Full Length Research Paper

ABSTRACT
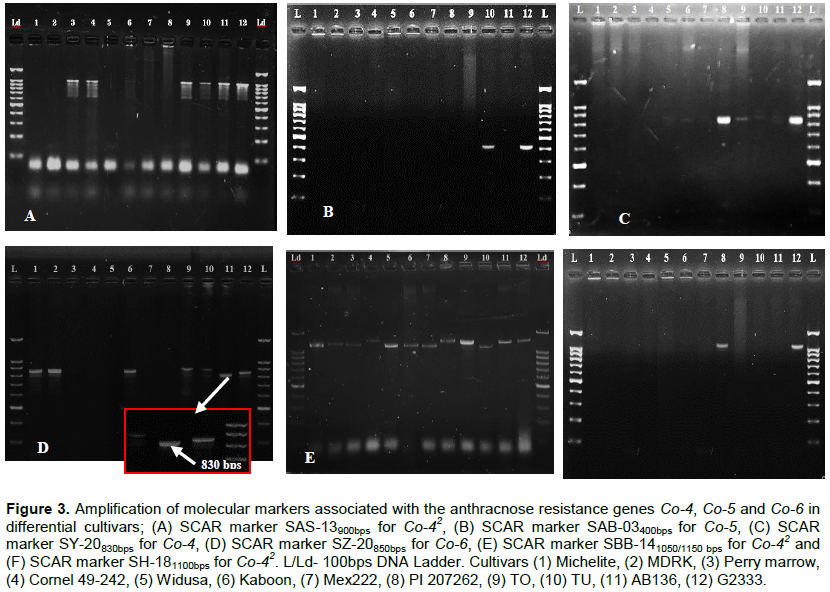
Common bean anthracnose caused by Colletotrichum lindemuthianum is one of the major biotic constraints to production of French beans (Phaseolus vulgaris L.) in Kenya. This study aimed at screening French bean genotypes in relation to their response to common bean anthracnose in order to identify potential sources of germplasm for breeding. The genotypes were tested in three sites (Kakamega, Mwea and Kutus) where results revealed a significant genotype by environment interaction effect (p ≤ 0.05) which emphasizes the strong influence of the growing conditions on the expression of host resistance. Physiological characterization identified a total of 14 distinct races out of 16 successfully plated isolates, revealing a very high diversity of C. lindemuthianum in Kenya. Six races have not been reported in previous studies in the country and are considered as new races, that is, races 84, 141, 246, 515, 576 and 768. Andean race 401 was the most virulent race with a virulence index of 67% among the genotypes. Molecular analysis using six sequence characterized amplified region (SCAR) markers revealed polymorphism among the genotypes. The SCAR markers SBB-14, SH-18 and SAB-03 have shown to be useful for marker assisted selection (MAS) of the target resistance genes. This study has also identified locally improved breeding lines as potential donors for resistance breeding to C. lindemuthianum in Kenya.
Key words: Anthracnose, sequence characterized amplified region (SCAR), marker assisted selection (MAS).
INTRODUCTION
MATERIALS AND METHODS


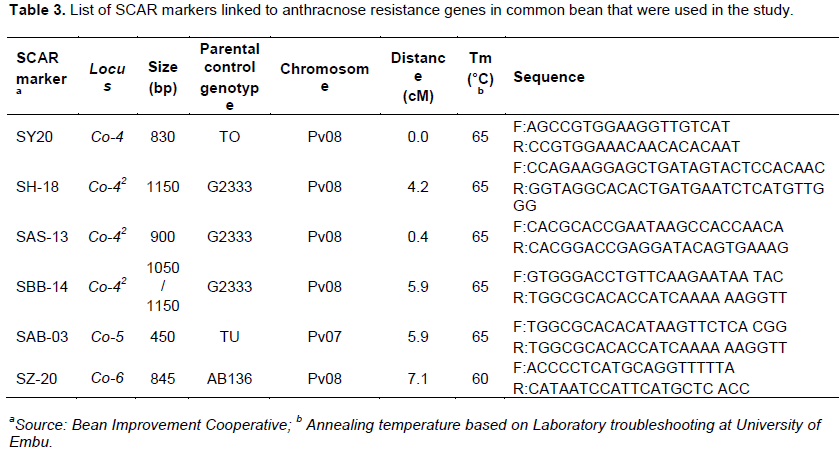
RESULTS AND DISCUSSION
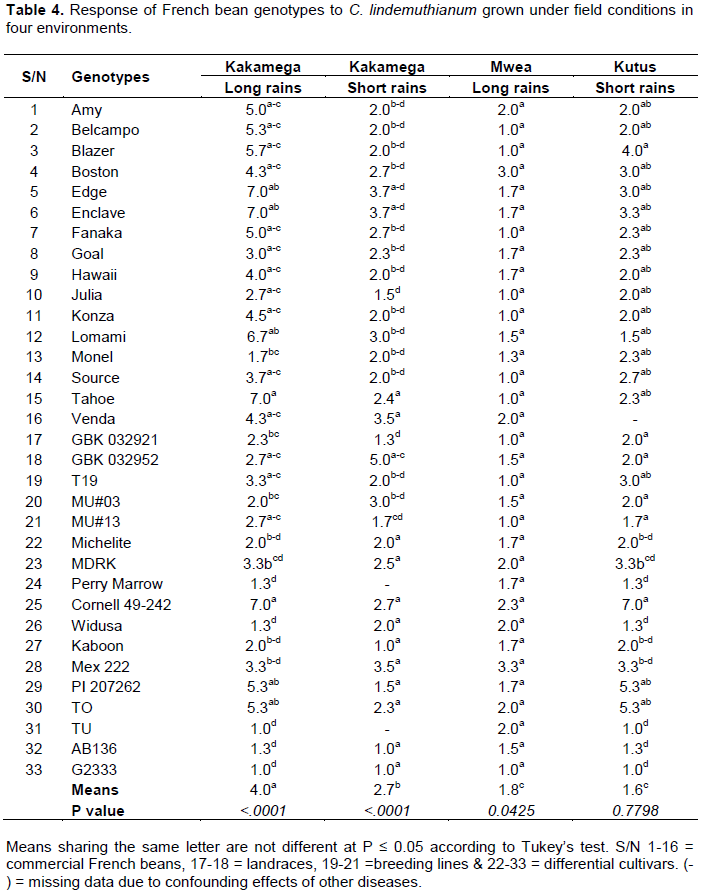
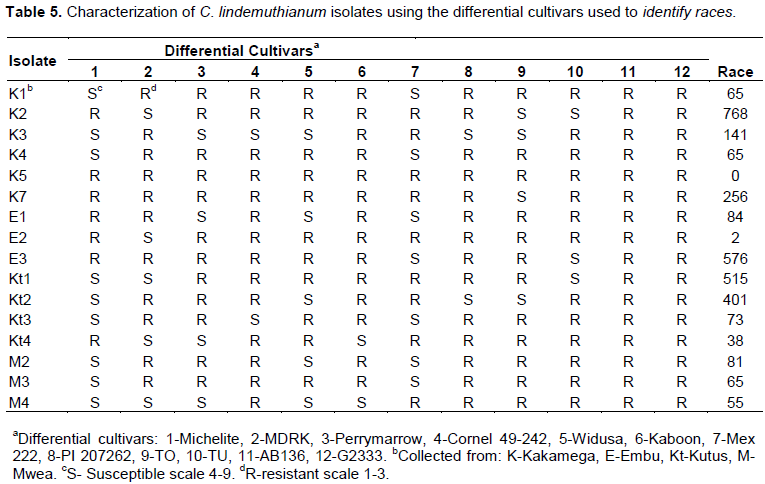
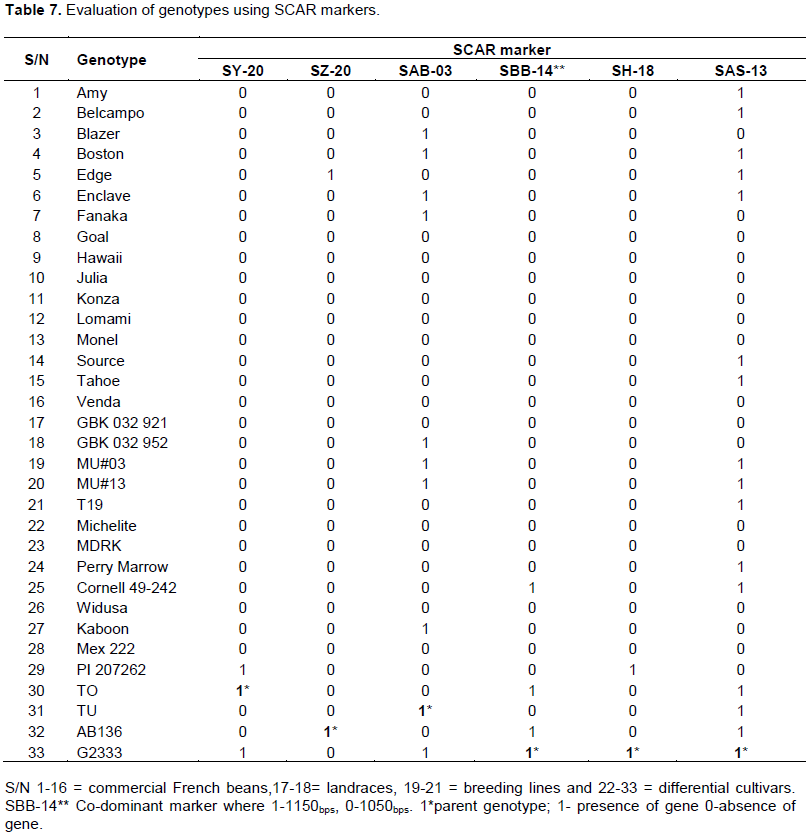
CONCLUSION
ACKNOWLEDGMENT
CONFLICT OF INTERESTS
REFERENCES
|
Allaire H, Brady T (2010). Classification and botanical description of legumes. Retrieved from |
|
|
Allen D, Edje O (1990). Common bean in African farming systems. Paper presented at the Progress in improvement of common bean in Eastern and Southern Africa. Network on Bean Research in Africa Workshop Series. |
|
|
Alzate-Marin AL, Baía GS, De Paula TJ, De Carvalho GA, De Barros EG, Moreira MA (1997). Inheritance of anthracnose resistance in common bean differential cultivar AB 136. Plant Disease and Genetics 81(9):996-998. |
|
|
Alzate-Marin AL, Menarim H, Chagas JM, Barros EGd, Moreira MA (2000). Identification of a RAPD marker linked to the Co-6 anthracnose resistant gene in common bean cultivar AB 136. Genetics and Molecular Biology 23(3):633-637. |
|
|
Aregbesola E, Ortega-Beltran A, Falade T, Jonathan G, Hearne S, Bandyopadhyay R (2020). A detached leaf assay to rapidly screen for resistance of maize to Bipolaris maydis, the causal agent of southern corn leaf blight. European Journal of Plant Pathology 156(1):133-145. |
|
|
Arunga EE, Odikara OS (2020). Characterization of Kenyan French Bean genotypes into gene pool affiliations using allele specific markers. African Journal of Biotechnology 19(9):653-660. |
|
|
Arunga EE, Kinyua M, Ochuodho J, Owuoche J, Chepkoech E (2015). Genetic diversity of determinate French beans grown in Kenya based on morpho-agronomic and simple sequence repeat variation. Plant Breeding and Crop Science 7:240-250. |
|
|
Arunga EE, Ochuodho J, Kinyua M, Owuoche J (2012). Characterization of Uromyces appendiculatus isolates collected from snap bean growing areas in Kenya. African Journal of Agricultural Research 7(42):5685-5691. |
|
|
Asfaw A, Blair MW, Almekinders C (2009). Genetic diversity and population structure of common bean (Phaseolus vulgaris L.) landraces from the East African highlands. Theoretical Applied Genetics 120(1):1-12. |
|
|
Awale HE, Kelly JD (2001). Development of SCAR markers linked to Co-4^ 2 gene in common bean. Annual Report-Bean Improvement Cooperative 44:119-120. |
|
|
Balardin R, Jarosz A, Kelly J (1997). Virulence and molecular diversity in Colletotrichum lindemuthianum from South, Central, and North America. Phytopathology 87(12):1184-1191. |
|
|
Balardin RS, Kelly JD (1998). Interaction between Colletotrichum lindemuthianum races and gene pool diversity in Phaseolus vulgaris. Journal of the American Society for Horticultural Science 123(6):1038-1047. |
|
|
Batureine MJ (2009). Diversity of Colletotrichum lindemuthianum and reaction of common bean germplasm to anthracnose disease. (B.Sc. AGR- Hons: Crop Option), Makerere University, Uganda. |
|
|
Beraldo ALA, Colombo CA, Chiorato AF, Ito MF, Carbonell SM (2009). Use of SCAR markers in common bean breeding for anthracnose resistance. Bragantia 68(1):53-61. |
|
|
Blair MW, Porch T, Cichy K, Galeano CH, Lariguet P, Pankhurst C, Broughton W (2007). Induced mutants in common bean (Phaseolus vulgaris), and their potential use in nutrition quality breeding and gene discovery. Israel Journal of Plant Sciences 55(2):191-200. |
|
|
Broughton WJ, Hernandez G, Blair M, Beebe S, Gepts P, Vanderleyden J (2003). Beans (Phaseolus spp.)-model food legumes. Plant Soil 252(1):55-128. |
|
|
Browne R, Cooke B (2004). Development and evaluation of an in vitro detached leaf assay forc pre-screening resistance to Fusarium head blight in wheat. European Journal of Plant Pathology 110(1):91-102. |
|
|
Burkett-Cadena M, Kokalis-Burelle N, Lawrence K, Van Santen E, Kloepper J (2008). Suppressiveness of root-knot nematodes mediated by rhizobacteria. Biological Control 47(1):55-59. |
|
|
Campa A, Rodríguez-Suárez C, Giraldez R, Ferreira JJ (2014). Genetic analysis of the response to eleven Colletotrichum lindemuthianum races in a RIL population of common bean (Phaseolus vulgaris L.). BMC Plant Biology 14(1):115. |
|
|
Chilagane LA (2017). Genotype-pathogen characterization and markers identification for angular leaf spot disease resistance in common bean in Tanzania. Sokoine University of Agriculture. |
|
|
Coimbraâ€Gonçalves GK, Gonçalvesâ€Vidigal MC, Coelho RT, Valentini G, Vidigal Filho PS, Lacanallo GF, Sousa LL, Elias HT (2016). Characterization and mapping of anthracnose resistance gene in Mesoamerican common bean cultivar Crioulo 159. Crop Science 56(6):2904-2915. |
|
|
Conner RL, Gillard CL, Mcrae KB, Hwang SF, Chen YY, Hou A, Penner WC, Turnbull GD (2019). Survival of the bean anthracnose fungus (Colletotrichum lindemuthianum) on crop debris in Canada. Canadian Journal of Plant Pathology 41(2):209-217. |
|
|
Costa M, Tanure J, Arruda K, Carneiro J, Moreira M, Barros E (2010). Development and characterization of common black bean lines resistant to anthracnose, rust and angular leaf spot in Brazil. Euphytica 176(2):149-156. |
|
|
De Lima CSA, Gonçalves-Vidigal MC, TA, Santana Gilio, Lacanallo GF, Valentini G, Martins Vanusa dSR, Song Q, Galván Marta Z, Hurtado-Gonzales OP, Pastor-Corrales MA (2017). Genetics and mapping of a new anthracnose resistance locus in Andean common bean Paloma. BMC Genomics 18(1):306. |
|
|
Deeksha J, Hooda K, Bhatt J, Mina B, Gupta H (2009). Suppressive effects of composts on soil-borne and foliar diseases of French bean in the field in the western Indian Himalayas. Crop Protection 28(7):608-615. |
|
|
Drijfhout E, Davis J (1989). Selection of a new set of homogeneously reacting bean (Phaseolus vulgaris) differentials to differentiate races of Colletotrichum lindemuthianum. Plant Pathology 38(3):391-396. |
|
|
Fernández MT, Fernandez M, Casares A, Rodriguez R, Fueyo M (2000). Bean germplasm evaluation for anthracnose resistance and characterization of agronomic traits: A new physiological strain of Colletotrichum lindemuthianum infecting Phaseolus vulgaris L. in Spain. Euphytica 114(2):143-149. |
|
|
Ferreira JJ, Campa A, Kelly JD, Varshney R, Tuberosa RJ, Wiley (2013). Organization of genes conferring resistance to anthracnose in common bean. Translational Genomics for Crop Breeding. |
|
|
Garzón LN, Ligarreto GA, Blair MW (2008). Molecular markerâ€assisted backcrossing of anthracnose resistance into andean climbing beans (Phaseolus vulgaris L.). Crop Science 48(2):562-570. |
|
|
Geffroy V, Sévignac M, Billant P, Dron M, Langin T (2008). Resistance to Colletotrichum lindemuthianum in Phaseolus vulgaris: a case study for mapping two independent genes. Theoretical Applied Genetics 116(3):407-415. |
|
|
Gonçalves-Vidigal M, Cardoso AA, Vieira C, Saraiva LS (1997). Inheritance of anthracnose resistance in common bean genotypes PI 207262 and AB 136. Brazilian Journal of Genetics 20. |
|
|
Gonçalves-Vidigal M, Pedro Filho SV, Medeiros A, & Pastor-Corrales M (2009). Common bean landrace Jalo Listras Pretas is the source of a new Andean anthracnose resistance gene. Crop Science 49(1):133-138. |
|
|
Gonçalves-Vidigal MC, Cruz AS, Garcia A, Kami J, Vidigal Filho PS, Sousa LL, McClean P, Gepts P, Pastor-Corrales M (2011). Linkage mapping of the Phg-1 and Co-14 genes for resistance to angular leaf spot and anthracnose in the common bean cultivar and 277. Theoretical Applied Genetics 122(5):893-903. |
|
|
Gonçalves-Vidigal MC, Kelly JD (2006). Inheritance of anthracnose resistance in the common bean cultivar Widusa. Euphytica 151(3):411-419. |
|
|
Gonçalves-Vidigal MC, Sakiyama NS, Vidigal Filho PS, Júnior ATA, Poletine JP, Oliveira VR (2001). Resistance of common bean cultivar AB 136 to races 31 and 69 of Colletotrichum lindemuthianum: the Co-6 locus. Crop Breeding Applied Biotechnology 1(2). |
|
|
González AM, Yuste-Lisbona FJ, Rodiño AP, De Ron AM, Capel C, García-Alcázar M, Lozano R, Santalla M (2015). Uncovering the genetic architecture of Colletotrichum lindemuthianum resistance through QTL mapping and epistatic interaction analysis in common bean. Frontiers in Plant Science 6:141. |
|
|
Kelly J, Gepts P, Miklas P, Coyne D (2003). Tagging and mapping of genes and QTL and molecular marker-assisted selection for traits of economic importance in bean and cowpea. Field Crops Research 82(2-3):135-154. |
|
|
Kelly JD, Vallejo VA (2004). A comprehensive review of the major genes conditioning resistance to anthracnose in common bean. HortScience 39(6):1196-1207. |
|
|
Kimno S, Kiplagat O, Arunga E, Chepkoech E (2016). Evaluation of Selected French Bean (Phaseolus vulgaris L.) Genotypes for Resistance to Angular Leaf Spot (Pseudocercospora griseola) in Western Kenya. American Journal of Experimental Agriculture 13(4):1-6. |
|
|
Kiptoo GJ, Kinyua MG, Kiplagat OK, Matasyoh LG (2020). Incidence and severity of anthracnose (Colletotrichum lindemuthianum) on selected common bean (Phaseolus vulgaris L.) genotypes. Agricultural Research 5(84):1-12. |
|
|
Lacanallo G, Gonçalves-Vidigal M, Vidigal Filho P, Kami J, Gonela A (2010). Mapping of an Andean gene for resistance to anthracnose in the landrace Jalo Listras Pretas. Paper presented at the Annual Report of the Bean Improvement Cooperative. |
|
|
Mahuku G, Jara C, Cajiao C, Beebe (2002). Sources of resistance to Colletotrichum lindemuthianum in the secondary gene pool of Phaseolus vulgaris and in crosses of primary and secondary gene pools. Plant Disease 86(12):1383-1387. |
|
|
Mahuku GS, Riascos JJ (2004). Virulence and molecular diversity within Colletotrichum lindemuthianum isolates from Andean and Mesoamerican bean varieties and regions. European Journal of Plant Pathology 110(3):253-263. |
|
|
Melotto M, Kelly J (2000). An allelic series at the Co-1 locus conditioning resistance to anthracnose in common bean of Andean origin. Euphytica 116(2):143-149. |
|
|
Miklas P, Kelly J, Beebe S, Blair M (2006). Common bean breeding for resistance against biotic and abiotic stresses: from classical to MAS breeding. Euphytica 147(1-2):105-131. |
|
|
Miklas P, Kelly J, Singh S (2003). Registration of anthracnose-resistant pinto bean germplasm line USPT-ANT-1. Crop Science 43(5):1889-1891. |
|
|
Mogita W, Ochuodho J, Gohole L (2011a). Analysis of Western Kenya Isolates of Colletotrichum lindemuthianum Casual Agent of Bean Anthracnose using Pathogenicty and BOX-A1R Analysis. African Journal of Education, Science and Technology 1(3):155-162. |
|
|
Mogita W, Ochuodho J, Gohole L, Arunga E, Billy M (2011b). Incidence of bean anthracnose in Western Kenya and its management using aqueous extract of Aloe vera. African Journal of Education, Science and Technology 3(3):6-12. |
|
|
Mpeguzi M, Susan NM, Mabagala R, Chilagane LA (2020). Races of Colletotrichum lindemuthianum (Sacc. Magnus) Briosi Cavara in major bean growing regions in Tanzania. African Journal of Plant Science 14(8):308-314. |
|
|
Musyimi AS (2014). Marker assisted gamete selection for multiple disease resistence in Andean Bean genotypes and characterization of Colletotrichum lindemuthianum in Kenya. (BSc Agriculture), University of Nairobi, 2014. |
|
|
Mwaninki OK (2017). Determination of pre-harvest interval for carbendazim used for control of fungal diseases in French beans. University of Nairobi. |
|
|
Njuguna SM (2014). Marker Assisted Gamete Selection For Multiple Disease Resistance in Mesoamerican Bean Genotypes and Race Typing of Angular Leaf Spot Pathogen In Kenya. University of Nairobi. |
|
|
Nkalubo S (2006). Study of anthracnose (Colletotrichum lindemuthianum) resistance and its inheritance in Ugandan dry bean germplasm. |
|
|
Oblessuc P, Baroni R, da Silva Pereira G, Alisson Fernando, Carbonell S, Brinez B, Luciano Da Costa ES, Garcia AA, Luis EA, Kelly JD (2014). Quantitative analysis of race-specific resistance to Colletotrichum lindemuthianum in common bean. Molecular Breeding 34(3):1313-1329. |
|
|
Ombiri J, Zinkernagel V, Gathuru E, Achwanya O (2002). First report of race 485 of Colletotrichum lindemuthianum in Kenya and its implication in bean resistance breeding. Gartenbauwissenschaft 67(3):81-85. |
|
|
Otsyula R, Rachier G, Ambitsi N, Juma R, Ndiya C, Buruchara R, Sperling L (2004). The use of informal seed producer groups for diffusing root rot resistant varieties during period of acute stress. Paper presented at the Addressing seed security in disaster response: Linking relief with development. |
|
|
Padder B, Sharma P, Awale H, Kelly J (2017). Colletotrichum lindemuthianum, the causal agent of bean anthracnose. Journal of Plant Pathology 99(2):317-330. |
|
|
Pastor-Corrales M (1991). Standardization of differential varieties and race designation of Colletotrichum lindemuthianum. Phytopathology 81:694. |
|
|
Pastor-Corrales MA, Jara C, Singh SP (1998). Pathogenic variation in, sources of, and breeding for resistance to Phaeoisariopsis griseola causing angular leaf spot in common bean. Euphytica 103(2):161-171. |
|
|
Queiroz V, de Sousa C, Costa M, Sanglad D, Arruda K, de Souza T, Ragagnin V, de Barros E, Moreira M (2004). Development of SCAR markers linked to common bean anthracnose resistance genes Co-4 and Co-6 (0084-7747). |
|
|
Ragagnin VA, Sanglard DA, De Souza T, Moreira MA, De Barros EG (2003). Simultaneous transfer of resistance genes for Rust, Anthracnose and Angular Leaf Spot to cultivar Perola assited by molecular markers. Paper presented at the Annual Report-Bean Improvement Cooperative. |
|
|
Rezene Y, Tesfaye K, Mukankusi C, Arunga E, Gepts P (2018). Simple and rapid detached leaf technique for screening common beans (Phaseolus vulgarise L.) in vitro against angular leaf spot (Pseudocercospora griseola) disease. African Journal of Biotechnology 17(35):1076-1081. |
|
|
Schwartz HF, Steadman JR, Hall R, Forster RL (2005). Compendium of bean diseases: American Phytopathological Society (APS Press). |
|
|
Sharma P, Sharma O, Padder B, Kapil R (2008). Yield loss assessment in common bean due to anthracnose (Colletotrichum |
|
|
lindemuthianum) under sub temperate conditions of North-Western Himalayas. Indian Phytopathology 61(3):323. |
|
|
Silvério L, Gonçalves-Vidigal M, Filho P, Barelli MA, Thomazella C, Nunes WM (2002). Genetic resistance to Collectotrichum lindemuthianum race 2047 in G2333. Paper presented at the Annual Report-Bean Improvement Cooperative. |
|
|
Singh A, Singh AP, Ramaswamy HS (2015). Effect of processing conditions on quality of green beans subjected to reciprocating agitation thermal processing. Food Research International 78:424-432. |
|
|
Singh SP, Schwartz HF (2010). Breeding common bean for resistance to diseases: a review. Crop Science 50(6):2199-2223. |
|
|
Sousa LL, Gonçalves AO, Gonçalves-Vidigal MC, Lacanallo GF, Fernandez AC, Awale H, Kelly JD (2015). Genetic characterization and mapping of anthracnose resistance of common bean landrace cultivar Corinthiano. Crop Science 55(5):1900-1910. |
|
|
Sperling L (2001). The effect of the civil war on Rwanda's bean seed systems and unusual bean diversity. Biodiversity Conservation 10(6):989-1010. |
|
|
Tu J (1986). A detached leaf technique for screening beans (Phaseolus vulgaris L.) in vitro against anthracnose (Colletotrichum lindemuthianum). Canadian Journal of Plant Science 66(3):805-809. |
|
|
Twizeyimana M, Ojiambo P, Tenkouano A, Ikotun T, Bandyopadhyay R (2007). Rapid screening of Musa species for resistance to black leaf streak using in vitro plantlets in tubes and detached leaves. Plant Disease 91(3):308-314. |
|
|
Valentini G, Gonçalves-Vidigal MC, Hurtado-Gonzales OP, de Lima Castro SA, Cregan PB, Song Q, Pastor-Corrales MA (2017). High-resolution mapping reveals linkage between genes in common bean cultivar Ouro Negro conferring resistance to the rust, anthracnose, and angular leaf spot diseases. Theoretical Applied Genetics 130(8):1705-1722. |
|
|
Vallejo V, Kelly JD (2001). Development of a SCAR marker linked to Co-5 locus in common bean. Paper presented at the Annual Report-bean Improvement Cooperative. |
|
|
Vallejo V, Kelly JD (2009). New insights into the anthracnose resistance of common bean landrace G 2333. Open Horticulture Journal 2(1):29-33. |
|
|
Vaz Bisneta M, Gonçalvesâ€Vidigal MC (2020). Integration of anthracnose resistance loci and RLK and NBSâ€LRRâ€encoding genes in the Phaseolus vulgaris L. genome. Crop Science. |
|
|
Vieira A, Almeida L, Rodrigues L, Costa J, Melo L, Pereira H, Sanglard D, Souza T (2018). Selection of resistance sources to common bean anthracnose by field phenotyping and DNA marker-assisted screening. Embrapa Arroz e Feijão-Artigo em periódico indexado. |
|
|
Wahome S, Kimani P, Muthomi J, Narla R, Buruchara R (2011). Multiple disease resistance in snap bean genotypes in Kenya. African Crop Science Journal 19(4):289-302. |
|
|
Wortmann CS (1998). Atlas of common bean (Phaseolus vulgaris L.) production in Africa: CIAT. |
|
|
Young R, Melotto M, Nodari R, Kelly J (1998). Marker-assisted dissection of the oligogenic anthracnose resistance in the common bean cultivar,'G2333'. Theoretical Applied Genetics 96(1):87-94. |
|
|
Young RA, Kelly JD (1997). RAPD markers linked to three major anthracnose resistance genes in common bean. Crop Science 37(3):940-946. |
|
|
Zuiderveen GH, Padder BA, Kamfwa K, Song Q, Kelly JD (2016). Genome-wide association study of anthracnose resistance in Andean beans (Phaseolus vulgaris). PLoS ONE 11(6):e0156391. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


