zero. The larval repellency was calculated using the simplified contact repellency test. The mean percentage of larvas that repelled into the untreated side was corrected by a control test. The square root of the per cent of repellency in each test was arcsin converted, and ANOVA and the multiple comparisons of the repellency by LSD was performed using SAS®, On-Demand for Academics.
RESULTS
Chemical composition and content of Lantana camara L. essential oils
The GC-MS analysis of L. camara EO from the same species growing in diverse climatic zones of Kenya resulted in 123 compounds with a higher content than 0.01%, of the oils. Sesquiterpenes followed by monoterpenes dominated the significant components identified (over 1.0% in content) (Table 1) listed in order of their elution on the HP-5 MS column. The sesquiterpenes were dominated by the β-caryophyllene (5.11 - 14.31%) and spathulenol (4.22 - 9.50%) as sesquiterpene hydrocarbon and oxygenated monoterpene, respectively. On the other hand, the monoterpenes were dominated by Sabinene (2.84 - 12.54%) and eucalyptol (1.44 - 8.81%) as monoterpene hydrocarbon and oxygenated monoterpene, respectively.
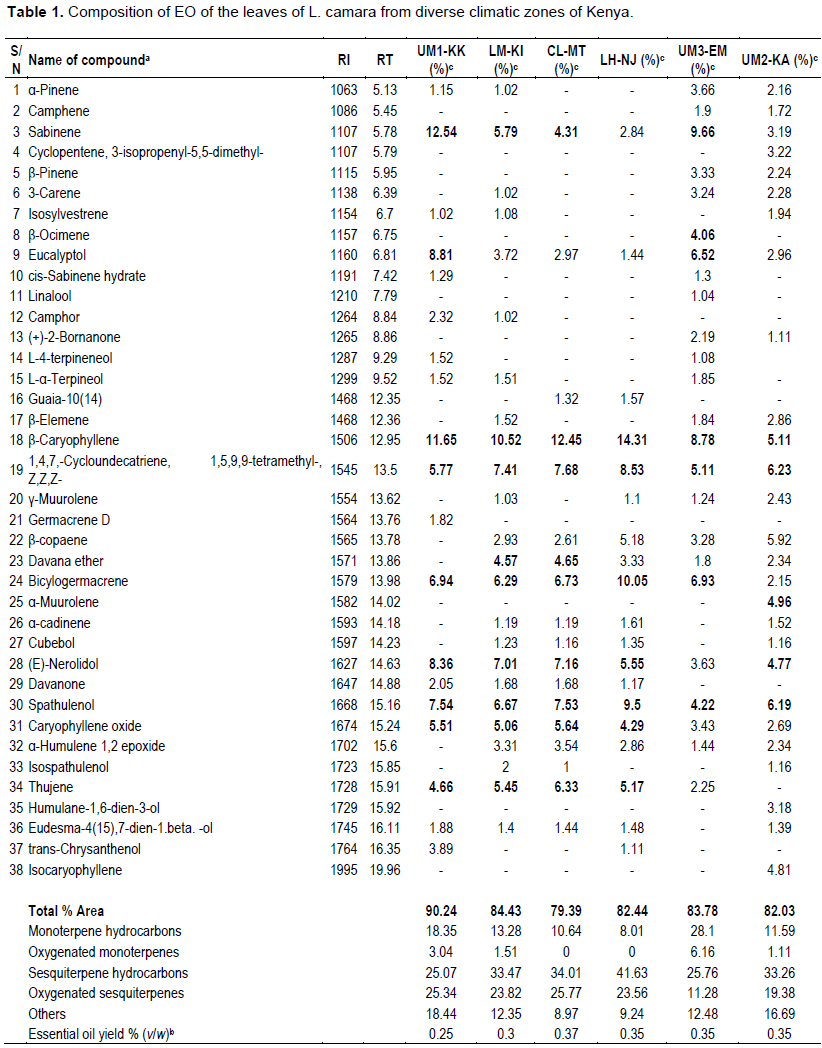

The yield of the essential oil of L. camara collected at different regions was obtained from dried leaves that ranged from 0.25 to 0.37% w/w. The highest yield of oil was observed in samples collected from Mtwapa (CL-MT) (0.37%), characterized by high mean temperatures (26°C) and average precipitation of 125 mm. The opposite result was observed for the oil obtained from samples collected in Kakamega, which presented a lower yield of 0.25%. During this period, there was lower maximum (24.5°C) and minimum (15°C) temperatures, in addition to higher volume of total precipitation (283 mm). In terms of the compounds, the samples from Kakamega exhibited higher monoterpenes levels, with Sabinene being the predominant compound, while the samples from Mtwapa exhibited higher levels of Sesquiterpenes with β-caryophyllene being the predominant compound.
Toxicological bioassays
The toxicological two-way analysis of variance suggests that both the dose concentration rate (F = 13.11; df = 2; p < 0.0001) and the EO formulations (F = 6.44; df = 7; p < 0.0001), registered at 24 h, 48 h, 72 h and 96 h after the treatment influenced the larva mortality. The statistical analysis revealed a significant interaction between dose concentration, EO formulations and observation time (p < 0.0001). However, there was no interaction among the two variables, the dose concentration and EOs formulations (F = 0.01; df = 10; p 1.00).
Contact toxicity on larvae
The mean average larval mortality in the positive control-insecticide -Flubendiamide) (PC-INS) was 16, 30, 76 and 93% after 24, 48, 72 and 96 h of exposure, respectively. Whereas in the negative control-water (NC-WA) treatment, no larvae died during the 96 h of the experiment. The positive control (PC-INS) showed the highest larval mortality compared to the EOs formulations (Figure 2).
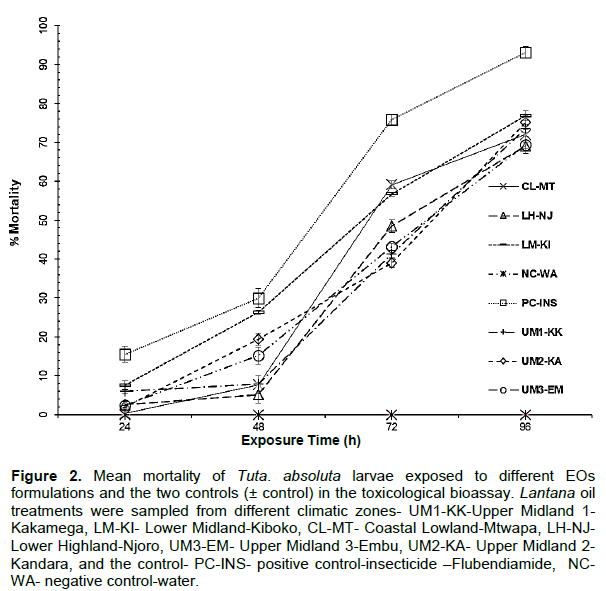
In the first sampling (24 h after the treatment), LM-KI and UM1-KK EO formulations were the most effective in killing the larvae’s 8 and 6% mortality, respectively. Overall, the positive control (PC-INS), on average, was most effective than the EO formulations (Figure 2). In the second sampling (48 h), the mortality of T. absoluta larvae increased significantly in all the treatments compared to the first sampling. In the third, and fourth sampling, the mortality rate increased for all the treatments, each attaining above 40% larval mortality except UM-KA, treatment which was 39%. In the fourth sampling, all the treatments reached a mortality rate above 60%.
Throughout the trial, the LM-KI EO formulation was the most effective against the larvae (maximum average mortality = 77%), whereas the LH-NJ and UM3-EM EO formulation could only kill a maximum of 69% of the exposed larvae. Overall, there was no significant difference in the mortality induction by the different EO formulations.
The dose concentration profoundly influenced the larval mortality rate. In all the time point, the positive control (0.1 µl/µl-insecticide recommended dosage), on average, was most effective in killing the larvae’s (average mortality= 93%) than the EO formulations dosage concentrations of 0.01, 0.001, and 0.0001 µl/µl (average mortality= 89, 71 and 58% respectively) (Figure 3). The longer the larvae’s exposure to the treatment, the more the mortality rate increased in all the dose concentration. The highest dosage of the EO formulations (0.01 µl/µl) showed the highest mortality at all the time point of sampling, which was reasonably comparable to the positive control (0.1 µl/µl).
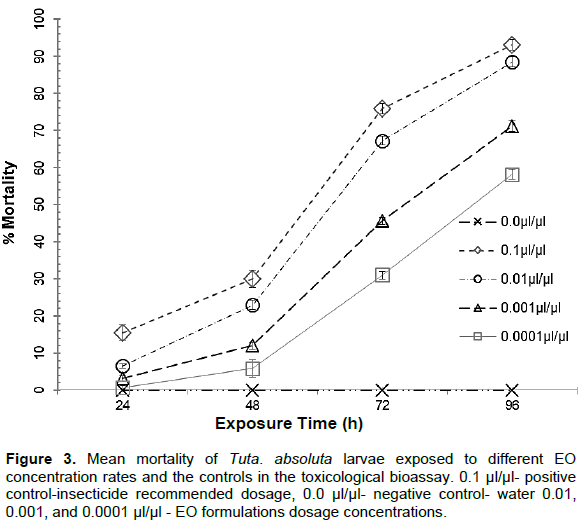
The EO concentrations of 0.1 µl/µl were the most effective in killing the larvae at 16% mortality, followed by 0.01 µl/µl at 7% in the first sampling (24 h after the treatment). The mortality rate of T. absoluta larvae increased in all the treatments in the second sampling (48 h), and in the 0.01 µl/µl dose concentration, the mortality rates increased three times, compared to the first sampling. None of the dosage concentration, including the positive control, attained 50% larval mortality within the early 48 h of larval exposure. However, by 72 h, the positive and 0.01 µl/µl dosage concentration had achieved above 50% mortality.
Median lethal concentrations (LC50) for the larvae
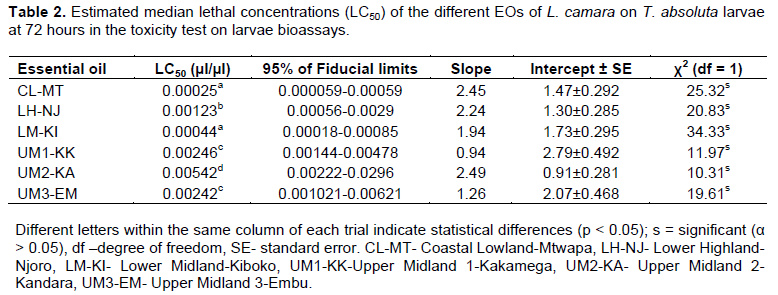
The LC50 values (Table 2) was calculated for 72 h because the maximum mortality registered at 24 and 48 h were less than 50%, while at 96 h, it was over 50%. Therefore, this was the best time since the longer the exposure time, the lower the LC50 values. The dose-response mortality data exposed to the EO formulations presented χ2 values < 34.33. This parameter shows the appropriateness of the model to estimate the LC50. Evaluation of toxicity of the EOs, based on LC50 values and fiducial limits, showed that significant differences among- EO toxicity for CL-MT, LH-NJ, LM-KI, UM1-KK, UM2-KA and UM3-EM were 0.00025, 0.00123, 0.00044, 0.00246, 0.00542 and 0.00242 μl/μl, respectively (Table 2). The EO extracted from CL-MT climatic zone showed the highest capacity to kill the exposed larvae (LC50 = 0.00025 µl/µl). The EO from the UM2-KA formulation required the highest concentration (LC50 = 0.00542 µl/µl) to kill 50% of the exposed larvae (Table 2). The mortality rate of the larvae depended on the concentration of EO and increased with increasing oil dose concentrations. For all the six oil formulations, the highest average mortality (93.44%) was observed at the highest dose concentration (0.01µl/µl) (Figure 3).
Repellent activity
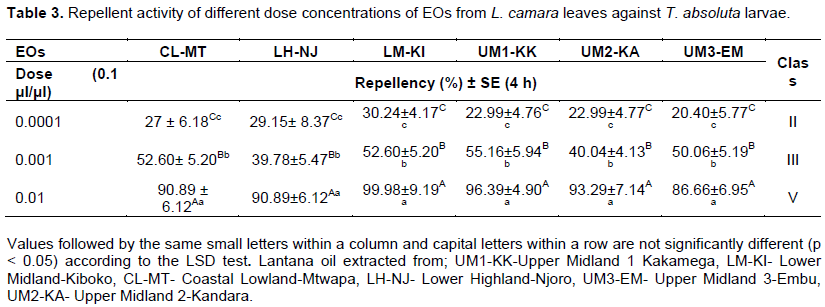
Results given in Table 3 describes the repellent activity of different EO formulations and concentrations of L. camara leaf EO. The results revealed that there was no significant difference between the EO formulations (F = 1.68; df = 5; p = 0.1505) and the interaction between the variables, dosage concentration and EO formulations interaction (F = 0.77; df = 10; p = 0.6539) after 4 h of exposure. The EO of L. camara formulations showed significant repellent activity against the second instar larvae of T. absoluta at all concentrations (F = 156.28; df = 2; p < 0.0001) within the 4 h of exposure. The repellency activity of L. camara EO was dose-dependent. The repellency increased when dosage levels increased. Complete repellency was observed when the highest dose level (0.01 μl/μl) was applied although there was a slight variation in the repellent effects within EO formulations. LM-KI EO showed the most repellency activity with 99.97% of repellency induced by 0.01μl/μl whereas the treatment of 0.0001 μl /μl induced only 30.24% of repellency of T. absoluta larvae, with an average rate of 60.93%. UM3-EM EO showed the lowest repellency activity within 4 h with 86.66% repellency induced by 0.01 μl/μl whereas the dose of 0.0001 μl/μl induced only 20.40% of repellency of T. absoluta larvae, with an average rate of 55.70%. Based on the mean repellency rate, EO formulations of all the L. camara leaves from diverse climatic zones showed repellency classes II, III and V with increasing dosage.
DISCUSSION
Essential oils from plants produce a significant amount of valuable bioactive compounds. These compounds are mixtures of monoterpenes, sesquiterpenes, phenols, aldehydes, alcohols or other compounds (Olayemi, 2017). Compound accumulation and yield variations in EO composition are rather typical even within the same species. It depends on the genotype, plant organ, harvest, geographical region, season, plant nutritional status, climatic conditions-temperature, humidity, and light intensity (Moustafa et al., 2016; Ncube et al., 2012; Pereira et al., 2019; Swamy et al., 2017). The oils of L. camara showed considerable variability in the chemical composition, percent constituent and oil yield from the same species growing in diverse ecological conditions of Kenya. Our results corroborate Bendera (2007)and Syombua (2015), who also found variability in the chemical profile of L. camara EOs harvested from Maseno-western Kenya and the eastern part (Kitui and Machakos), respectively. Similarly, reports from the different parts of the world also show remarkable differences in the chemical composition (El et al., 2014; Khalid, 2019; Moustafa et al., 2016; Murugesan et al., 2016; Nea et al., 2017; Omoregie et al., 2016; Swagatika and Smaranika, 2017).
This study established that the EO of the L. camara wild population in Kenya is rich in sesquiterpene and monoterpene compounds. They are occupied mainly by hydrocarbons and oxygenated sesquiterpene compounds with an average of 36.68 - 22.73% and 22.49 - 11.28%, respectively. The sesquiterpenes were discriminated by the dominant presence of β-caryophyllene, which agrees with previous studies, such as Dos et al. (2019). They found β-caryophyllene content at 13 - 8.9% in Brazil, while Khalid (2019)and Dougnon and Ito (2019)found it as 17.9 and 16.7 - 8.9% in Egypt and Benin, respectively. On the other hand, Sabinene (12.54 - 3.19%) dominated the monoterpenes, corroborating the findings of Nea et al. (2017)with up to 9.0% relative abundance. These results are consistent with our study; however, there are some differences in the percentages of detected compounds related to genetic and environmental factors (climate, seasons, geography and geology) variability. Several versatile standard components were present in all the EO analyzed in this work, including; bicylogermacrene, spathulenol, eucalyptol, (E)-nerolidol, and caryophyllene oxide, which are beneficial bioactive compounds. Based on the above fact, L. camara, cultivation widely in Kenya, stands a chance as a source for the isolation of various natural compounds and their bioactive properties tested for pest management.
The oil yields from L. camara leaves obtained in different regions significantly varied (Table 1). The populations around Mtwapa had the highest yield, while the lowest yield was observed in Kakamega. The populations of Mtwapa grow on the coastal lowland characterized by high temperature and a reasonable amount of rains. The environments there are sandy soils and have vast day/night temperature difference. These factors may have resulted in high essential oil contents. However, in Kakamega, the prevailing conditions were dominated by low average temperatures and higher precipitation, providing a conducive growth environment. The differences in the chemical composition and yield of essential oils can be related to climatic and geographic factors such as temperature, ultraviolet radiation, atmospheric pollution, altitude, water and nutrient availability, and the developmental stage of the plant and genetic factors.
This paper shows the bioactivity studies of L. camara EO as a potential bioinsecticide to manage the tomato leaf miner, T. absoluta. While previous reports showed the potential of L. camara oil to be effective in the control of lepidopteran pests (Chau et al., 2019; Javier et al., 2017; Kasmara et al., 2018; Murugesan et al., 2016), our study provides the observed bioactivity of the EOs of L. camara from various climatic zones against the second instar larvae of the leaf miner, T. absoluta, together with their repellence activity; thus contributing to a thorough understanding of the potential use of EOs in the management of agricultural pests.
Notwithstanding, L. camara EOs have undergone bioassay tests as insecticides on several pests, but only a few studies have focused on the tomato borer. Furthermore, most bioassays reported focuses on trials lasting for less than 48 h, whereas the lasting effects of these compounds are lacking. To the best of our knowledge, no study on L. camara EO bioactivity against T. absoluta, together with their toxicity differences from oils sourced from the same plant but differing climatic zones, and their repellency activity has been recorded. However, previously, several researchers have tested the EOs from other plants against T. absoluta larvae for toxicity (Abdel-Baky and Al-Soqeer, 2017; Allam Tarik, 2015; Chegini et al., 2018; Javier et al., 2017; Khaoula, 2013; Moreno et al., 2012; Soares et al., 2019; Yarou et al., 2018)and their repellence activity (Allam Tarik, 2015). The EOs showed considerable toxicity and repellency activity on the T. absoluta larvae, comparable to the other plants EOs.
In our study, the insecticidal activity varied with the dosage concentration of the EO formulations. In the toxicity trial, the larval mortality was dose-dependent, and larval susceptibility increased with increasing dosage. Meanwhile, among the L. camara EO formulations, the larval mortality recorded did not significantly differ but differed substantially from the positive control (insecticide). Thus, the exposure concentration of the EO is proportional to the level of toxin and toxicity. Besides, the longer the exposure time, the higher the mortality rate recorded since longer time increases toxin accumulation. As shown by the LC50 value of L. camara, EO formulations at 72 h of exposure showed a more substantial toxicity level.
The present study shows that leaf application of L. camara EO is equally practical as a contact toxicant against the leaf miner, T. absoluta. Exposure of larvae to botanical compounds occurs through contact or systemic actions (Rwomushana et al., 2019). In particular, exposure to EO affected the larvae by weakening their development (Javier et al., 2017)and increased mortality. Larval mortality increased to over 80%, at the maximum dose concentration, showing that the EO of L. camara has larvicidal properties. For instance, in an experiment involving T. absoluta larvae, the neem extract at different concentrations resulted in 86.7 to 100% larval mortality (Elshiekh et al., 2014).
Besides, our study suggests that the T. absoluta second instar larval stage is relatively a critical stage to target the leaf miners. Eggs are less susceptible to insecticides (Elshiekh et al., 2014; Javier et al., 2017)because of the high numbers of enzymes that break down insecticides (Campbell et al., 2016). Our results were in line with those reported by Javier et al. (2017)and Kasmara et al. (2018), who found that L. camara EO caused over 50% larval mortality of Spodopteralitura (Lepidoptera). Other plants (eucalyptus, basil, castor bean, garlic, chinaberry, geranium, thyme, and onion) tested against T. absoluta larvae show insecticidal activity with different efficacies (Abd El-Ghany et al., 2016). The results also show that the L. camara EO has an outstanding repellency action on the second instar larvae of leaf miner, T. absoluta.
The volatile constituents such as β-caryophyllene, sabinene, bicylogermacrene, spathulenol, eucalyptol, (E)-nerolidol, thujene, caryophyllene oxide, β-copaene, davana ether, trans-chrysanthenol, α-humulene epoxide, linalool, limonene, terpineol, terpene-4-ol, α-pinene and camphor in the L. camara EO could be responsible for their observed insecticidal and repellent activity. Zandi-Sohani et al. (2012)also concluded that it might involve these components in the repellent and insecticidal activities against C. maculatus. A trial against T. absoluta larval stages using pure α?pinene moderated toxicity effects was reported (Chegini et al., 2018). Limonene, 3?carene, terpinolene, β?myrcene and γ?terpinene have larvicidal activities to Aedes aegypti and Aedes albopictus larvae (Cheng et al., 2009). However, the role of a single compound in the EO is not definite, but the minor constituents act as synergists, enhancing the effectiveness of the EO (Akhtar and Isman, 2013; Hanem, 2012).
The EO formulations of L. camara were effective in controlling the target pest. Therefore, this could be a source of new bioinsecticide against T. absoluta. Further works may be necessary to assess their efficacy under realistic field conditions. However, there is a need to increase efficiency by developing methods that will allow for long-lasting effectiveness. Besides, there is no information on the potentially lethal and phytotoxic effects of L. camara EO on non-target organisms and target crop. Therefore, this is a new avenue for future research and serves as a source of hypotheses for further research on L. camara essential oil as a potential bioinsecticide.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Abd El-Ghany NM, Abdel-Razek AS, Ebadah IM A, Mahmoud YA (2016). Evaluation of some microbial agents, natural and chemical compounds for controlling tomato leaf miner, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Journal of Plant Protection Research 56(4):372-379. |
|
|
Abdel-Baky NF, Al-Soqeer A.A (2017). Controlling the 2nd instar larvae of Tuta absoluta meyrick (Lepidoptera: Gelechiidae) by simmondsin extracted from Jojoba seeds in KSA. Journal of Entomology 14(2):73-80. |
|
|
Adehan SB, Biguezoton A, Adakal H, Dossa F, Dougnon, TJ, Youssao E, Sessou P (2016). Acaricidal Activity of Ethanolic and Volatile Extracts of The Leaves of Selected Plants Used in Veterinary Pharmacopeia on The Larvae of Rhipicephalus Microplus in Benin. Alexandria Journal of Veterinary Sciences 49(1):1-11. |
|
|
Akhtar Y, Isman MB (2013). Plant Natural Products for Pest Management: The Magic of Mixtures BT - Advanced Technologies for Managing Insect Pests (I. Ishaaya, S. R. Palli, and A. R. Horowitz, eds.). |
|
|
Allam TBA (2015). Evaluation of the Insecticidal Effect of the Essential Oil of Cinnamomum zeylanicum Against Tuta absoluta (Meyrick). International Journal of Innovative Research in Science, Engineering and Technology 4(9):8033-8037. |
|
|
Bendera MM (2007). Isolation and Characterization of Essential Oils from Ocimum americanum, Lantana camara, Lantana trifolia and Tephrosia vogelii. (October). Diss. Egerton University |
|
|
Campbell BE, Pereira R, Koehler PG (2016). Complications with Controlling Insect Eggs. Insecticides Resistance, 61848. https://doi.org/10.5772/61848 |
|
|
Campolo O, Cherif A, Ricupero M, Siscaro G, Grissa-Lebdi, K, Russo A, Palmeri V (2017). Citrus peel essential oil nanoformulations to control the tomato borer, Tuta absoluta: Chemical properties and biological activity. Scientific Reports 7(1). |
|
|
Campos MR, Biondi A, Adiga A, Guedes RNC, Desneux N (2017). From the Western Palaearctic region to beyond: Tuta absoluta 10 years after invading Europe. Journal of Pest Science 90(3):787-796. https://doi.org/10.1007/s10340-017-0867-7 |
|
|
Chau NNB, Tu DTC, Quoc NB (2019). Antifeedant activity of essential oil Lantana camara L. against Spodoptera litura Fabr. (Lepidoptera: Noctuidae) and Plutella xylostella Curtis (Lepidoptera: Plutellidae). Can Tho University Journal of Science 11(1):1. https://doi.org/10.22144/ctu.jen.2019.001 |
|
|
Chegini SG, Abbasipour H, Karimi J, Askarianzadeh A. (2018). Toxicity of Shirazi thyme, Zataria multiflora essential oil to the tomato leaf miner, Tuta absoluta (Lepidoptera: Gelechiidae). International Journal of Tropical Insect Science 38(4):340-347. |
|
|
Cheng SS, Chang HT, Lin CY, Chen PS, Huang CG., Chen WJ, Chang ST (2009). Insecticidal activities of leaf and twig essential oils from Clausena excavata against Aedes aegypti and Aedes albopictus larvae. Pest Management Science 65(3):339-343. https://doi.org/10.1002/ps.1693 |
|
|
Costa JG, Rodrigues FO, Sousa EMS, Junior D, Campos A, Coutinho H, de Lima S (2010). Composition and larvicidal activity of the essential oils of Lantana camara and Lantana montevidensis. In Chemistry of Natural Compounds (Vol. 46). |
|
|
Damalas CA, Koutroubas SD (2018). Current status and recent developments in biopesticide use. Agriculture (Switzerland) 8(1). |
|
|
Deshmukhe PV, Hooli A.A, Holihosur SN (2011). Effect of Lantana camara ( L .) on growth , development and survival of tobacco caterpillar (Spodoptera litura Fabricius )*. Karnataka Journal of Agricultural Sciences 24(2):137-139. |
|
|
Desneux N, Luna MG, Guillemaud T, Urbaneja A (2011). The invasive South American tomato pinworm, Tuta absoluta, continues to spread in Afro-Eurasia and beyond: The new threat to tomato world production. Journal of Pest Science 84(4):403-408. |
|
|
Dos S, RC, De MF, AA Chagas, EA Fernández, IM Takahashi, JA, Ferraz VP (2019). Influence of diurnal variation in the chemical composition and bioactivities of the essential oil from fresh and dried leaves of Lantana camara. Journal of Essential Oil Research 31(3):228-234. |
|
|
Dougnon G, Ito M (2019). Sedative effects of the essential oil from the leaves of Lantana camara occurring in the Republic of Benin via inhalation in mice. Journal of Natural Medicines (0123456789). https://doi.org/10.1007/s11418-019-01358-9 |
|
|
El B, GS, Goda HM, Khalifa EA, Abd EHH (2014). Antimicrobial and antioxidant activities of leaves and flowers essential oils of Egyptian Lantana camara L. Der Pharma Chemica 6(6):246-255. |
|
|
Elshiekh N, Kona M, Taha AK, Mahmoud MEE (2014). Effects of Botanical Extracts of Neem(Azadirachta indica) and Jatropha (Jatropha curcus) on Eggs and Larvae of Tomato Leaf Miner, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Persian Gulf Crop Protection 3(3):41-46. |
|
|
Gill HK, Harsh G(2014). Environmental Impacts and Management Strategies,. In M. L. L. and S. Soloneski (Ed.), Pesticides - Toxic Aspects (pp. 187-229). |
|
|
Gotyal BS, Srivastava C, Walia S (2016). Fumigant Toxicity of Essential Oil from Lantana camara Against Almond Fumigant Toxicity of Essential Oil from Lantana camara Against Almond Moth , Cadra cautella. Journal of Essential Oil Bearing Plants (August). |
|
|
Guedes RNC, Roditakis E, Campos MR, Haddi K, Bielza P, Siqueira HAA, Nauen R (2019). Insecticide resistance in the tomato pinworm Tuta absoluta: patterns, spread, mechanisms, management and outlook. Journal of Pest Science 92(4):1329-1342. |
|
|
Hanem FK (2012). Prospects of botanical biopesticides in insect pest management. Pharmacologia 3:641-656. |
|
|
Javier A MV, Ocampo VR, Ceballo FA, Javier PA (2017). Insecticidal Activity of Selected Essential Oil Extracts Against Common Cutworm, Spodoptera litura Fabricius ( Lepidoptera?: Noctuidae ) 146:247-256. |
|
|
Kasmara H, Melanie N, DA Hermawan, Panatarani C (2018). The toxicity evaluation of prepared Lantana camara nano extract against Spodoptera litura (Lepidoptera: Noctuidae). AIP Conference Proceedings, 1927. |
|
|
Khalid KA(2019). Harvest stages and their influences on Lantana camara L. essential oil. Biocatalysis and Agricultural Biotechnology 22:101403. |
|
|
Khaoula Z (2013). Bio-insecticidal potential of essential oils of two Citrus species against two Greenhouse pests Tuta absoluta. Microbiologie Hygiène Alimentaire 25:84-88. |
|
|
Moreno SC, Carvalho G A, Picanço MC, Morais EGF, Pereira RM (2012). Bioactivity of compounds from Acmella oleracea against Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) and selectivity to two non-target species. Pest Management Science 68(3):386-393. |
|
|
Moustafa MF, Hesham AEL., Quraishi MS, Alrumman S A(2016). Variations in genetic and chemical constituents of Ziziphus spina-christi L. populations grown at various altitudinal zonation up to 2227 m height. Journal of Genetic Engineering and Biotechnology 14(2):349-362. |
|
|
Murugesan S, Rajeshkannan C, Suresh B, D, Sumathi, Manivachakam P (2012). Identification of insecticidal properties in common weed - Lantana camara Linn by Gas Chromatography and Mass Spectrum (GC-MS-MS). Pelagia Research Library 3(5):2754-2759. |
|
|
Murugesan S, Senthilkumar N, Suresh B, Rajasugunasekar D (2016). Chemical constituents and toxicity assessment of the leaf oil of Lantana camara Linn from Tamilnadu regions. Asian Journal of Plant Science and Research 6(3):32-42. |
|
|
Ncube B, Finnie JF, Van SJ (2012). Quality from the field: The impact of environmental factors as quality determinants in medicinal plants. South African Journal of Botany 82:11-20. |
|
|
Nea F, Tanoh EA, Yapi TA, Garcia G, Tomi F, Tonzibo Z. F(2017). Chemical investigation on leaf, flower and fruit oils of lantana camara from Côte d'Ivoire. Natural Product Communications 12(4):607-610. |
|
|
Nuruzzaman M, Liu Y, Rahman MM, Dharmarajan R, Duan L., Uddin AFMJ, Naidu R(2019). Nanobiopesticides: Composition and preparation methods. In Nano-Biopesticides Today and Future Perspectives. |
|
|
Olayemi RF (2017). The Role of Monoterpenoids and Sesqiterpenoids as defense Chemicals in Plants - a Review Olayemi , R . F .: The Role of Monoterpenoids and Sesqiterpenoids as defense Chemicals in Plants - a Review. Nigerian Research Journal of Chemical Sciences 3:1-15. |
|
|
Omoregie E., Aliyu I, Doris E, Ehiabhi O, Folashade K (2016). Chemical Components of the Essential Oil of Lantana camara L. Found in North Central Sudan Region of Nigeria. American Chemical Science Journal 12(1):1-7. |
|
|
Özkara A, Aky?l D, Konuk M (2016). Pesticides, Environmental Pollution, and Health. In M. L. Larramendy and S. Soloneski (Eds.), Environmental Health Risk - Hazardous Factors to Living Species (Vol. 1, p. 13). |
|
|
Pereira KLG, Nogueira PCDL, Arrigoni-Blank MDF, Nizio DAD, Silva DC, Pinto JAO, Blank AF(2019). Chemical diversity of essential oils of Lantana camara L. native populations. Journal of Essential Oil Research 00(00):1-16. |
|
|
Roditakis E, Skarmoutsou C, Staurakaki M, del Rosario Martínez-Aguirre, M., García-Vidal L., Bielza P, Teixeira LA (2013). Determination of baseline susceptibility of European populations of Tuta absoluta (Meyrick) to indoxacarb and chlorantraniliprole using a novel dip bioassay method. Pest Management Science 69(2):217-227. |
|
|
Roditakis E, Vasakis E, García-Vidal, L., Martínez-Aguirre MR, Rison JL., Haxaire-Lutun MO, Bielza P (2018). A four-year survey on insecticide resistance and likelihood of chemical control failure for tomato leaf miner Tuta absoluta in the European/Asian region. Journal of Pest Science 91(1):421-435. |
|
|
Rwomushana I, Beale T, Chipabika G, Day R, Gonzalez-Moreno P, Lamontagne-Godwin J, Makale F, Pratt C, Tambo JR (2019). Tomato leafminer (Tuta absoluta): Impacts and coping strategies for Africa. CABI (Center for Agriculture and Bioscience International) Working Paper. |
|
|
Silva JE, Maria L, Ribeiro S, Vinasco N, Narciso R., Siqueira HÁA (2019). Field ? evolved resistance to chlorantraniliprole in the tomato pinworm Tuta absoluta?: inheritance , cross ? resistance profile , and metabolism. Journal of Pest Science 92(4):1421-1431. |
|
|
Soares MA, Campos MR, Passos LC, Carvalho GA., Haro MM, Lavoir AV, Desneux N (2019). Botanical insecticide and natural enemies: a potential combination for pest management against Tuta absoluta. Journal of Pest Science 92(4):1433-1443. |
|
|
Swagatika P, Smaranika P (2017). AbstractView. Resource Journal of Pharmacognosy and Phytochemistry 9(4):203-209. |
|
|
Swamy MK, Arumugam G, Kaur R, Ghasemzadeh A, Yusoff MM, Sinniah UR (2017). GC-MS Based Metabolite Profiling, Antioxidant and Antimicrobial Properties of Different Solvent Extracts of Malaysian Plectranthus amboinicus Leaves. Evidence-Based Complementary and Alternative Medicine 2017. |
|
|
Syombua ME (2015). Effects of essential oils of Lantana camara and two Ocimum species on bean weevil (Acanthoscelides obtectus) and their chemical compositions. Nairobi: School of Pure and Applied Sciences, Kenyatta University. |
|
|
Yarou BB, Bawin T, Boullis A, Heukin S, Lognay G, Verheggen FJ, Francis F (2018). Oviposition deterrent activity of basil plants and their essentials oils against Tuta absoluta (Lepidoptera: Gelechiidae). Environmental Science and Pollution Research 25(30):29880-29888. https://doi.org/10.1007/s11356-017-9795-6 |
|
|
Yuan Z, Hu XP (2012). Repellent , Antifeedant , and Toxic Activities of Lantana camara Leaf Extract Against Reticulitermes flavipes (Isoptera?: Rhinotermitidae). Journal of Economic Entomology, 105(6), 2115-2121. |
|
|
Zandi-Sohani N, Hojjati M, Carbonell-Barrachina ÁA (2012). Bioactivity ofLantana camara L. Essential Oil Against Callosobruchus maculatus (Fabricus). Chilean Journal of Agricultural Research 72(December): 502-506. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


