Full Length Research Paper

ABSTRACT
Selection done using molecular markers has spread to all types of agricultural plants and animals. Marker assisted selection (MAS) is safe for ecology and food products consumed. The purpose of the study is to develop Sub1A gene (which determines tolerance to prolonged flooding) in the Russian rice varieties using DNA-markers and Polymerase chain reaction (PCR) analysis. A large volume of pre-breeding rice material (F2-F4, BC2F2) was created with tolerance genes for prolonged flooding, to control weed in rice agrophytocenosis. The forms with 120-130-day vegetation period were selected. An analysis of the co-inheritance of simple sequence repeat (SSR) markers Sub1Aq, Sub1C173, Sub1A203 with the Sub1 gene was carried out. It revealed the codominance of the selected microsatellites. They can be used to identify and visualize the allelic state of a given target gene in the breeding material. The recurrent parental rice forms, donors of the gene Sub 1 and the hybrids of F2 and ВС2F1 population were tested on tolerance to prolonged flooding. The gene is effective in ensuring resistance to this stress factor within 2 to 3 weeks and it can be recommended for the breeding programs to develop modern genetic resources of rice tolerant to prolonged and deep flooding for weed control on rice fields.
Key words: Polymerase chain reaction (PCR), rice, Sub genes, tolerance to flooding, weeds.
INTRODUCTION
According to the occupied areas and a share in the gross volume of the yields rice is the third largest important food crop in the world. More than 3 billion people eat it and it provides 30% of food calories (FAOSTAT, 2016). In the Krasnodar territory, more than 80% of Russian rice is produced on an area of 123-131 thousand hectares; in the Rostov region it occupies 8% of 14 to 15 thousand hectares. The main stress factors limiting the yield of this crop in the rice-growing regions are diseases, pests and weeds.
Some studies show that weed invasion is a problem for the agricultural ecosystems, affecting production. It causes water stress, affects light and nutrients. Water stress is a problem for production as well as light. Many studies recently show that weed is to be managed with feed management. They explain fertilizer and seeds management. Seeding is very important for high yields and feed efficiency. Recent studies show that very high seeding are important to yield (Sevik and Cetin, 2015; Cetin et al., 2018; Tiwari et al., 2014; Torres et al., 2010; Goncharova et al., 2018; Kostylev et al., 2017).
Weeds compete for elements of nutrition, light and other factors with rice resulting in 20-50% of rice productivity shortage. Chemical protection of plants in some cases is neither efficient nor profitable (Kostylev et al., 2015).
Therefore, to eliminate weeds on rice fields, comprehensive solutions are extremely needed that could significantly reduce the use of herbicides, improve the quality of products and the profitability of the rice industry. In rice-growing, the production of sprouts from under the water layer, at which the malicious rice weeds (the Echinochloa species) die is considered to be the most environmentally friendly technology. However, its use causes a number of problems, primarily related to the high mortality of rice seedlings due to the lack of oxygen caused by the increase of the water level for weed control.
Therefore, an effective way to protect rice without herbicides is to grow varieties that are resistant to anaerobic germination and prolonged flooding (Dubina et al., 2017a).
The rice tolerance to flooding is controlled by the Sub1 (Submergence) locus, which was discovered by Xu and Mackill (1996) and Xu et al. (2006). It regulates the reaction to ethylene and gibberellin, which results in a restriction of the consumption of carbohydrates and the calmness of sprouts under water; it also promotes tolerance to prolonged and deep immersion. Expression of this gene increases with an increased concentration of ethylene (in flooding), which results in a decrease of gibberellin induced for plant growth.
Xu and Mackill (1996), using the population obtained from the cross-over of the rice-resistant rice line IR40931-26 (subspecies indica) and the susceptible line P154385 (japonica subspecies) in 1996, mapped the main QTL on the 9th chromosome near its centromere between the RLFP markers C1232 and RZ698 and gave them the designation Sub1 (Figure 1).
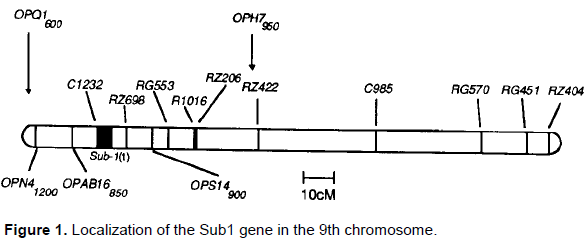
For the genetic mapping Xu et al. (2006) mapped this locus on the 0.06 morganide interval using a population of 4022 plants derived from the hybridization of the resistant variety FR13A (IR40931-26) (subspecies indica) and the unstable variety M.202 (subspecies japonica).
The Sub1 region is bounded by the CR25K and SSR1A markers and covers more than 182,000 base pairs. This interval codes three ethylene response factors, labeled as Sub1A, Sub1B and Sub1C, but only the Sub1A gene enhances plant tolerance to flooding (Mackill et al., 2012).
The International Rice Institute (IRRI, Philippines) worked on identifying the varieties tolerant to prolonged flooding for several decades (Hille, 1982). The Philippine colleagues have recently combined different resistance genes for prolonged flooding, including Sub1. On the genetic basis of the short rice variety ‘Swarna’, the first “super variety” ‘Swarna-Sub1’ with the Sub1 locus was developed. Based on the Sub1 sequent, there have been developed some specific molecular markers that greatly facilitate the marker assisted backcrossing system (MABC), which has been successfully introduced into the breeding process for the development and cultivation of “super-varieties” in Asia and Africa (Mackill et al., 1993; Septiningsih et al., 2009).
Due to MABC, the high productivity, as well as the consuming and nutritional quality of these “super-varieties” was maintained, which stimulates their demand among producers (Septiningsih et al., 2009). By 2011 eight such varieties had already been developed.
The purpose of the study is to develop rice breeding forms with the gene Sub1A, which determines tolerance to prolonged flooding, on the genetic basis of Russian varieties using DNA-markers and PCR analysis.
The development of rice varieties tolerant to this abiotic factor of the environment is of great necessity today. Their cultivation will significantly reduce the costs of rice producers, as it will allow reducing the use of pesticides, avoiding ecosystem pollution as well as increasing the ecological status of rice growing in the Russian Federation.
MATERIALS AND METHODS
The work was carried out in the laboratory of biotechnology and molecular biology of the FSBSI ‘All-Russia Research Institute of Rice’ and in the laboratory of rice breeding and seed-growing of the FSBSI ‘Agricultural Research Center ‘Donskoy’’. Five varieties of foreign selection ‘Khan Dan’, ‘BR-11’, ‘CR-1009’, ‘Inbara-3’, ‘TDK-1’ (Vietnam) of the subspecies of indica with a vegetation period of 156-160 days have become the donors for the introduction of the Sub1A gene into Russian rice geneplasm (Azarin et al., 2016). In the south of Russia, it is preferable to cultivate varieties that mature not more than 125 days. As maternal forms for hybridization, the followings were selected: an early-ripening variety ‘Novator’ with a vegetation period of 105 to 110 days (developed by the All-Russian Research Institute of Rice); large-kernelled early ripening lines ‘VNIIR-9678’, ‘KP-25-14’, and rice lines ‘PK-171 (Pi-ta)’, ‘KP-62 (Pi-2)’ with blast resistance genes (developed by the All-Russian Research Institute of Rice), which allow obtaining rice geneplasm with complex resistance to biotic and abiotic factors of the environment. Hybridization of plants was carried out by pneumo-emasculation. Panicles were pollinated on the day of emasculation by the TVELL method (Los, 1987). For molecular genetic studies, a vegetative DNA was isolated from green leaves of the analyzed rice samples using the Murray CTAB method (Murray et al., 1980). Polymerase chain reaction (PCR) was carried out according to a standard procedure with 40-50 ng DNA, in a final volume of 25 μl in the DNA-amplifier ‘Tercik’ (Kozhukhova, 1998). For establishing of the PCR, optimal conditions were selected and a protocol was drawn up where the output of the amplified product along with the minimum number of synthesized nonspecific DNA fragments was really high, that is initial denaturation is 5 min at 94°C (1 cycle). As for the next 35 cycles, denaturation step is 35 s at 94°C; primers annealing is 45 s at 60°C; synthesis is 30 s at 72°C. The synthesis lasts for 5 min at 72°C per 1 cycle. To identify the Sub1A gene, 76 flanking microsatellite SSR markers taken from the literature sources were used (Bailey-Serres et al., 2010; Xu et al., 2006; Yu et al., 2002; McCouch et al., 2001). Their sequence is presented in the NCBI genetic resources database (www.ncbi.nih.gov). Separation of amplification products with microsatellite markers was performed by electrophoresis in an 8% polyacrylamide gel. The electric field strength at electrophoresis was 3.9 to 4.5 V/cm. After electrophoresis, the gel was stained in a solution of ethidium bromide and photographed under ultraviolet light. To determine the length of the amplified fragments, a molecular weight marker was used (100 bp + 1.5 Kb + 3 Kb (SibEnZim) (0.05 g/L)) (Dubina et al., 2017b). Testing of recurrent and recipient forms of rice, hybrids, as well as weed-field forms of Echinochloa was carried out in the concrete tanks (1.0 mx 2.5 m). At the vegetative stage of rice plants, 5-6 leaves were deeply buried (50 cm of a water layer). After 14 days of flooding, the water was slowly pumped out. On the third day after the removal of water from the reservoir, the survived rice plants and weed species were counted.
RESULTS AND DISCUSSION
The need to create genetic sources of agricultural plants, offered for cultivation by environmental technologies, is constantly increasing. The Sub1A gene enhances rice tolerance to prolonged flooding, which can be used as an environmentally friendly factor for controlling weeds on rice fields.
At the first stage of the work, we searched for the information and made a selection of molecular (microsatellite) markers in the database www.ncbi.nih.gov to identify and control the allelic state of the Sub1A gene in Russian rice varieties and donor varieties by PCR. Among 76 SSR markers tested for the presence of the Sub1A gene in the genotype of rice donor and maternal forms, 3 of the 4 microsatellites (Sub1Aq, Sub1C173, Sub1A203) showed the greatest level of polymorphism. Figure 2 shows the results of their testing. Sub1B showed no difference with the recessive alleles.

The selected SSR markers can be used in breeding programs to create rice varieties tolerant to this stress for identifying and visualizing the allelic state of the Sub1 gene in the hybrid rice plants developed by PCR.
At the next stage, the donors of Sub1A genes were hybridized tolerant to prolonged flooding with the varieties developed by the All-Russian Research Institute of Rice, which have high grain qualitative characteristics. There was obtained F1 generation characterized by a high degree of sterility (90-95%) and brown color of flowering scales during maturation that indicates significant genetic differences between the parental forms. In the second generation, a wide spectrum of splitting was observed for all combinations in vegetative period, plant height, length and shape of panicle, number of spikelets, and awnness.
In the ‘Novator × Khan Dan’ combination in F2, 184 plants were obtained, among which there were identified the samples combining the short height, resistance to shed its grain and fertility of the spikelets. Using DNA-marking, homozygous and heterozygous forms for the Sub1A gene were selected among these samples for further breeding (Figure 3).
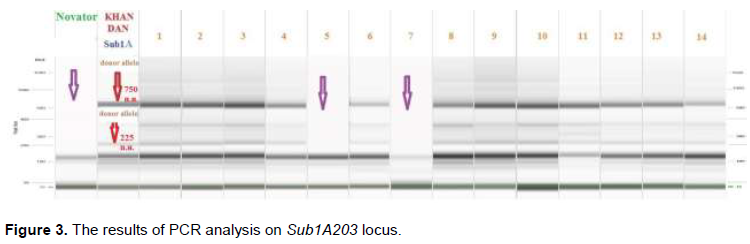
Figure 3 shows that the hybrid plants N° 5 and 7 have a DNA profile and carry the allele of maternal form ‘Novel’; the plant N° 11 has a specific allele of the Sub1A gene in the homozygous state.
The molecular mass of the marker Sub1A203 is 203 bp. All other studied hybrid plants at this locus are heterozygotes. The plants that do not have a target gene according to the PCR analysis were rejected.
Based on the results obtained by the marker analysis of 184 plants, the following ratio was obtained: 39 plants carry a dominant allele in the homozygous state, 104 plants carry a dominant allele in the heterozygous, 41 plants carry homozygotes according to the recessive, which corresponds to a monogenic Mendelian split 1:2:1. This confirms the codominance of the SSR-marker Sub1A203, selected after approbation and its co-inheritance with the gene Sub1A.
Among the hybrids ‘BR-11 × Novator’ analyzed by the SSR-marker RM7481, Sub 1A gene (in the homo- and heterozygous state) was present in nine hybrids N° 4, 7, 8, 9, 11, 12, 13, 18, 19 in a ratio of 9:11, although it should have been 15:5 with a monohybrid splitting.
In a hybrid combination ‘CR-1009×Novator’, DNA analysis data showed splitting in F2 in a ratio of 18:2, that is, almost all selected plants had the Sub 1A gene. The hybrids ‘Inbara-3×Innovator’ and ‘TDK-1×Novator’ had a splitting of 14:6, or about 3:1, that is, closer to the Mendelian. Deviations in the splitting of two combinations can be explained by the effect of selection and the cohesion of genes.
Among 264 plants of five hybrid combinations, 198 plants were identified with the target gene. The selected samples were sown at the field belonging to the Proletarskaya Experimental Station in the Rostov Region and on the lysimetric plots of the FSBSI ‘ARRI of Rice’ to evaluate economic characteristics.
The annual selection of basic plants from hybrid populations F3-F5 with a set of economically valuable traits was accompanied by a PCR analysis to identify the Sub1A gene in the laboratory of biotechnology and molecular biology of the FSBSI ‘ARRI of Rice’. In 2017 more than 100 F5 families were grown and propagated in each hybrid combination.
As the donor varieties have 156-day vegetation period in the conditions of the south of Russia, in F1 there were obtained hybrid plants with 140-day vegetation period, and in F2 there was a wide range of variability from early maturity (‘Novator’ type) to very late (at the end of September) flowering types. The forms with 120-130-day vegetation period have been selected for further work.
As earlier discussed, in the local rice cultivation area it is preferable to cultivate varieties with a vegetation period of no more than 125 days; therefore, work was done on the scheme of saturating crosses (backcrosses) in parallel with the earlier described strategy. The BC1F1 and BC2F1 generation has been obtained. This will allow creating rice lines with a set of economically valuable signs adapted to the growing conditions in southern Russia (with a vegetation period of not more than 125 days). According to the DNA analysis, heterozygous hybrid plants that will be involved in the further backcrossing were selected.
The next stage of this work was to test the donor variety ‘Khan Dun’ with the gene Sub1A, hybrids F2, BC2F1, as well as the Russian rice varieties and lines ‘Novator’, ‘VNIIR 9678’, ‘KP-25-14’ for tolerance to prolonged flooding. The test was carried out over the rice plants with 5 to 6 leaves in vegetation vessels in concrete tanks (lysimeters) under full flooding (50 cm water layer). Sowings were done by hand. The seeds of a hedgehog (chicken millet) Echinochloa species were also sown in the lysimeters. After 14 days the water was removed from the lysimeters, and on the third day after water discharge, continuation of the development or death of rice and weed plants were assessed (Figure 4).

From the experiment, the rice hybrids and the donor variety ‘Khan Dun’ with the Sub1A gene showed increased survival in the conditions of long-term and prolonged flooding in comparison with plants of Russian varieties. The donor variety and hybrids restored their vital functions after flooding more quickly, producing more new leaves. The plants of the tested Russian rice varieties with 5-6 leaves were weak, depressed after reconstitution, and some of them died. The survived plants poorly formed the vegetative mass. The formed panicles were small in size (5 to 7 cm) and had 40% of sterile kernels. It was noted that on the experimental plots the weeds died under water.
Thus, a preliminary assessment of the obtained hybrids, Russian rice varieties and the donor of the Sub1 gene for tolerance to prolonged flooding has shown that this gene is effective for providing tolerance to this stress factor for 2 to 3 weeks and it should be recommended for breeding programs to develop modern genetic resources of rice, tolerant to prolonged and deep flooding, as a factor of weeds control.
CONCLUSION
1. The information was searched and a selection of informative molecular (microsatellite) markers was made in the database www.ncbi.nih.gov to identify the Sub1A gene by PCR in the obtained rice hybrids.
2. The testing of selected SSR-markers was done and their level of polymorphism in the contrasting rice samples was identified. Three SSRs with a high level of polymorphism on the basis of ‘tolerance to prolonged flooding’ were selected.
3. Hybridization of donor varieties of the Sub1A gene with Russian highly productive rice varieties and lines has been done, that is early-ripening variety ‘Novator’, large-kernelled early-ripening lines ‘VNIIR-9678’, ‘KP-25-14’, as well as with the lines ‘KP-171’ and ‘KP-62’ with blast resistant genes Pi-ta and Pi-2, respectively. The generations F1-F4 and BC2F1 were obtained.
4. DNA-analysis of the obtained hybrids on the identification of the Sub1A gene was done. The plants having a target gene in the hetero- and homozygous state in the genotype were selected.
5. The co-inheritance of the Sub1 gene linked to the sign of ‘tolerance to prolonged flooding’ was studied based on PCR analysis and the codetermination of the selected SSR-markers was established.
6. The testing of the selection rice resources on prolonged flooding resistance during the vegetative experiment was done. The tolerant forms of rice were selected and approved for use in the breeding process to create new rice varieties with valuable agricultural characteristics.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
The study was carried out with the support of the FSBE ‘RFFI’ grant ‘r…a’ No. 16-44-230435 "Development of the methodological foundations of the DNA identification of rice tolerant genes for prolonged flooding as a factor in the ecological weed control and the creation of genetic resources for the selection of resistant varieties ‘Oryza sativa L.’ in cooperation with the All-Russian Scientific Research Institute of Rice (Krasnodar) and the Agricultural Research Center ‘Donskoy’ (Zernograd).
REFERENCES
|
Azarin KV, Usatov AV, Kostylev PI, Makarenko MS, Kovalevich AA (2016). SSR-markers of rice tolerance to prolonged flooding. Grain Economy of Russia 4(46):21-24. |
|
|
Bailey-Serres J, Fukao T, Ronald P, Ismail A, Heuer S, Mackill D (2010). Submergence Tolerant Rice: SUB1's Journey from Landrace to Modern Cultivar. Rice 3:138-147. |
|
|
Cetin M, Sevik H, Yigit N, Ozel HB, Aricak B, Varol T (2018). The variable of leaf micromorphogical characters on grown in distinct climate conditions in some landscape plants. Fresenius Environmental Bulletin 27(5):3206-3211. |
|
|
Dubina EV, Shilovsky VN, Garkusha SV, Kostylev PI, Esaulova LV, Avakyan ER, Balyasny IV (2017b). The role of the gene Sub1A in the rice breeding on tolerance to prolonged flooding. Science of the Kuban 1:77. |
|
|
Dubina EV, Shilovsky VN, Kostylev PI, Garkusha SV, Kovalev VS, Esaulova LV, Balyasny IV, Strakholysova MG, Din TS, Le LN (2017a). The gene Sub1A in the rice breeding on tolerance to flooding as a factor of weed control. Rice Growing 2(35):20-26. |
|
|
FAOSTAT (2016). Crops |
|
|
Goncharova JK, Haritonov EM, Maljuchenko EA, Bushman NJ (2018). Genetics of the traits defining the quality of Russian rice varieties. Vavilovskii Zhurnal Genetiki i Selektsii 22(1):79-87. |
|
|
Hille Ris Lambers D, Seshu DV (1982). Some ideas on breeding procedures and requirements for deepwater rice improvement. In: Proceedings of the 1981 International Deepwater Rice Workshop. International Rice Research Institute. Los Banos, Philippines pp. 29-44. |
|
|
Kostylev PI, Krasnova EV, Red'Kin AA, Dubina EV, Mukhina ZM (2017). Combination of rice blast resistance genes in the genotypes of Russian rice varieties with the use of marker assisted selection. Ecological Genetics 15(3):54-63. |
|
|
Kozhukhova NE (1998). Polymerase chain reaction and features of its application for the analysis of polymorphism. The use of PCR analysis in genetic-breeding studies. Scientific and methodical guidance. Agricultural Science pp. 21-33. |
|
|
Los GD (1987). A promising method of rice hybridization. Agricultural Biology 12:15-17. |
|
|
Mackill DJ, Amante MM, Vergara BS, Sarkarung S (1993). Improved semidwarf rice lines with tolerance to submergence of seedlings. Crop Science 33:749-753. |
|
|
Mackill DJ, Ismail AM, Singh US, Labios RV, Paris TR (2012): Development and rapid adoption of submergence-tolerant (Sub1) rice varieties. Advances in Agronomy 115:303-356. |
|
|
McCouch SR, Temnykh S, Lukashova A, Coburn J, DeClerck G (2001). Microsatellite markers in rice: abundance, diversity, and applications. Rice genetic 4. Proceeding of the fourth international rice genetic symposium. Los Banos pp. 117-135. |
|
|
Murray MG, Thompson WF (1980). Rapid isolation of high molecular weight plant DNA. Nucleic Acids Research 10:4321-4325. |
|
|
Septiningsih EM, Pamplona AM, Sanchez DL, Neeraja CN, Vergara GV, Heuer S, Ismail AM, Mackill DJ (2009). Development of submergence-tolerant rice cultivars: the Sub1 locus and beyond. Annals in Botany 103:151-160. |
|
|
Sevik H, Cetin M (2015). Effects of water stress on seed germination for select landscape plants. Polish Journal of Environmental Studies 24(2):689-693. |
|
|
Tiwari S, Tomar RS, Chand S, Singh NK (2014). Combining multiple rust resistance genes by phenotypic and marker assisted selection in wheat (triticum aestivum l). Indian Journal of Genetics and Plant Breeding 74(2):181-188. |
|
|
Torres AM, Avila CM, Gutierrez N, Palomino C, Moreno MT, Cubero JI (2010). Marker-assisted selection in faba bean (Vicia faba L). Field Crops Research 115(3):243-252. |
|
|
Xu K, Mackill DJ (1996). A major locus for submergence tolerance mapped on rice chromosome 9. Molecular Breeding 2:219-224. |
|
|
Xu K, Xu X, Fukao T, Canlas P, Maghirang-Rodriguez R, Heuer S, Ismail AM, Bailey-Serres J, Ronald PC, Mackill DJ (2006). Sub1A is an ethylene-response-factor like gene that confers submergence tolerance to rice. Nature 442:705-708. |
|
|
Yu J, Hu S, Wang J, Wong GK, Li S, Deng Y (2002). A draft sequence of the rice genome (Oryza sativa L. ssp. indica). Science 296:79-91. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


