Full Length Research Paper

ABSTRACT
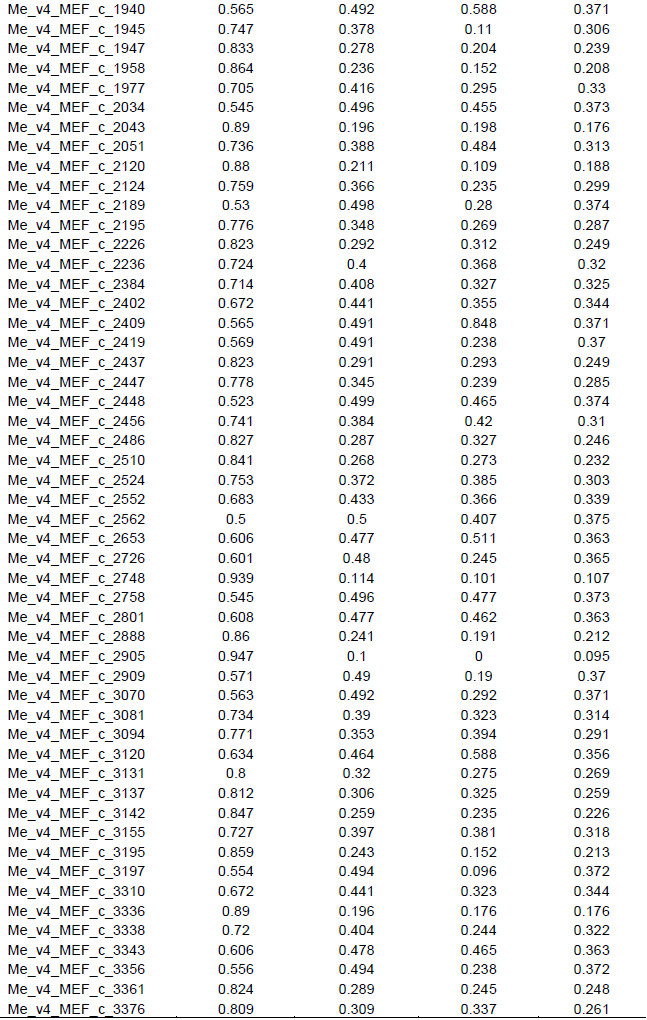
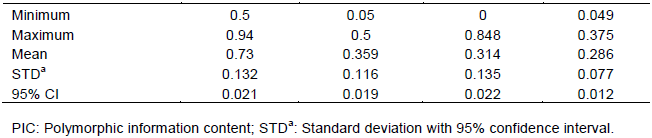
This study seeks to determine the genetic diversity among cassava landraces using single nucleotide polymorphic (SNP) markers. One hundred and five cassava landraces were assayed with 195 SNP markers. Major allele frequency varied from 0.500 to 0.942 with an average of 0.728. Average gene diversity, heterozygosity and polymorphic information content (PIC) were 0.359, 0.314 and 0.286 respectively. These values were generally high considering the bi-allelic nature of SNPs, hence the cassava landraces studied showed moderate to high genetic diversity. This suggests availability of unique and useful alleles that could be exploited for breeding purposes. Inclusion of these landraces in our crop improvement activities will enhance the development of farmer preferred cassava varieties. SNP markers used for the study were highly informative, polymorphic and revealed good estimates of genetic diversity among the landraces. Higher level of genetic variation was observed within population based on analysis of molecular variance (AMOVA). Principal component analysis (PCA) and cluster analysis also grouped landraces into three distinct clusters; however, they did not group in accordance to geographical origin. This could be due to high frequency of germplasm exchange between farmers and subsequent change of the name of the same cultivar. Results from this study may contribute significantly to cassava breeding and germplasm conservation programs.
Key words: Genetic diversity, single nucleotide polymorphisms (SNPs), polymorphic information content (PIC), polymorphic, alleles, heterozygosity, germplasm.
INTRODUCTION
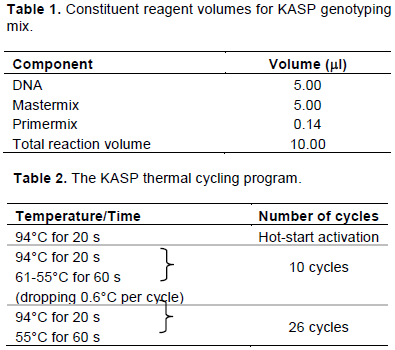
MATERIALS AND METHODS
RESULTS AND DISCUSSION
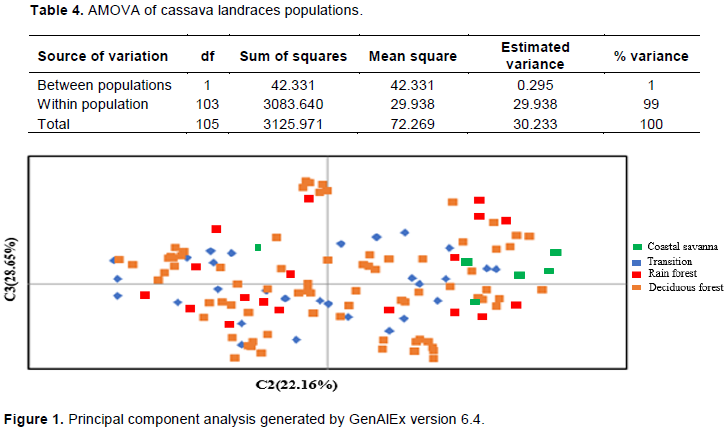

CONCLUSION
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
REFERENCES
|
Adjebeng-Danquah J, Manu-Aduening J, Asante IK, Agyare RY, Gracen V, Offei SK (2020). Genetic diversity and population structure analysis of Ghanaian and exotic cassava accessions using simple sequence repeat (SSR) markers. Heliyon 6(1):e03154. |
|
|
Benesi IRM, Labuschagne MT, Herselman L, Mahungu N (2010). Ethnobotany, morphology and genotyping of cassava germplasm from Malawi. Journal of Biological Sciences 10:616-623. |
|
|
Bostein D, White RL (1980). Construction of a genetic map in man using restriction fragment polymorphism. American Journal of Human Genetics 32(3):314-331. |
|
|
Ching A, Caldwell KS, Jung M, Dolan M, Smith OS, Tingey S, Morgante M, Rafalski AJ (2002). SNP frequency, haplotype structure and linkage disequilibrium in elite maize inbred lines. BioMed Central Genetics 3:19. |
|
|
de Albuquerque HYG, Carmo CD, Brito AC, de Oliveira EJ (2018). Genetic diversity of Manihot esculenta Crantz germplasm based on single nucleotide polymorphism markers. Annals of Applied Biology 2018:1-14. |
|
|
Egnin M, Mora A, Prakash CS (1998). Factors enhancing Agrobacterium tumefaciens mediated gene transfer in peanut (Arachis Hypoges L.). In vitro cellular and Developmental Biology Plants 34:310-318. |
|
|
Emmanuelli F, Lorenzi S, Grzeskowiak L, Catalano V, Stefanini M, Troggio M, Myles S, Martinez-Zapater JM, Zyprian E, Moreira FM, Grando MS (2013). Genetic diversity and population structure assessed by SSR and SNP markers in a large germplasm collection of grape. BMC Plant Biology 2013:13:39. |
|
|
Ferguson ME, Shah T, Kulakow P, Ceballos H (2019). A global overview of cassava genetic diversity. PLoS ONE 14(11):e0224763. |
|
|
Ferguson ME, Hearne SJ, Close TJ, Wanamaker S, Moskal WA, Town CD, de Young J, Marri PR, Rabbi IY, de Villiers EP (2012). Identification, validation and high-throughput genotyping of transcribed gene SNPs in cassava. Theoretical Applied Genetics 124(4):685-695. |
|
|
Filippi CV, Aguirre N, Rivas JG, Zubrzycki J, Puebla A, Cordes D, Moreno MV, Fusari CM, Alvarez D, Heinz RA (2015). Population structure and genetic diversity characterization of a sunflower association mapping population using SSR and SNP markers. BMC Plant Biology 15:52. |
|
|
Gonçalves TM, Vidigal F, Vidigal PS, Ferreira MCG, Rocha RCU, Ortiz VPC, Kvitschal MV (2017). Genetic diversity and population structure of traditional sweet cassava accessions from Southern of Minas Gerais State, Brazil, using microsatellite markers. African Journal of Biotechnology 16:346-358. |
|
|
Kamanda I, Blay ET, Asante IK, Danquah A, Ifie BE, Parkes E, Kulakow P, Rabbi I, Conteh A, Kamara JS, Mensah HK, Whyte JBA, Sesay S (2020). Genetic diversity of provitamin-A cassava (Manihot esculenta Crantz) in Sierra Leone. Genetic Resources and Crop Evolution 67:1193-1208. |
|
|
Karim KY, Ifie B, Dzidzienyo D, Danquah EY, Blay ET, Whyte JBA, Kulakow P, Rabbi I, Parkes E, Omoigui L, Norman PE, Iluebbey P (2020). Genetic characterization of cassava (Manihot esculenta Crantz) genotypes using agro-morphological and single nucleotide polymorphism markers. Journal of Physiology and Molecular Biology of Plants 26(2):317-330. |
|
|
Kawuki RS, Ferguson M, Labuschagne M, Herselman L, Kim DJ (2009). Identification, characterization and application of single nucleotide polymorphisms for diversity assessment in cassava (Manihot esculenta Crantz). Molecular Breeding 9:9264-9280. |
|
|
Liu K, Muse SV (2005). PowerMarker: Integrated analysis environment for genetic marker data. Bioinformatics 21(9):2128-2129. |
|
|
Lopez C, Piegu B, Cooke R, Delseny M, Tohme J, Verdier V (2005). Using cDNA and genomic sequences as tools to develop SNP strategies in cassava (Manihot esculent Crantz). Theoretical and Applied Genetics 110:425-431. |
|
|
Mtunguja MK, Ranjan A, Laswai HS, Muzanila Y, Ndunguru J, Sinha NR (2017). Genetic diversity of farmer-preferred cassava landraces in Tanzania based on morphological descriptors and single nucleotide polymorphisms. Plant Genetic Resources 15:138-146. |
|
|
Mtunguja MK, Laswai HS, Muzanila YC Ndunguru J (2014). Farmer's knowledge on selection and conservation of cassava (Manihot esculenta) genetic resources in Tanzania. Journal of Biology Agriculture and Healthcare 4:120-129. |
|
|
Nei M (1973). Analysis of gene diversity in subdivided populations. Proceedings of the National Academy of Sciences USA 70:3321-3323. |
|
|
Oliveira EJ, De Ferreira CF, Santos VS, Jesus ON, Oliveira GAF, Silva MS Da. (2014). Potential of SNP markers for the characterization of Brazilian cassava germplasm. Theoretical and Applied Genetics 127:1423-1440. |
|
|
Parkinson J, Blaxter M (2009). Expressed sequence tags: An overview. Methods Molecular Biology 533:1â€12. |
|
|
Peakall R, Smouse PE (2006). GENALEX 6: genetic analysis in excel. Population genetic software for teaching and research. Molecular Ecology Notes 6:288-295. |
|
|
Pedri ECM, Hoogerheide ESS, Tiago AV, Cardoso ES, Pinto JMA, Santos LL, Yamashita OM, Rossi AAB (2019). Genetic diversity of cassava landraces cultivated in northern Mato Grosso State, Brazil, using microsatellite markers. Genetics and Molecular Research 18(3):gmr18315. |
|
|
Rabbi I, Hamblin M, Gedil M, Kulakow P, Ferguson M, Ikpan AS (2014). Genetic mapping using genotyping-by-sequencing in the clonally propagated cassava. Crop Science 54(4):1384-1396. |
|
|
Rabbi IY, Kulembeka HP, Masumba E, Marri PR, Ferguson M (2012). An EST-derived SNP and SSR genetic linkage map of cassava (Manihot esculenta Crantz). Theoretical and Applied Genetics 125:329-342. |
|
|
Ren J, Sun D, Chen L, You FM, Wang J, Peng Y, Nevo E, Sun D, Luo MC, Peng J (2013). Genetic diversity revealed by single nucleotide polymorphism markers in a worldwide germplasm collection of durum wheat. International Journal of Molecular Sciences 14:7061-7088. |
|
|
Rimoldi F, Vidigal-Filho PS, Kvitschal MV, Goncalves‑Vidigal, MC, Prioli AJ, Prioli SM, Costa TR (2010). Genetic divergence in sweet cassava cultivars using morphological agronomic traits and RAPD molecular markers. Brazilian Archives of Biology and Technology 53:1477-1486. |
|
|
Singh N, Choudhury DR, Singh AK, Kumar S, Srinivasan K, Tyagi RK, Singh NK, Singh R (2013). Comparison of SSR and SNP markers in estimation of genetic diversity and population structure of Indian rice varieties. PLoS ONE 8:e84136. |
|
|
Tamura K, Dudley J, Nei M, Kumar S (2007). MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Molecular Biology and Evolution 24:1596-1599. |
|
|
Tiago AV, Rossi AAB, Tiago PV, Carpejani AA, Silva BM, Hoogerheide ESS, Yamashita OM (2016). Genetic diversity in cassava landraces grown on farms in Alta Floresta-MT, Brazil A.V. Genetics and Molecular Research 15(3):1-10. |
|
|
Tumuhimbise T, Shanahan P, Melis R, Kawuki R (2014). Combining ability analysis of storage root yield and related traits in cassava at the seedling evaluation stage of breeding. Journal of Crop Improvement 28(4):530-546. |
|
|
Turyagyenda LF, Kizito EB, Ferguson ME, Baguma Y, Harvey JW, Gibson P, Wanjala BW, Osiru DSO (2012). Genetic diversity among farmer-preferred cassava landraces in Uganda. African Crop Science Journal 20(1):15-30. |
|
|
Zhu YL, Song QJ, Hyten DL, Tasell CP, Matukumali LK, Grim DR, Hyatt SM, Fickus EW, Young ND, Cregan PB (2003). Single nucleotide polymorphisms in soybean. Genetics 163:1123-1134. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


