Full Length Research Paper

ABSTRACT
Species from Aphanomyces genus were investigated in 225 limnologically and trophically different water bodies (springs, rivers, ponds and lakes) as well as in humid soils of north- eastern Poland. Distribution of particular species and their role in diverse ecosystems has been investigated. Thirty taxa, including: 2 species of cultivated parasitic plants, 3 species of parasitic animals, 6 saprotrophic species and 5 species of saprotrophic/ opportunistic species were recorded. Some of Aphanomyces species occurred also as parasites of algae, straminipiles and invertebrates. Among parasitic species: A. cochlioides, A. euteiches (cultivated plant parasite), A. astaci (crayfish parasite), A. piscicida (fish parasite) playing economically important role were found. Amino acid, carbohydrate and urease assimilation tests were used.
Key words: Aphanomyces species, ecological diversity, hydrochemistry, parasite, economical importance.
INTRODUCTION
MATERIALS AND METHODS
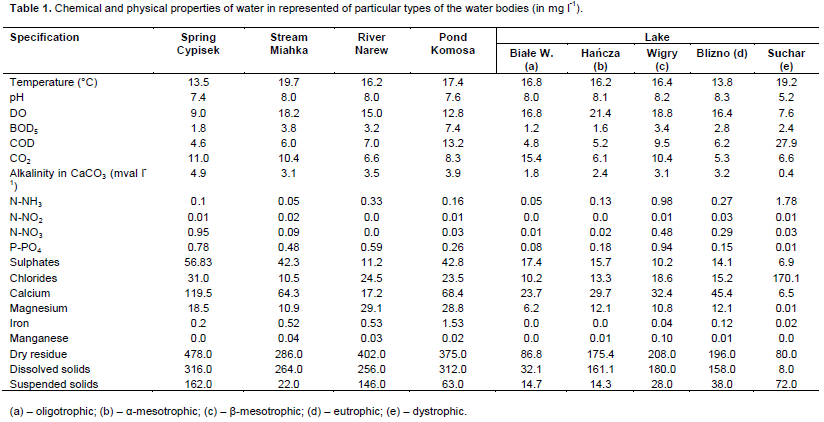
RESULTS
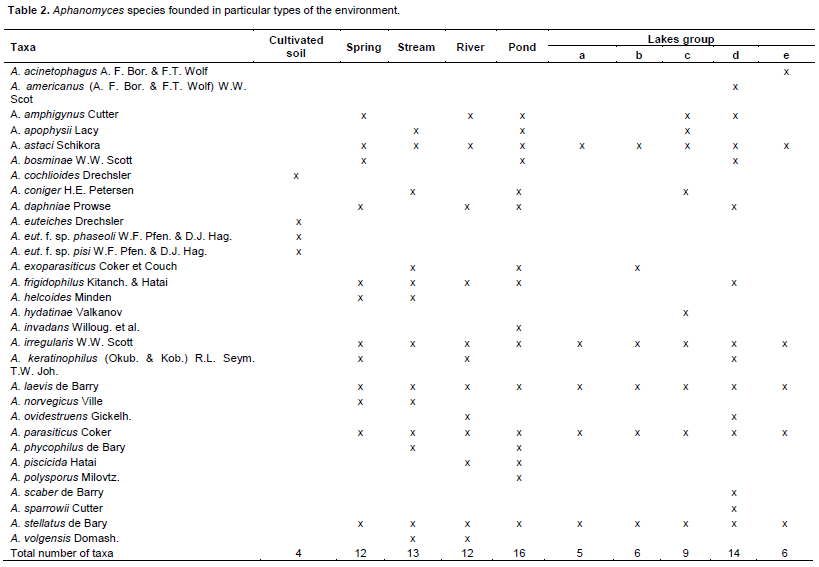
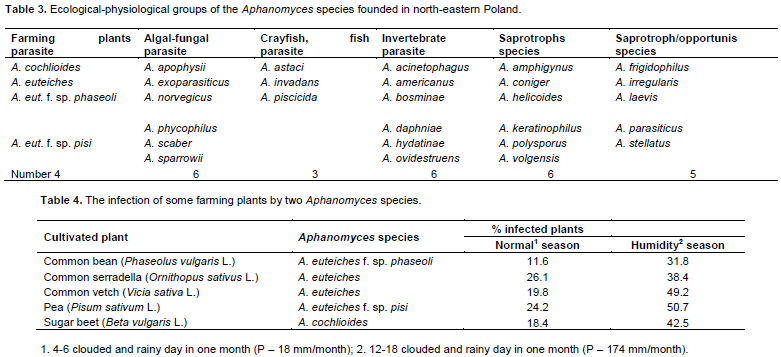

DISCUSSION
CONCLUSIONS
CONFLICT OF INTEREST
REFERENCES
|
Ali EW (2009). Antifungal activity of sodium chloride on Saprolegnia diclina and Aphanomyces sp. Acta Mycol. 44:125-138. |
|
|
APHA (American Public Health Association) (2005). Standard methods for the examination of water and wastewater, APHA, Washington, DC, USA. |
|
|
Ballesteros I, Martin MP, Dieguez-Uribeondo J (2006). First isolation of Aphanomyces frigidophilus (Saprolegniales) in Europe. Mycotaxon 95:335-340. |
|
|
Bangsund DA, Leistritz FL (1993). Economic contribution of the sugarbeet industry to the economy of North Dakota and Minnesota. 1998 Sugarbeet Res. Ext. Rept. 29:160-179. |
|
|
de Bary HA (1860). Einige neue Saprolegnieen. Jahrb. Wiss. Bot. 2:169-192. |
|
|
Batko A (1975). Hydromycology – an overview. PWN, Warszawa. |
|
|
Brantner JR, Windels CE (2001). Variability of spore production and aggressiveness of Aphanomyces cochlioides on sugarbeet 2000. Sugarbeet Res. Ext. Rept. 31:241-246. |
|
|
Bruno DW, Wood BP (1999). Saprolegnia and other Oomycetes. In: Woo PTK & Bruno DW (Eds.), Fish Diseases and Disorders. Viral, Bacterial and Fungal Infections, vol. 3. CABI Publishing, Wallingford, Oxon, UK, pp. 599-659. |
|
|
Cerenius L, Söderhäll K (1985). Repeated zoospore emergence as a possible adaptation to parasitism in Aphanomyces. Exper. Mycol. 9:259-263. |
|
|
Chien C–Y (1981). Observations on the growth and morphology of saprolegniaceous fungi isolated from rainbow trout (Salmo gairdneri). Fish Pathol. 15:241-247. |
|
|
Chinabut S (1998). Epizootic alcerative syndrome: information up to 1997. Fish Pathol. 33:321-326. |
|
|
Czeczuga B (1997). Aquatic fungi growing on lamprey eggs (Petromyzontidae). Bull. Lampetra 3:7-19. |
|
|
Czeczuga B, Bartel R, Semeniuk A, Czeczuga–Semeniuk E, MuszyÅ„ska E, Godlewska A, Mazalska B, Grochowski A (2011a). Straminipilous organisms (Mycota) growing on the eggs of Atlantic salmon (Salmo salar L.) entering Polish rivers for spawning or reared in fresh water. Trends Comp. Biochem. Physiol. 15:73-81. |
|
|
Czeczuga B, Czeczuga-Semeniuk E, Semeniuk A (2011b). Aquatic fungi developing on eggs of Chinook salmon Oncorhynchus tshawytscha and same of their biochemical characteristics. Trends Comp. Biochem. Physiol. 16:85-92. |
|
|
Czeczuga B, Czeczuga-Semeniuk E, Semeniuk A, Semeniuk J (2013). Straminipiles (Oomycota) developing on the eggs of an African catfish, Clarias gariepinus Burchell in water bodies of Poland. Afr. J. Microbiol. Res. 7(20):2378-2384. |
|
|
Czeczuga B, Kiziewicz B, Muszyńska E (2004). Presence of zoosporic fungus species on the eggs of whitefish from Lake Gołdapiwo, Mazury Region. Med. Weteryn. 60:379-383. |
|
|
Czeczuga B, Muszyńska E (1997). Aquatic fungi growing on the eggs of Polish cobitid fish species. Acta Hydrobiol. 39:67-75. |
|
|
Czeczuga B, Muszyńska E (1999). Aquatic fungi growing on the eggs of fishes representing 33 cyprinid taxa (Cyprinidae) in laboratory conditions. Acta Ichthyol. Piscat. 29:53-72. |
|
|
David JC, Kirk PM (1997). Index of Fungi. Elsevier Publication 6:706. |
|
|
Dick MW (2001). Straminipilous Fungi: Systematics of the Peronosporomycetes including accounts of the marine straminipilous protists the plasmodiophorids and similar organisms. Kluwer Academic Publishers, Dordrecht. |
|
|
Dieguez-Uribeondo J, Garcia MA, Cerenius L, Kozubikova E, Ballesteros I, Windels C, Weiland J, Kator H, Söderhall K, Martin MP (2009). Phylogenetic relationships among plant and animal parasites, and saprotrophs in Aphanomyces (Oomycetes). Fung. Gen. Biol. 46:365-376. |
|
|
Drechsler C (1929). The beet water mold and several related root parasites. J. Agric. Res. 38:335. |
|
|
Dyer AT, Szabo LJ, Windels CE (2004). Characterization and spatial distribution of Aphanomyces in sugarbeet fields. J. Sugar Beet Res. 41:1-16. |
|
|
Egusa S, Masuda N (1971). A new fungal disease of Plecoglassus altivelis. Fish Pathol. 6:41-46. |
|
|
Dudka IA, Isaeva NM, Davydova ON (1989). Saprolegniaceae breeding fish mycoses. Mycol. Phytopatol. 23:488-498. |
|
|
El-Hissy FT, Ali EH, Abdel-Raheem A (2004). Diversity of zoospore of fungi of recovered from the surface of water bodies in four Egyptian lakes. Ecohydrol. Hydrobiol. 4:77-84. |
|
|
El-Sharouny HM, Badram RAM (1995). Experimental transmission and pathogenicity of some zoosporic fungi to Tilapia fish. Mycopathology 132:95-105. |
|
|
FAO (2012). Aquatic Sciences and Fisheries Information System (ASFSZ) Species List, Rome. |
|
|
Fuller MS, Jaworski A (1987). Zoosporic Fungi in Teaching and Research. Southeastern Publishing, Athens. |
|
|
Golterman HL, Clymo RS (1969). Methods for chemical analysis of fresh waters. IBP, Handbook No 8, Blackwell Scientific Publications, Oxford. |
|
|
Hatai K (1980). Studies on pathogenic agents of saprolegniasis in fresh water fishes. Spec. Rep. Nagasaki Prefect. Inst. Fish 8:1-95. |
|
|
Hatai K, Hoshiai G (1992). Mass mortality in cultured coho salmon (Oncorhynchus kisutch) due to Saprolegnia parasitica Coker. J. Wild Dis. 28:532-536. |
|
|
Hoagland KD, Rosowsky JR, Gretz MR, Roemet SC (1993). Diatom extracellular polymeric substances: function, fine structure, chemistry and physiology. J. Phycol. 29:537-566. |
|
|
Holub EB, Grau CR, Parke JL (1991). Evaluation of the forma specialis concept in Aphanomyces euteiches. Mycol. Res. 95:147-157. |
|
|
Johnson TW, Seymour RL, Padgett DE (2002). Biology and systematics of the Saprolegniaceae. |
|
|
Johnson RA, Zabrecky J, Kiryu Y, Shields JD (2004). Infection experiments with Aphanomyces invadans in four species of estuarine fish. J. Fish Dis. 27:287-295. |
|
|
Kitancharoen N, Hatai K (1997). Aphanomyces frigidophilus sp. nov. from eggs of Japan char, Salvelinus leucomaenis. Mycoscience 38:135-140. |
|
|
Kitancharoen N, Hatai K (1998). Some biochemical characteristics of fungi isolated from salmonid eggs. Mycoscience 39:249-255. |
|
|
Kiziewicz B, Dieguez-Uribeondo J, Martin MP (2013). Aphanomyces frigidophilus, fungus – like organisms isolated from water of springs in BiaÅ‚ystok, Poland. Afr. J. Biotechnol. 12(44):6310-6314. |
|
|
Lartzeva LV (1986). Saprolegnia on the spawn of sturgeons and salmon. Hydrobiol. J. 22:103-107. |
|
|
Levenfors JP, Fatehi J (2004). Molecular characterization of Aphanomyces species associated with legumes. Mycol. Res. 108:682-689. |
|
|
Lilley JH, Hart D, Panyavachira V, Kanchanak S, Chinabut S, Söderhäll K, Cerenius L (2003). Molecular characterization of the fish – pathogenic fungus Aphanomyces invadans. J. Fish Dis. 26:263-275. |
|
|
OIE (World Organisation for Animal Health) (2007). World animal health information database (WAHID) interface OIE, Paris. |
|
|
Papavizas GC, Ayers A (1974). Aphanomyces species and their root diseases in pea and sugar beet. US Dept. Agric. Technol. Bull. P 1485. |
|
|
Parke JL, Grau CR (1992). Aphanomyces. In: Singleton LL, Mihail JD, Rush CM (Eds.) Methods for Research on Soilborne Phytopathogenic Fungi. APS Pr St. Paul, MN, pp. 27-30. |
|
|
Paterson RRM, Bridge PD (1994). Biochemical Techniques for Filamentous Fungi. CAB International, UK. |
|
|
Patwardhan A, Gandhe R, Ghule V, Mourya D (2005). Larvicidal activity of the fungus Aphanomyces (Oomycetes: Saprolegniales) agains Culex quinqefasciatus. J. Commun. Dis. 37:69-274. |
|
|
Petrini LE, Petrini O (2013). Identyfying Moulds. A Practical Guide. J. Cramer, Stuttgart. |
|
|
Phadee P, Kurata O, Hatai K (2004a). A PCR method for the detection of Aphanomyces piscicida. Fish Pathol. 39:5-32. |
|
|
Phadee P, Kurata O, Hatai K, Irono I, Aoki T (2004b). Detection and identification of the fish phatogenic Aphanomyces piscicida using polymerase chain reaction (PCR) with species – specific primers. J. Aquat. Anim. Health 16:220-230. |
|
|
Prowse GA (1954). Aphanomyces daphniae sp. nov., parasitic on Daphnia hyalina. Trans. Br. Mycol. Soc. 37:22-28. |
|
|
Pystina KA (1994). Ordines Saprolegniales, Leptomitales, Lagenidiales. Nauka, Sankt Petersburg. |
|
|
Roberts RJ, Willoughby LG, Chinabut S (1993). Mycotic aspects of epizootic ulcerative syndrome (EUS) of Asian fishes. J. Fish Dis. 16:169-183. |
|
|
Royo F, Andersson G, Bangyeekhun E, Muzquiz JL, Söderhäll K, Cerenius L (2004). Physiological and genetic characterization of some new Aphanomyces strains isolated from freshwater crayfish. Vet. Microbiol. 104:103-112. |
|
|
Sati SC, Khulbe RD (1981). A new host record for the fungal genus Achlya. Curr. Sci. (India) 50(16):313. |
|
|
Sauvage H, Moussarat A, Boist F, Tivioli B, Barray S, Laal K (2007). Development of a molecular method to detect and quantyfi Aphanomyces euteiches in soil. Ferms Microbiol. Lett. 273:64-69. |
|
|
Schikora F (1903). Über die Krebpest und ihren Erreger. Fischereiztg. (neudamm) 6:353-355. |
|
|
Scott WW (1961). A monograph of the genus Aphanomyces. Wirginia Agric. Exp. Station Technol. Bull. 151:1-95. |
|
|
Seymour RL, Fuller MS (1987). Colletion and isolation of water molds (Saprolegniaceae) from water and soil. In: Fuller M.S., Jaworski A. (Eds.). Zoosporic Fungi in Teaching and Research. Southeastern Publishing, Athens, pp. 125-127. |
|
|
Shaheen AA, Elsayed E, Faisal M (1999). Isolation Aphanomyces sp(p) associated with skin lesions and mortalities in striped (Mugil cephalus) and the thin lip (Liza ramada) grey mullets. Bull. Eur. ass. Fish Pathol. 19:79-82. |
|
|
Söderhäll K, Cerenius L (1999). The crayfish plaque fungus: history and recent advances. Fresh. Crayf. 12:11-35. |
|
|
Watanabe T (2002). Pictorial Atlas of Soil and Seed Fungi: Morphologies of Cultured Fungi and Key Species. CRC Press Boca Raton, Florida. |
|
|
Willoughby LG, Roberts RJ (1994). Improved methodology for isolation of the Aphanomyces fungal pathogen of epizootic ulcerative (EUS) in Asian fishes. J. Fish Dis. 17:541-543. |
|
|
Willoughby LG, Roberts RJ, Chinabut S (1995). Aphanomyces invaderis sp. nov., the fungal pathogen of freshwater tropical fish affected by epizootic ulcerative syndrome. J. Fish Dis. 18:273-275. |
|
|
Yuasa K, Hatai K (1996). Some biochemical characteristics of the genera Saprolegnia, Achlya and Aphanomyces isolated from fishes with fungal infection. Mycoscience 37:477-479. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


