Full Length Research Paper

ABSTRACT
Allometric relationships have been used widely as a fast approach to estimate biomass and leaf area in many plant species without destructive sampling. Several allometric functions have been developed to estimate above-ground biomass (AGB) in field-grown East African Highland Bananas (EAHB). However, it is not clear if these functions apply to potted apple bananas hence the need for this study. Plants were randomly selected from different treatments to make a representative sample (n=60) across the range of plant heights and vigor in the trial. For each plant, one fully expanded and unfurled leaf was randomly selected to measure and record its length (L) and width (W) at the widest point. The leaf was drawn to scale on squared paper to determine its true area, which was later compared with the area estimated as the product of L, W and an allometric correction factor. The existing and developed functions were evaluated qualitatively using unit plots and quantitatively using the root mean square error (RMSE) and squared correlation coefficient (r2). Girth at 30 cm (G-30) was the best (p<0.001 and R2 = 89%) input variable for estimating apple banana AGB. It was recommended that leaf area for the potted apple banana plants can be estimated using the existing function but the developed function should estimate AGB with G-30 as the independent variable.
Key words: Destructive sampling, dry matter accumulation, Musa spp-AAB, plant girth, plant height.
INTRODUCTION
Biomass accumulation and leaf area of a plant are key indicators for plant growth and productivity (de la Riva et al., 2016; Lemaire et al., 2019). Leaf area is important in processes like photosynthetic light absorption, carbon uptake and assimilation, transpiration of water, and emission of volatile organic compounds (Fleck et al., 2012). Conventional methods for determining plant biomass accumulation involve destructive sampling, which is undesirable in some studies requiring repeated measures on the same plant (Stevens et al., 2020). On the other hand, the shape of most plant leaves renders ordinary mathematical formulae for determining area rather cumbersome, yet leaf area is an important variable affecting light interception, and hence photosynthesis and carbohydrate production. The use of a correction factor has been advanced to enable estimation of leaf area from conventional formulae for area determination with simple leaf dimensions such as length and width (Awal et al., 2004). Allometric functions have been widely used as a fast approach to estimate biomass and leaf area in the fields of forestry, ecology and horticulture (Nyombi et al., 2009; Ma et al., 2020) without destructive sampling. Allometry is the quantitative relationship between relative dimensions of parts of an organism and its overall size (Gayon, 2000; Niklas, 2004). In trees and bananas, the relative dimensions are usually the easily measurable plant parameters such as girth at the collar (or base), diameter at breast height (DBH), plant height, leaf width and length, and a number of functional leaves (Nyombi et al., 2009; Okore, 2014; Stevens et al., 2020). These parameters have been used to accurately estimate accumulated plant biomass in trees, forage species, coffee, cocoa, mango, soybean, black pepper and bananas among other crops (Tittonell et al., 2005; Kamusingize et al., 2017; Lemaire and Belanger, 2019), leaf area of Grande Naine bananas (Potdar and Pawar, 1991) and East African highland bananas (Nyombi et al., 2009; Stevens et al., 2020).
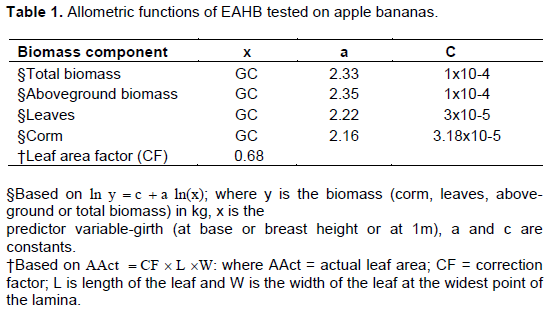
Girth at breast height (DBH) is the commonly used predictor variable (Shem et al., 2013; Kamusinginze et al., 2017) for estimating plant biomass. Other parameters used include girth at the base (Yamaguchi and Araki, 2004; Nyombi et al., 2009), and girth at 100-cm (Wairegi et al., 2009) and use of plant height (Kamusingize et al., 2017). Generally, the equations developed to fit in the general linear equation (Equation 1) with girth as the best predictor variable although they do not agree at what point of girth measurement.

Where, y is biomass (corm, leaves, above-ground or total biomass) in kg, x predictor variable-girth (at base or breast height or at 1m), a and c are constants.
These functions have been used in predicting biomass accumulated in East African Highland cooking bananas but have not been tested for applicability to apple bananas. For this study, functions developed by Nyombi et al. (2009) predicting total biomass, above-ground biomass, the mass of leaves, and the mass of corm (Table 1) were evaluated for applicability in potted apple bananas since these were young plants of less than 100 cm in height. This was necessary because the accuracy of allometric functions’ prediction is species-specific since morphological traits vary with plant species and crop nutrition (Lemaire et al., 2019). Within a diverse species such as bananas, allometric relationships may be expected to differ between cultivars or clone sets of the same species or cultivar (Stevens et al., 2020).
East African highland bananas exhibit morphological traits that markedly differ from those of apple bananas (Karamura et al., 2012). More so, these studies have been done on field-grown banana plants, whose morphology differs from that of small potted plants.
Diameter at breast height (taken at a height between 100 and 130 cm above the ground) was considered as the best explanatory variable for prediction of biomass in several species (Shem et al., 2013; Kamusingize et al., 2017), yet potted plants with confined rooting volume might not attain such height. Therefore, there was a need to test the applicability of existing allometric functions for estimating aboveground biomass and leaf area of potted apple bananas.
General objectives
The broad objective of this study was to identify allometric functions that are suitable for estimating above-ground biomass and leaf area in young potted apple banana plants.
Specific objectives
(1) To test the existing allometric functions for predicting above-ground biomass and leaf area on potted young apple banana plants.
(2) To develop suitable allometric functions for predicting above-ground biomass and leaf area of potted young apple banana plants, if none of the existing functions proves suitable.
MATERIALS AND METHODS
Study area
The study was carried out in Kifu Forest Reserve (00°28’N and 32°44’E at an elevation of 1250 m above sea level). The climate at the Kifu is warm-humid with an average temperature of 25°C and precipitation of 1560 mm per annum distributed in two seasons (March – June, and August – November) (Ochola et al., 2014).
Experimental set up and data collection
A pot experiment was carried out in a screen house set up in Kifu Forest Reserve. Tissue culture plantlets of apple banana (Sukali ndizi) were planted in pots containing 10 kg of sterilized soil. Potassium was applied at 150 kg ha-1 from inorganic fertilizer (Muriate of Potash, 60% K2O), manure and Potassium solubilizing bacteria (KSB). Dry manure was used at a rate equivalent to 150 kg ha-1 and Frateuria auranta was used as the biofertilizer containing the potassium- solubilizing bacteria at a rate of 1 L ha-1. The inputs were applied at planting in full factorial combination resulting in 7 treatments (3 sole inputs, 3 binary combinations of the inputs, and 1 with all the 3 inputs integrated) plus a control that constituted the trial. That is, T1-no input (control), T2-MOP alone, T3-Manure alone, T4-MOP and Manure, T5-KSB alone, T6-MOP and KSB, T7-Manure and KSB, and T8-MOP x Manure x KSB. These were replicated 3 times in a randomized complete design. The experiment was kept weed-free through hand weeding. The pots were kept well-watered at all times.
Sixty plants were randomly selected from all treatments and control such that the sample space spanned a wide range in vigor and morphological traits. The selected plants were further randomly split into two subgroups of 30 plants each. The first subgroup (calibration subgroup) was used in deriving allometric functions for estimating above-ground biomass and leaf area correction factor for estimating leaf area from leaf length and width observations. The second subgroup (evaluation subgroup) was used to evaluate the performance of the generated allometric functions or leaf area correction factor and also to test the reliability of suitable existing allometric functions and correction factor on young potted apple banana plants.
Allometric function for apple banana biomass
For each selected plant, a count of functional (over 50% of the lamina still green and petiole still attached to the pseudostem) and dead leaves (less than 50% of the lamina still green or detached from the pseudostem) was done. Girth at the collar and 30 cm-height and plant height were also recorded. The plants were then carefully uprooted from the pots, washed to remove all soil without losing any roots and excess water mopped off before weighing individually using a digital weighing scale (±0.1 g) to determine total fresh weight. The different biomass parts (roots, corm, pseudostem, and leaves) were separated from each other using transverse cuts as shown in Plate 1. The fresh weight of each biomass part was taken on a digital scale and recorded separately for each plant. Thereafter, a sample was taken from each biomass part and its fresh weight taken on a digital scale and recorded before placing it in a uniquely labeled paper bag for oven drying to constant weight.

After oven-drying to constant weight, the ratio of the sample dry weight to the sample fresh weight was multiplied by the total fresh weight of the biomass part to obtain the dry matter content of the biomass part (Bi et al., 2001; Kamusingize et al., 2017). The dry weights of all biomass parts for a given plant were then summed up to give the accumulated total dry weight for the plant sampled.
Allometric function for apple banana leaf area
From each plant, a leaf was randomly sampled after taking the total fresh weight of the leaves for the plant. The length (L) of the leaf was measured as the distance between the apex and the point of attachment of the leaf lamina to the petiole and recorded. The width (W) of the leaf at the widest part of the lamina was also measured and recorded. These were used to determine apparent leaf area (AApp) as a product of L and W.
The width of the lamina was taken at two leaf points A and B (ensuring that the ruler was perpendicular to the leaf’s horizontal axis) where A was 2 cm from apex and B 2 cm from point of attachment of the lamina to the petiole. The lamina between points A and B was then divided into 10 segments, at each of which width was recorded. Using a suitable scale factor along with the length and width data, the shape of the leaf was drawn on a size A4 squared paper by joining the points demarcating the known leaf widths (Table 1). The actual area (AAct) of the leaf was then determined by the summation of full and truncated squares.
Data analysis
The observed biomass per plant was regressed on their corresponding values predicted (Piñeiro et al., 2008). The performance of each selected allometric function was assessed by comparing the deviation of the slope from 1 and the deviation of the intercept from 0 using t-tests in GenStat 19 (Baird et al., 2017). The performance of the existing allometric functions were further evaluated using the normalized root mean square error (NRMSE) and squared coefficient of correlation (r2) as computed from Equations 2 and 3, respectively (Taulya, 2015).
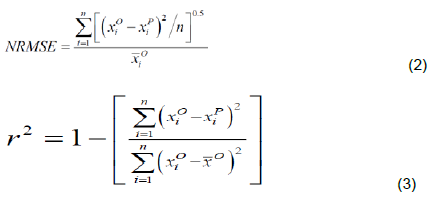
Where, xio and xip are observed and predicted values for the ith sampled plant, respectively and ?0 is the mean of observed values for a given morphological input variable in a sample space of n plants.
Data were natural log-transformed to fit the assumptions of linear regression before running a simple linear regression analysis in GenStat 19 (Baird et al., 2017). This generated allometric functions whose accuracy and precision were assessed using the evaluation dataset as described above for the existing allometric functions.
The observed leaf area (AAct) was regressed on the apparent leaf area (AApp) to estimate the correction factor as the slope of this regression according to Equation 4.
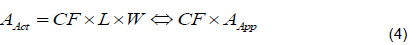
The resulting allometric equation was evaluated for accuracy and precision on the testing dataset following the procedure described above for evaluating the existing allometric functions. All significant results are reported at a 95% confidence level.
RESULTS
Validating the previously developed functions
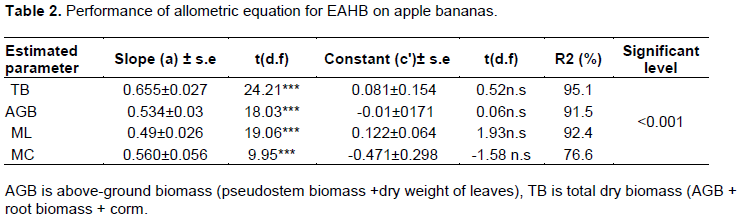
Using girth at the collar (GC) and constants derived by Nyombi et al. (2009) gave statistically significant estimates of biomass (Table 2). However, from the unit plots, the existing functions under-estimated AGB by 15%. The total biomass and corm mass estimates were half the observed values while the leaf biomass estimate a quarter of the observed values (Figure 1).
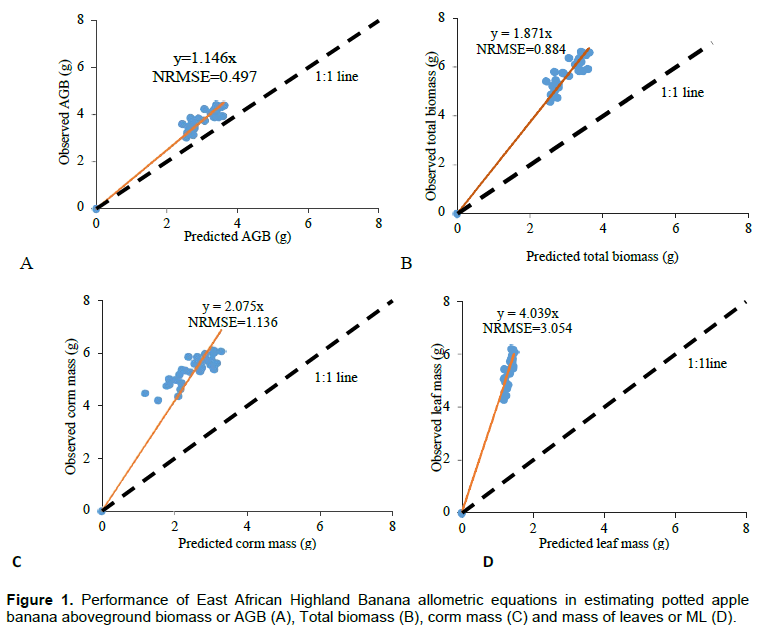
Allometric functions to estimate biomass in apple bananas
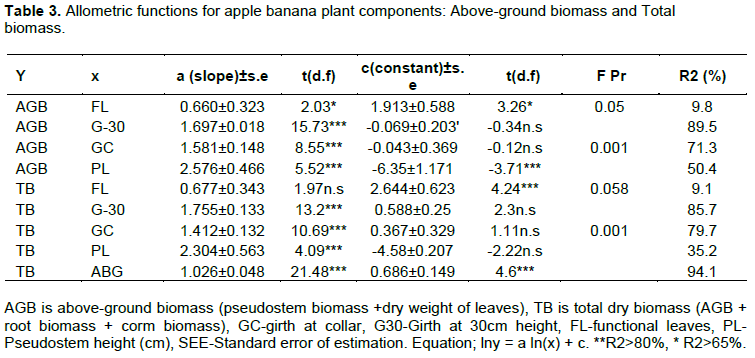
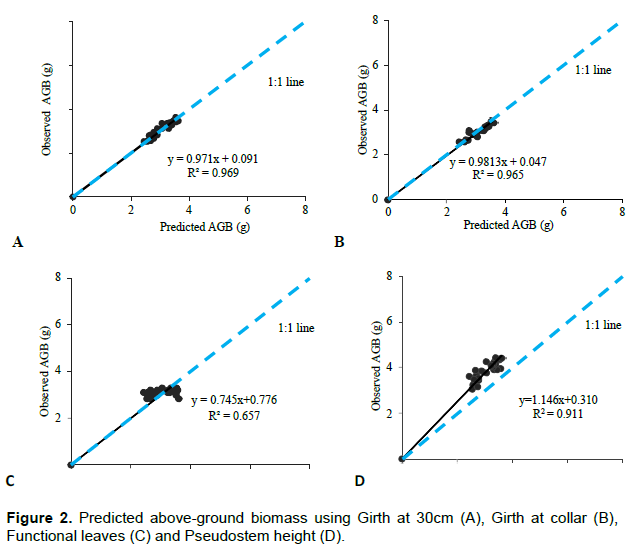
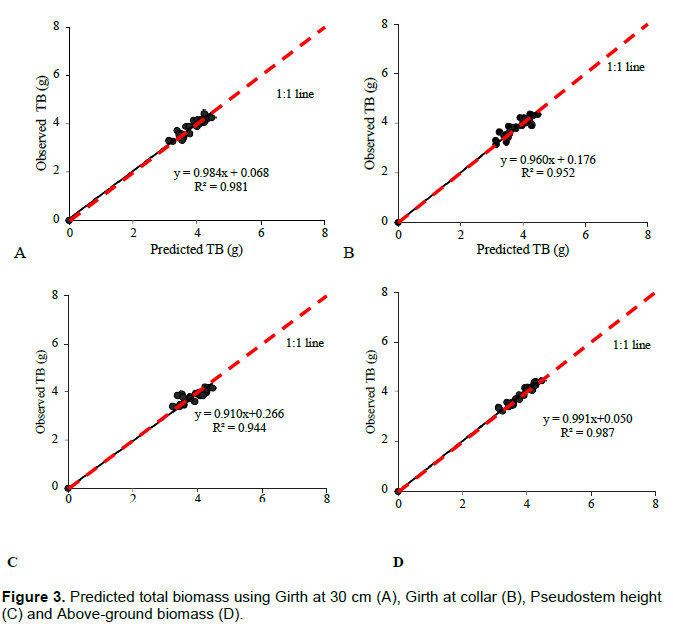
All predictor variables; girth, pseudostem height, and number functional leaves were significantly related to above-ground biomass in apple bananas. However, the number of functional leaves recorded a low regression coefficient (R2) and a high standard error of estimate as compared to other variables. Girth at 30 cm height and girth at the collar recorded significant estimations (p<0.001) for total and above-ground biomass. Pseudostem height also recorded significant estimation of above-ground biomass and total biomass (p<0.001) but with low correlation coefficients of 50.4 and 35.2% for above-ground and total biomass respectively. Above-ground biomass gave a strong regression coefficient in estimating total biomass of 94% (Table 3). A comparison of observed above-ground biomass and the predicted biomass against the 1:1 plot is presented in Figure 2A-D and Figure 3A-D. Girth at 30 cm height and Girth at the collar gave the most suitable estimates of above-ground biomass with coefficients greater than 96%. Functional leaves produced the weakest coefficient of estimation at 65% while a plot of estimates using pseudostems gave higher values than expected. Girth at the collar, girth at 30 cm, pseudostem, and above-ground biomass was also used in estimating total biomass. A comparison against the 1:1 plot is presented in Figure 3A to D.
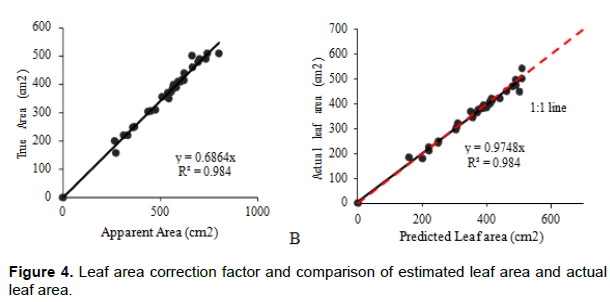
True leaf area was estimated by a leaf area correction factor of 0.68 from the apparent area (Figure 4A). A comparison of true area and estimated leaf area using the determined leaf area correction factor produced a strong positive correlation between the parameters (Figure 4B).
DISCUSSION
The previously developed equations to predict above-ground biomass in EAHB did not accurately predict above-ground biomass of young potted apple banana plants. The equations gave higher predicted estimates than expected (Figure 1 A-D). This could be due to the distinct morphological differences between EAHB and apple bananas. Apple bananas generally have thin pseudostems that support a small bunch of fruit of about 8 kg (Karamura et al., 2012), unlike the EAHB which has stout pseudostems meant to support heavier bunches of fruit. It is also possible that the generated allometric equations could not predict biomass accurately in apple bananas because they were generated on mature plants and included more components that were not catered for in this study.
The leaf area correction factor for apple banana was found to be 0.68 and an R2 of 0.98 (Figure 4A.). This was comparable with what was reported for EAHB (Nyombi et al., 2009), but lower than 0.84 reported in other banana varieties in India (Potdar and Pawar, 1991). Potdar and Pawar (1991) did not explain the protocol used in their study and also did not indicate the width and length range used. The leaf area correction factor generated by Nyombi et al. (2009) for EAHB of 0.68 on the other hand accurately predicted leaf area in apple bananas (Figure 4B). The correction factor by Nyombi et al. (2009) gave accurate predictions possibly because the shapes of banana leaves do not much differ between the mature and potted young plants.
The results obtained in this study indicated that girth at 30 cm height was the best predictor variable followed by girth at the collar to accurately predict above-ground and total biomass, p<0.001 and R2 ranging from 65 to 90%. These results are in agreement with what was obtained by Okore (2014) who found significant above-ground biomass estimation in young and old forest trees using girth at 30 cm height. Kamusingize et al. (2017) found girth at 100 and at 130 cm height to accurately predict total biomass in banana cultivars, Enyeru, Nakinyika, and Nakitembe. The same study identified that girth at collar was not an accurate predictor variable for the total biomass which contradicts the findings of Nyombi et al. (2009). Girth at 130 cm height was also found to accurately predict above-ground biomass in forest trees (Ketterings et al., 2001).It was suggested by Kamusingize et al. (2017) that in absence of girth at breast height (taken at 130 cm height) as in the case of young plants, which were used in this study, pseudostem height could be used as an alternative variable to estimate above-ground biomass. However, this was not the case in this study. The pseudostem height gave moderate estimates of above-ground biomass R2 50%. Contrary to the studies of Ma et al. (2020) where leaf area accurately predicted biomass of Fritillaria przewalskii, functional leaves did not accurately predict above-ground biomass in apple bananas. This study proposes girth at collar as an alternative variable to estimate above- ground biomass in young plants in absence of girth at 30 cm height. This study also indicated that above-ground biomass can accurately be used to estimate total biomass of the entire plant at R2 94%. This could be because above-ground biomass constitutes the biggest percentage of total biomass than below-ground biomass (Okore, 2014).
CONCLUSION
Allometric functions developed for EAHB to estimate biomass did not accurately predict in young apple bananas. The above-ground biomass was predicted by variable parameters in the order G- 30> GC> PL>Functional leaves. However, the same leaf area correction factor can accurately predict leaf area in both EAHB and apple bananas.
To predict above-ground biomass in young apple bananas, girth at 30 cm height was a better parameter than either pseudostem height or girth at the collar. For estimation of above-ground biomass and total biomass in the subsequent studies, allometric equations having girth at 30-cm height as the predictor variable should be considered since it was consistently better than others in this study. Allometric functions developed for EAHB should be further tested on field grown apple bananas to ascertain their applicability. It is recommended that these developed functions be tested for applicability and reliability on grown apple banana plants since they were developed using young ones.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Awal MA, Ishak W, Endan J, Haniff M (2004). Determination of specific leaf area and leaf area-leaf mass relationship in oil palm plantation. Asian Journal of Plant Sciences 3(3):264-268. |
|
|
Baird D, Murray D, Payne R, Soutar D (2017). Introduction to Genstat® for WindowsTM. 19th Edition. VSN International, 2 Amberside, Wood Lane, Hemel Hempstead, Hertfordshire HP2 4TP, UK. |
|
|
de la Riva EG, Olmo M, Poorter H, Ubera. JL, Villar R (2016). Leaf mass area (LMA) and its relationship with leaf structure anatomy in 34 Mediterranean woody species along water availability gradient. PLoS ONE 11:2. |
|
|
Fleck S, Raspe S, Cater M, Shcleppi P,Ukomaanaho L, GreveM, Hertel C, Weis W, Rumpf S (2012). Leaf area Measurements. 37PP, Manual part XVII. In: Manual on harmonized sampling, assessment, Monitoring, and analysis of effects of air pollution on forests. http.//www.icpforests.org/manual.htm. |
|
|
Gayon J (2000). History of the concept of allometry. American Zoologist 40(5):748-758. |
|
|
Kamusingize D, Majaliwa JM, Komutunga E, Tumwebaze S, Nowakunda.K, Namanya P, Kubiriba J (2017). Allometric relationships and carbon content for biomass-carbon estimation of East African Highland Bananas (Musa spp. AAA-EAHB) cv. Kibuzi, Nakitembe, Enyeru and Nakinyika. African Journal of Agricultural Research 12(14):1217-1225. |
|
|
Karamura DA, Karamura E, Tinzaara W (editors) (2012). Banana cultivar: Names, synonyms and their usage in East Africa. Bioversity International, Uganda. |
|
|
Ketterings QM, Coe R, van-Noordwijk M, Ambagau Y, Palm CA (2001). Reducing uncertainty in the use of allometric biomass equations for predicting above-ground tree biomass in mixed secondary forests. Forest Ecology and Management 146:199-209. |
|
|
Lemaire G, Belanger G (2019). Allometries in Plants as Drivers of Forage Nutritive Value: A Review. Agriculture 10(1)5. |
|
|
Lemaire G, Sinclair T, Sadras V, Belanger G (2019). Allometric approach to crop nutrition and implications for crop diagnosis and phenotyping. A review. Agronomy for Sustainable Development 39:27. |
|
|
Ma R, Xu S, Chen Y, Guo F, Wu R (2020). Allometric relationships between leaf and bulb traits of Fritillaria przewalskii Maxim. grown at different altitudes. Plos One 15(10):e0239427. |
|
|
Nyombi K, van Asten PJA, Leffelaar, PA, Corbeels M, Kaizzi CK, Giller KE (2009). Allometric growth relationships of East Africa highland bananas (Musa AAA-EAHB) cv. Kisansa and Mbwazirume. Annals of Applied Biology ISSN 0003-4746. |
|
|
Ochola D, Ocimati W, Tinzaara W, Blomme G, Karamura E (2014). Interactive effects of fertilizer and inoculum concentration on subsequent development of xanthomonas wilt in banana. African Journal of Agricultural Sciences. |
|
|
Okore LMK (2014). The performance of local generic allometries In estimating biomass carbon of trees in agriculturally dominated landscapes of Nyando basin, Kenya. Msc. Thesis, Kenyatta University. |
|
|
Piñeiro G, Perelman S, Guerschman JP, Paruelo JM (2008). How to evaluate models: Observed vs. predicted or predicted vs. observed? Ecological Modelling 216(3-4):316-322. |
|
|
Potdar MY, Pawar K (1991). Non-destructive leaf area estimation in banana. Scientia Horticulturae 45(3-4):251-254. |
|
|
Shem K, Johannes D, Catherine M, van Noordwijk M, Henry N (2013). Allometry and partitioning of above and below-ground biomass in farmed eucalyptus species dominant in Western Kenyan agricultural landscapes. Biomass Bioenergy 55:276-284. |
|
|
Stevens B, Diels J, Brown A, Bayo S, Ndakidemi PA, Swennen R (2020). Banana Biomass Estimation and Yield Forecasting from Non-Destructive Measurements for Two Contrasting Cultivars and Water Regimes. Agronomy 10:1435. |
|
|
Taulya G (2015). Ky'osimba onaanya: Understanding productivity of East African highland banana. PhD thesis, Wageningen University. |
|
|
Tittonell P, Vanlauwe B, Leffelaar PA, Giller KE (2005). Estimating yields of tropical maize genotypes from non-destructive on-farm plant morphological measurements. Agriculture, Ecosystems and Environment 105(1-2):213-220. |
|
|
Wairegi. LWI, van Asten. PJA, Tenywa M, Bekunda M (2009). Quantifying bunch weights of east African highland bananas (Musa AAA-EA) using non-destructive field observations. Scientia Horticulturae 121(1):63-72. |
|
|
Yamaguchi J, Araki S (2004). Biomass production of banana plants in the indigenous farming system of the East African Highland. A case study on the Kamachumu Plateau in northwest Tanzania. Agriculture, Ecosystems and Environment 102(1):93-111. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


