ABSTRACT
This experiment aimed to assess the development, yield and chemical composition of Petroselinum crispum essential oil (EO), popularly known as parsley under different levels of water stress deficit: 30 to 40% (moderate stress), 50 to 60% (severe stress) and control 0 to 10%. The plants were kept in a greenhouse to complete the cycle, harvested and measured for their biomass (g), development (cm), yield (%) and chemical composition of EO of aerial parts and root. The EO was extracted by hydrodistillation and the chemical analysis done by gas chromatography/mass spectrometry (GC/MS). In the control and moderate stress, greater development of the aerial part occurred without showing difference in the root development. A yield increase of EO extracted from the roots was verified when it was submitted to moderate stress, but there was no difference in the aerial part yield. The EO chemical composition was influenced by the different conditions of water deficit, producing 100% of apiole in moderate stress and control of aerial parts and roots. A different behavior was observed for severe stress, which presented three compounds (89.98% apiole, 6.53% β-sesquiphellandrene and 3.49% myristicin) in aerial parts, but four compounds were found for the root (88.40% apiole, 5.83% β-sesquiphellandrene, 3.61% myristicin and 2.16% elemicin). These results indicated that under greater water deficit, the plant produced other compounds besides apiole, probably as a defense mechanism. The results showed that there was influence of the water stress on the plant development as well as on the essential oil yield and chemical composition.
Key words: Apiole, gas chromatography/mass spectrometry (GC/MS), essential oil, water stress, parsley.
The species Petroselinum crispum (Mill.) belongs to the family Apiaceae, popularly known as parsley. This plant is an erect, evergreen herb with strong aromatic compound leaves, and inflorescences in the shape of terminal umbels over the leaves, with small yellow-greenish flowers. The fruits are achenes, and the plants propagate by seeds, reaching 15 to 30 cm of height (Lorenzi and Matos, 2008; Vora et al., 2009). Parsley stands out because it is one of the most consumed herbs in the world. It can be utilized either in the food industry or as medicinal plant. It can be used fresh or dehydrated, and in that case, the most consumed parts are leaves, petioles and seeds. The plants develop mostly on sandy clay soils with high content of organic matter, good fertility and pH between 5.8 and 6.8 (Heredia et al., 2003). The essential oil (EO) utilized to aromatize foods is extracted from seeds, roots and leaves. The chemical composition of this oil consists of pinene, myrcene, phellandrene, cymene, methatriene, elemene, myristicin and apiole (Petropoulos et al., 2008).

Because of the worldwide demand for raw materials, essential oils have had an important role in the food, drug, cosmetic and perfume industries. They can substitute or act in synergism with chemical compounds, mainly the synthetic ones, since they have antimicrobial potential (Farag et al., 1989; Kamel, 2000; Tzakou et al., 2001), besides having immomodulating function, antioxidant and food preservative properties (Farag et al., 1989).
During their life cycle, plants are exposed to adverse conditions; among them are the different water stresses. Water deficit is one of the limiting factors of vegetal growth and development, since water works as a reagent in most metabolic reactions (Krieg, 1993). Several processes are altered in the plant due to water deficit such as gas exchange, carbohydrate metabolism, amino acids and other organic compounds (Pagter et al., 2005). Therefore, to survive under such environmental conditions, plants have developed a complex genetic-molecular and cellular signaling network that makes them respond through different metabolic pathways (Guo et al., 2011).
Secondary metabolites are involved in different defense mechanisms under abiotic pressures, and water deficit is among the main causes of yield loss, but presents direct correlation with the concentration of secondary metabolites. There are reports in the literature that water stress usually induces the increase in the concentration of some terpenoids (Morais, 2009). Studies evaluating the influence of water stress on the production of P. crispum essential oil showed that under deficit conditions of 45 to 60%, the concentration of essential oil (EO) was higher (44%) than fresh leaves (Petropoulos et al., 2008).
High concentrations of secondary metabolites resulted in more tolerant plants, but the increase in secondary metabolite production represents a high energetic demand from the plant which consequently reduces its growth, production, biomass and reproduction (Eulgem et al., 1999). This metabolite increase, usually under deficit situations, made plants evolve due to their induced defense system. Thus, in order to evaluate the influence of water stress on parsley development, this study aimed to measure the development, yield and chemical composition of EO of P. crispum under different conditions of water deficit.
Soil preparation
The utilized soil presents sandy superficial texture with low contents of clay and organic matter, and is classified as distroferric red latosol (Fidalski et al., 2013). Thus, it was necessary to prepare the soil since parsley needs organic matter to develop itself (Heredia et al., 2003). The soil was prepared using a mixture containing 65% of manure and 35% of soil with 710 µm granulometry. Later, it was sent for analysis to Laboratório Solo Fértil (CRQ-03751 and CRF-PR 015963/0) located in the city of Umuarama, PR, Brazil (Table 1).
Obtaining plant vegetal
In November, 21-day-old seedlings of Portuguese P. crispum cultivar, approximate size of 4.0 cm, were collected from their seedbed and transferred to 2 L vases. The experiment had random design, was carried out in a greenhouse located in Paranaense University - UNIPAR, in the city of Umuarama, northwestern region of Paraná Sate, with coordinates S23° 46.225’ and WO 53° 16.730’ and altitude of 391 m. A voucher specimen was deposited at the UNIPAR Herbarium (code number 192). After a month, the plants were submitted to two treatments of water deficit in which irrigation was applied at soil water potentials of 30 to 40% (moderate stress) and of 50 to 60% (severe stress), whereas the control was 0 to 10% (control). These percentages represent 0% of soil in field capacity and 100% of dry soil (Petropoulos et al., 2008). 40 vases were utilized for each treatment. The plants were kept in vases until reaching flowering, when they were harvested. The length (cm) and fresh mass (g) of their roots and aerial parts were measured.
Extraction of essential oil (EO)
The fresh material from aerial part and roots was submitted to hydrodistillation (Stanković et al., 2005; Petropoulos et al., 2008) for three hours, and the extractions were done in triplicate for each treatment. The oil was removed from the device with hexane (PA) and filtered with anhydrous Na2SO4, stored in amber flasks under refrigeration and open until hexane had evaporated completely. After total evaporation of the solvent, each extract was weighed to calculate oil yield (%).
Physical and chemical indexes of essential oil obtained by hydro distillation
The refraction and relative density indexes of P. crispum essential oil were determined. The tests were carried out in the Laboratory of Natural Product Chemistry of UNIPAR, Paranaense University, Campus of Umuarama, Brazil. The refraction index was determined by a ABBE-type refractometer, model RL3, PZO Warszawa, at 20°C (FARMACOPÉIA BRASILEIRA, 1988). A substance relative density is its mass ratio by the same water volume mass, both at 20°C according to the technique described in the Brazilian Pharmacopeia (FARMACOPÉIA BRASILEIRA, 1988).
Determining chemical compounds of essential oil through GC/MS
The GC-MS analyses were performed utilizing an Agilent GC, 6890 model, and for MS, 5973 Network Mass Selective Detector. The column had 5% phenyl methylsiloxane (DB-5) and its size was (30 m × 0.25 mm × 0.25 µm). The injector temperature was 250°C in “Split” mode 2.1:1, and the carrier gas was helium with a flow of 4.8 ml/min. The temperatures of the transfer line, ion source and quadrupole were 320, 230 and 150°C, respectively. The column temperature was programmed for initial temperature of 40°C, heating rate of 6°C/min, and final temperature of 300°C (1 min), total analysis time of 44.33 min. The detection system was MS in “Scan” mode, in the mass/load ratio range (m/z) of 50 to 550, with "Solvent Delay" of 3.5 min (Petropoulos et al., 2008). The injections were done in triplicate for each irrigation pattern.
Statistical analysis
Data descriptive analysis was done and the data were expressed by determining the averages and standard error through analysis of variance (ANOVA) and Tukey’s test at 5% significance utilizing Miinitab®17 software
This study evaluated the influence of two different treatments of water deficit in the development of aerial parts and roots (Table 2), and it was observed that different water stress conditions of 30 to 40% (moderate stress) and 50 to 60% (severe stress) significantly influenced the development of aerial parts, but did not change the development of roots. In the aerial parts, moderate stress and control presented greater masses (128.33 and 144.54 g, respectively) but did not differ between them when compared to severe stress that presented smaller mass (90.36 g). Plants with less irrigation (severe stress) had smaller biomass development, probably due to the low water levels that caused stomata closing, decreasing photosynthesis rate and consequently, plant biomass (Tardieu et al., 1992). There was no significant difference for the root length (cm) and mass (g) in the different treatments (Table 2). Cultivation in vases with greater density may produce smaller root biomass and length due to the smaller space for the plant growth (Marotti et al., 1994).
Studies carried out by Petropoulos et al. (2008) evaluated the development of biomass (g) of aerial parts and roots of parsley submitted to water deficit levels of (0 to 10%) control; (30 to 45%) level 1 and (45 to 60%) level 2, resulting in greater biomass of aerial parts (62.3 g) in the control when compared to level 1 (32.8 g) and level 2 (29.6 g). These results corroborate others found in this experiment in which lower irrigation levels induced the decrease of plant biomass and resulted in (144.41 g) for control, in (128.33 g) for moderate stress and in (90.36 g) for severe stress. For the roots, Petropoulos et al. (2008) found biomass values (17.8, 11.3 and 7.3 g) inversely proportional to the increasing order of water deficit. These values are different from the ones found in this experiment in which there was inverse relation but not significant difference of the root biomass under the different conditions of water stress (145.41, 137.00 and 109.34 g).
Bettaieb et al. (2009) analyzed the influence of water deficit levels that were severe (75%), moderate (50%) and control (0%) in Salvia officinalis plant, evaluating the parameters of aerial part biomass (g) and length (cm), and found a reduction of biomass (4.96%) and length (14.15 cm) under severe water deficit condition (12.63%, 19.78 cm) for moderate water deficit, and (19.53%, 26.50 cm) for control, respectively. These results are in accordance with the present experiment, making it evident that the greater the water deficit the slower the plant development.
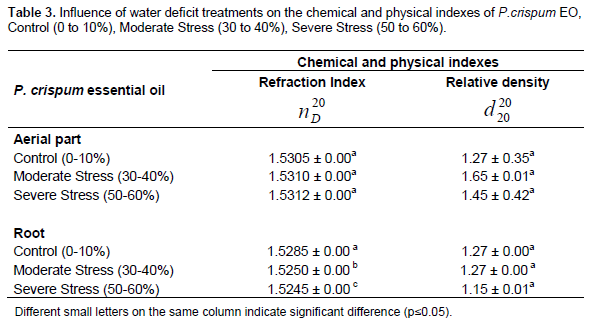
This experiment evaluated the influence of water deficit in the physical and chemical indexes of parsley, measuring the density (g ml-1) and the refraction index, as shown in Table 3. The results showed that there was no significant difference for the density of EO from aerial parts and roots under the evaluated water stress conditions. For the refraction index of EO, there was no difference among the aerial parts; however, there was a significant difference of this parameter of EO in the two evaluated treatments and control. The refraction index has direct correlation with the density, but this relationship was not observed in the results presented in Table 3.
The results found for relative density and refraction index are in accordance with the report by Stanković et al. (2005) who analyzed parsley oil from the region of Belgrade in Serbia, and found density ranging from 1.039 to 1.061 g ml-1, and refraction index varying from 1.52214 to 1.5273.
Regarding water stress in the yield of EO from aerial parts and roots of P. crispum (Figure 1), the yield (%) of EO from aerial parts did not present significant difference between the evaluated water deficit treatments; however, EO from roots showed significant difference between the three water deficit parameters, as shown in Figure 1. There is a greater yield of EO 0.14% for moderate stress and 0.01% for severe stress when compared to (0.05%) for control. These results are in accordance to the ones obtained by Bettaieb et al. (2009) in Salvia officinalis EO. However, Petropoulos et al. (2008) also assessed the yield of EO from aerial parts and root of parsley but did not find a difference between the evaluated water deficit treatments.
The difference of water deficit levels in the yield of Pimpinela anisum EO, also belonging to the family Apiaceae, was studied by Zehtab et al. (2001) who observed greater oil yields when the plant was submitted to a water stress standard below 20%. Farahani et al. (2009) assessed the influence of four water deficit levels (20, 40, 60 and 80%) on Melissa officinalis plant, and found the greatest EO yield at 40% water stress. The cited experiments corroborate the results found in this study in which there was a greater yield of EO from the roots when the plant was submitted to water deficit of 30 to 40%. According to Morais (2009), alterations of the essential oil biosynthesis may function as an adaptive response to water stress, relating some physiological response to environmental variations.
Through the chemical analysis by GC/MS, 10 compounds were found in P. crispum EO from the aerial parts and roots that are listed in elution order in Table 4 and Figure 2. The water deficit of the control and moderate stress made evident the presence of 100% of apiole in EO from aerial parts and roots.
In severe stress, EO from the aerial parts contained apiole (89.98%), β-sesquiphellandrene (6.53%) and myristicin (3.49%) whereas EO from the roots had apiole (88.40%), β-sesquiphellandrene (5.83%), myristicin (3.61%) and elemicin (2.16%), respectively. These results indicated that under greater water deficit (severe stress), the plant responded by decreasing the content of apiole and increasing the amount of myristicin, β-sesquiphelandrene and elemicin, beside apiole, probably as a defense mechanism (Morais and Castanha, 2012). Similar results were found by Ekren et al. (2012) in which there was an increase in the number of Ocimum basilicum oil compounds as there was an increase in water deficit. This increase of compounds may be metabolically explained because the main compounds of parsley oil are made of phenylproanoids that are derived from trans cinamic acid which is formed by phenylalanine aminoacid in a reaction catalyzed by phenylalanine ammonium liase (PAL).The activity of this enzyme in plants is increased as a consequence of biotic and abiotic stresses and then a lot of phenylpropanoids are produced, justifying the increase of myristicin and elemicin when the plant was submitted to the water deficit (Dixon and Paiva, 1995).
Petropoulos et al. (2008) also evaluated the influence of three levels of water deficit: (0 to 10%), (30 to 45%) and (45 to 60%) in the chemical composition of parsley EO from aerial parts and roots cultivated in Greece. Ten main compounds were found: myristicin (28.63, 33.43 and 33.35%) in the aerial parts and (13.55, 16.05 and 7.7%) in the roots, β phellandrene (25.07, 23.63 and 21.97%) in the aerial parts and (20.95, 14.37 and 19.8%) in the roots, and apiole (2.91, 1.90 and 6.35%) in the aerial parts and (23.59, 33.92 and 25.70%) in the roots; β-pinene (16.43, 12.97 and 21.00%) only in the roots. The results from this experiment and from literature (Petropoulos et al., 2008) showed that the chemical composition of parsley EO was influenced directly by the different levels of water deficit (Table 4 and Figure 1). In our experiment, the main compound in all treatments of water deficit was apiole. Petropoulos et al. (2008) found myristicin and β-phellandrene as the main compounds. This difference in the chemical composition probably occurred due to the cultivar, utilized substrate, geographical location, and weather since they are factors that interfere in the chemical composition of EO (Morais and Castanha, 2012).
These results are promising when the experimental objective is to isolate main compounds for commercial purposes because studies done on P. crispum EO showed that phenylpropanoids like apiole and myristicin have antioxidant properties; myristicin can also be utilized as a chemopreventive (Reyes-Munguía et al., 2012) and apiole can be used in several biological activities, mainly antifungal ones (Farag et al., 1989).
This experiment made evident that when the plant was submitted to a greater water stress condition (severe stress) there was a decrease of the aerial part development without interfering in the root development. Moderate stress provided greater yield (%) of EO from roots without changing the yield of aerial parts. The chemical composition of the EO showed apiole as the main compound in all evaluated treatments, 100% for the control and moderate stress. However, for severe stress (greater water deficit), myristicin, elemicin and β-sesquiphellandrene were also found, besides apiole. These results are promising to obtain isolated compounds utilized to determine biological activities and to be utilized in the food and pharmaceutical industries.
The author(s) have not declared any conflict of interests.
REFERENCES
|
Bettaieb I, Zakhama N, Aidi WW, Kchouk ME, Marzouk B (2009). Water deficit effects on Salvia officinalis fatty acids and essential oils composition. Sci. Hortic. 120(2):271-275.
Crossref
|
|
|
|
Dixon RA, Paiva NL (1995).Stress-Induced Phenyl propanoid Metabolism. Plant Cell. 7(7):1085-1097.
Crossref
|
|
|
|
|
Ekren S, Sonmez Ç, Ozçakal E, Kurttas YSK, Bayram E, Gurgulu H (2012). The effect of different irrigation water levels on yield and quality characteristics of purple basil (Ocimum basilicum L.). Agric. Water Manage. 109:155-161.
Crossref
|
|
|
|
|
Eulgem T, Rushton PJ, Schmelzer E, Hahlbrock K, Somssich IE (1999). Early nuclear events in plant defense signaling: rapid gene activation by WRKY transcription factors. EMBO J. 18:4689-4699.
Crossref
|
|
|
|
|
Farag RS, Daw ZY, Hewedi FM, El-Baroty GSA (1989). Antimicrobial activity of some Egyptian spice essential oils. J. Food Prot. 52(9):665-667.
|
|
|
|
|
Farahani HA, Valadabadi SA, Daneshian J, Khalvati MA (2009).Evaluation changing of essential oil of balm (Melissa officinalis L.) under water deficit stress conditions. J. Med. Plant Res. 3(5):329-333.
|
|
|
|
|
FARMACOPÉIA BRASILEIRA (1988). 4ª Ed. São Paulo: Atheneu.
|
|
|
|
|
Fidalski J, Tormena CA, Alves SJ, Auler PAM (2013). Influence of sand fractions on water retention and availability in caiuá and Paranavaí sandstone formations. Rev. Bras. Cienc. Solo. 37(3):613-621.
Crossref
|
|
|
|
|
Guo R, Yu F, Gao Z, An H, Cao X, Guo X (2011). GhWRKY3 a novel cotton (Gossypium hirsutum L.)WRKY gene, is involved in diverse stress responses. Mol. Bio. Rep. 38(1):49-58.
Crossref
|
|
|
|
|
Heredia ZNA, Vieira MC, Weisman LAL (2003). Bunching onion and parsley yield and gross income in mono-cropping and inter-cropping system. Hortic. Bras. 21(3):574-577.
|
|
|
|
|
Kamel C (2000). A novel look at a classic approach of plant extracts (special number) Feed Mix. The Int. J. Feed Nutr. Technol. 9(6):19-24.
|
|
|
|
|
Krieg DR (1993). Stress tolerance mechanisms in above ground organs. Proceedings of the Workshop on Adaptation of Plants to Soil Stress.2ª Ed. Nebraska: Intsormil. pp. 65-79.
|
|
|
|
|
Lorenzi H, Matos FJA (2008). Plantas medicinais no Brasil - Nativas e exóticas.2ªEd. Nova Odessa: Plantarum 544p.
|
|
|
|
|
Marotti M, Piccaglia R, Giovanelli E, Deans SG, Eaglesham E (1994). Effects of planting time and mineral fertilization on peppermint (Mentha x piperita L.) essential oil composition and its biological activity. Flavour Fragr. J. 9(3):125-129.
Crossref
|
|
|
|
|
Morais LAS, Castanha RF (2012). Chemical composition of sweet basil essential oil naturally submitted to Planococcus citri infestation. Hortic. Bras. 30(2):S2178-S2182.
|
|
|
|
|
Morais LAS (2009). Influência dos fatores abióticos na composição química dos óleos essenciais. Hort. Brás. 27: 4050-4063.
|
|
|
|
|
Pagter M, Bragato G, Brix H (2005). Tolerance and physiological responses of Phragmites australis to water deficit. Aquat. Bot. 81(4):285-299.
Crossref
|
|
|
|
|
Petropoulos SA, Daferera D, Polissiou MG, Passam HC (2008). The effect of water deficit stress on the growth, yield and composition of essential oils of parsley. Sci. Hortic. 115(4):393-397.
Crossref
|
|
|
|
|
Reyes-Munguía A, Zavala-Cuevas D, Alonso-Martínez A (2012). Perejil (Petroselinum crispum):compuestos químicos y aplicaciones. Tlatemoani: Revista Académica de Investigación. 11:18.
|
|
|
|
|
Stanković M, Nikolić N, Stanojević L, Petrović S, Cakić M (2005). Hydrodistillation kinetics and essential oil composition from fermented parsley seeds. CI&CEQ.11:25-29.
Crossref
|
|
|
|
|
Tardieu F, Bruckler L, Lafolie F (1992). Root clumping may affect the root water potential and the resistence to soil-root water transport. Plant Soil. 140(2):291-301.
Crossref
|
|
|
|
|
Tzakou O, Pitarokili D, Chinou IB, Harvala C (2001). Composition and antimicrobial activity of the essential oil of Salvia ringens. Planta Med. 67(1):81-83.
Crossref
|
|
|
|
|
Vora SR, Patil RB, Pillai MM (2009). Protective effects of Petroselinum crispum (Mill) Nyman ex A. W. Hill leaf extract on D-galactose-induced oxidative stress in mouse brain. Indian J. Exp. Biol. 47:338-342.
|
|
|
|
|
Zehtab-salmasi S, Javanshir A, Omidbaigi R, Alyari H, Ghassemi-golezani K (2001). Efects of water supply andsowing date on performance and essential oil production of anise (Pimpinella anisum L.). Acta Agron. Hung. 49(1):75-81.
Crossref
|
|