ABSTRACT
Recently, a skin ulcerative disease caused by Vibrio mimicus has led to heavy economic losses in catfish, including yellow catfish, southern catfish, and Zhengchuan catfish in China. Currently, there was no effective method of controlling the outbreak of this disease. In this study, the bacterial isolates were obtained from dying channel catfish and identified as V. mimicus, which consist of formalin-inactivated V. mimicus (antigen). After first immunization, four weeks later, fishes were exposed to V. mimicus and the immune response was analyzed: Fish survival, respiratory burst activity of blood leukocytes, serum agglutination titers, and lysozyme activity, every week (during four weeks). Survival was up 90%. Respiratory burst activity of blood leukocytes, serum agglutination titers, and lysozyme activity were determined at 1, 2, 3, and 4 weeks after primary immunization. Immunization of channel catfish protected hosts against V. mimicus infection with a survival percentage of more than 90%. Respiratory burst activity of blood leukocytes was not affected by vaccination. Serum agglutination titer and lysozyme activity were significantly increased after immunization, in comparison with un-vaccinated control fish. The obtained results indicated that vaccination is an effective method to control the outbreaks of V. mimicus through regulation of the humoral immune response.
Key words: Vibrio mimicus, catfish, skin ulcer, vaccine.
Vibrio mimicus, a Gram-negative bacteria similar to Vibrio cholerae, has been identified as a causative agent of human gastroenteritis, which is characterized by watery to dysentery-like diarrhea (Davis et al., 1981; Takahashi et al., 2007). V. mimicus is a natural inhabitant of aquatic environments, including freshwater, brackish water, and saltwater. V. mimicus has been isolated from water samples (Adeleye et al., 2010; Chowdhury et al., 1989), sediments (Adeleye et al., 2010), aquatic plants (Li et al., 2005), snails (Li et al., 2005), oysters (Li et al., 2005), crayfish (Eaves and Ketterer, 1994), turtle eggs (Campos et al., 1996), shrimp (Guardiola-Avila et al., 2016; Thune et al., 1991; Wang et al., 2003; Wong et al., 1995), crabs (Li et al., 2005), fish (Li et al., 2005).
V. mimicus has been reported to be responsible for ascites disease in aquatic animals (Cen et al., 2013).
Recently we and others have reported that V. mimicus is also a pathogenic agent causing skin ulcerative disease in freshwater catfish species, including yellow catfish (Pelteobagrus fulvidraco) (Geng et al., 2014), southern catfish (Silurus soldatovi meridionalis, Chen) (Geng et al., 2014), and Zhengchuan catfish (Silurus soldatovi meridionalis, Chen â™‚× Silurus asotus ♀) (Zhang et al., 2014). The most evident clinical symptoms of this skin ulcerative disease is the presence of regularly-shaped ulcers with clear boundaries. This disease has resulted in more than 70% cumulative mortality of freshwater fish farms, and has led to severely economic losses to aquaculture in south China according to the data from Guangdong Provincial center for disease control and prevention.
Antibiotics are a suitable strategy often used to control V. mimicus infection in aquaculture animals. However, the excessive use of antibiotics has led to the emergence of antibiotic-resistant bacteria (Liu et al. 2015), and to environmental deterioration (Nugroho and Fotedar, 2013). Therefore, other eco-friendly environment methods are needed to prevent outbreaks of this disease.
Recent studies have indicated that dietary supplementation with mannan oligosaccharide, with customized probiotics, or with organic selenium improved the resistance of marron Cherax tenuimanus (Ambas et al., 2013; Nugroho and Fotedar, 2013) to V. mimicus (Sang et al., 2009). In addition, Cen et al. (2013) and Zhang et al. (2014) have produced a vaccine based on outer membrane protein U (OmpU), protecting carp Ctenopharyngodon idella against V. mimicus infection. Despite the heavy economic losses due to catfish infection with V. mimicus in recent years, no prophylactic method has been developed to provide protection against V. mimicus infection, until now.
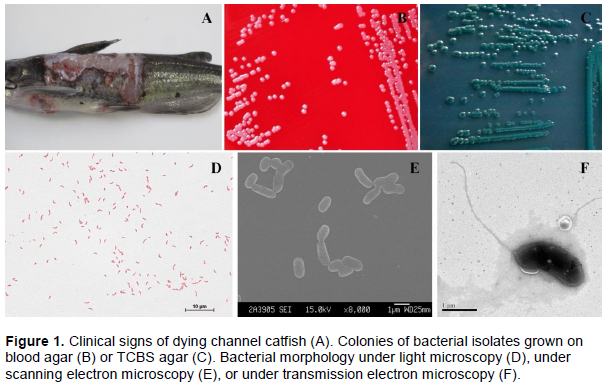
Channel catfish (Ictalurus punctatus Rafinesque) is an economically important fish species, reared in southern China, in places such as Guangdong and Sichuan. In recent years, a disease characterized by skin ulcers (Figure 1A) has been prevalent in farmed channel catfish. This symptom has been observed by Geng et al. (2014) and Zhang et al. (2014) in catfish infected by V. mimicus. Thus the aim of the present study was to isolate V. mimicus bacteria from dying channel catfish, develop an effective prophylaxis method, a formalin-inactivated V. mimicus vaccine. Results of this study will be of immense value to the aquaculture industry in southern China.
Fish
Dying channel catfish were brought to our laboratory for pathogen detection from a channel catfish aquaculture farm (Foshan, Guangdong Province, southern China). Healthy channel catfish were purchased from an aquaculture farm (Guangzhou, Guangdong Province). Fish were acclimatized in tanks for more than 14 days at 28±1°C, and fed daily with a commercial feed. Prior to experiments, five fish were randomly selected to confirm that they had not been infected with bacteria. Using conventional microbiological methods, such as 16S rDNA identification and tissue section, and no signs of bacterial infection were observed in any of the fish samples tested.
Isolation and identification of bacteria
For bacterial isolation, samples from brain, liver, spleen and kidney of the moribund catfish were taken using disposable inoculation loops, and inoculated immediately onto blood agar plates (Huankai, Guangzhou, southern China) or thiosulfate citrate bile salts sucrose(TCBS) agar plates (Huankai). The plates were incubated at 28°C for 48 h. Single colonies from plates were then selected and re-streaked on the same media. The isolates were stored in brain heart infusion (BHI) medium (Huankai) containing 20% (v/v) glycerol in liquid nitrogen.
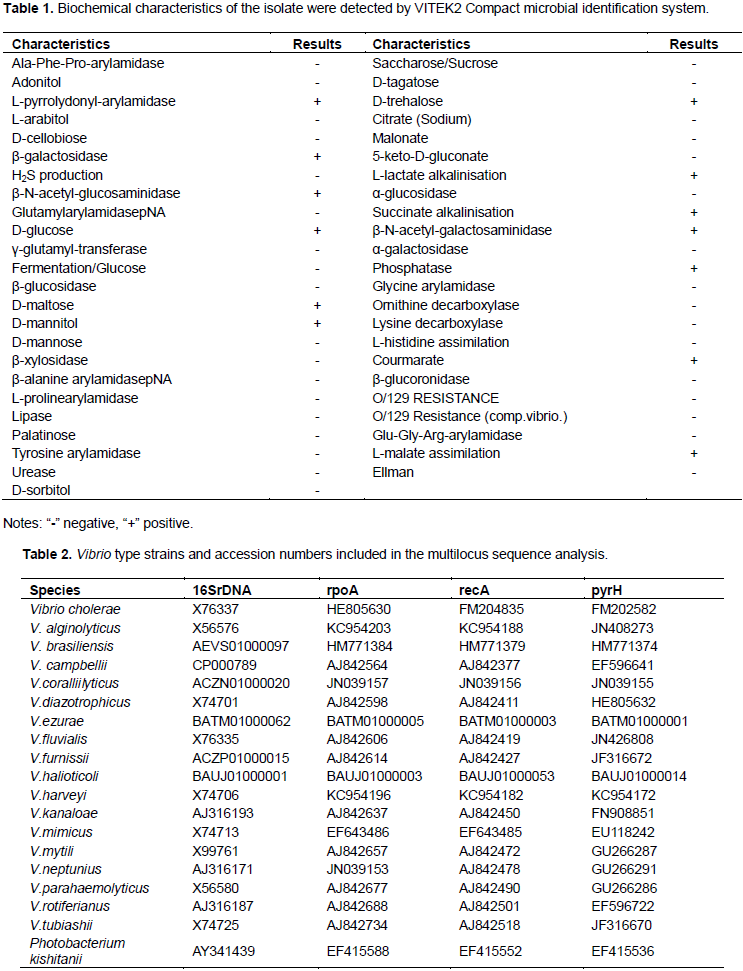
After samples were prepared with the standard methods as described previously (Sun et al., 2009), the morphology of bacteria was observed under light microscopy, scanning electron microscopy and transmission electron microscopy. The biochemical characterization of bacterial isolates was performed using the VITEK2 Compact microbial identification system (BioMerieux, Lyon, France). Additionally, 16S rDNA and three housekeeping genes encoding recombination repair protein (recA), urydilate kinase (pyrH) and RNA polymerase α-chain (rpoA) were selected to identify the bacterial species using the method described by Zhang et al. (2014). Table 2
Immunization and sampling
To prepare inactivated V. mimicus, the NH isolate was cultivated overnight in 5 ml BHI broth at 28°C with shaking at 150 rpm, allowing bacteria to reach the logarithmic growth phase. One milliliter sample of the culture was transferred into 100 ml fresh BHI broth, and cultivated at 28°C with shaking at 150 rpm for 48 h. The culture was then inactivated with 1% formalin at 4 °C for 24 h. The inactivated cells were harvested by centrifugation at 8000×g for 5 min, washed three times with phosphate-buffered saline (PBS) and then adjusted to a suitable concentration with PBS. The suspension of V. mimicus was mixed with an equal volume of Freund’s complete or incomplete adjuvant (Sigma, Missouri, USA).
Fish (33.8±3.7 g) were divided into three groups (85 fish each). Fish were left untreated (control), or injected intra-peritoneal with 0.1 ml vaccine in Freund’s complete adjuvant, containing 2×108 colony forming units (CFU) (Group I) or 2×105 (GroupII) CFU inactivated V. mimicus. Two weeks after primary immunization, vaccinated fish were boosted with the same dose of bacteria in Freund’s incomplete adjuvant. Control fish were injected intra-peritoneal with 0.1 ml of PBS. At week 1, 2, 3 and 4 post primary immunization, heparin-treated or normal blood was collected from the tail vein of five fish in each group. Heparin-treated blood was used to measure respiratory burst activity of blood leukocytes. Normal blood was stored at room temperature for 1 h and then at 4 °C for 5 h. Serum was the collected by centrifugation at 12,000 rpm for 5 min to determine the agglutination antibody titer and lysozyme activity.
Challenge
The NH isolate was cultured in BHI broth at 28°C for 24 h and harvested by centrifugation at 5000 rpm for 5 min. Bacteria were washed three times, suspended in sterile PBS, and then adjusted to suitable concentration. We then evaluated the virulence of the NH isolate to channel catfish that were not vaccinated. Fish (15.1±1.8 g) were randomly divided into seven groups (one control group and six infection groups; 20 fish in each group). For infection, fish were injected intra-peritoneal with 0.1 ml of the bacterial suspension, at a concentration of 4.3×102, 4.3×103, 4.3×104, 4.3×105, 4.3×106, or 4.3×107 CFU/ml. Control fish were injected intra-peritoneal with 0.1 ml of PBS. Fish mortality was recorded daily for 14 days after inoculation. The median lethal dose (LD50) values were calculated using the trimmed Spearman–Karber method (Hamilton et al., 1977).
To evaluate the vaccine’s immune protection, four weeks after the primary immunization, 60 fish from each group were randomly selected and injected intra-peritoneally with 0.1 ml of the bacterial cultures containing 5.6×105 CFU of V. mimicus. Fish mortality was recorded daily for 14 days after the infection, the relative percentage survival (RPS) was calculated using the formula: (1-mortality of immunized fish/control fish mortality) × 100%.
Respiratory burst activity
Blood leukocyte respiratory burst activity was measured according to the method described by Anderson and AK (1995). Briefly, 0.1 ml of 0.2% nitro blue tetrazolium buffer was added to 0.1 ml anticoagulated blood, and the mixture was incubated at room temperature for 30 min. Next, 0.05 ml of the mixture were added into 1 ml dimethylformamide, and the mix was then centrifuged at 3000×g for 5 min. The supernatant was collected, and the absorbance value was detected at 540 nm. Dimethylformamide alone was used as a negative control.
Agglutination antibody titer
Serum agglutination antibody titers were determined in 96-well microplates with round bottoms. Heat-inactivated serum (50 μl) was added to each well in serial two-fold dilution. PBS was used as negative control. An equal volume of inactivated V. mimicus was added to each well containing serum samples. Microplates were incubated at 28°C overnight. The maximum dilution factor of the solutions that caused complete clumping of bacteria was considered the agglutination antibody titer.
Lysozyme activity
Serum lysozyme activity was detected using an LZM test kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, China). In brief, 100 ml of distilled water, standard liquid (supplied by the kit), or serum samples were added to 1 ml of a bacterial solution. After mixing and incubating at 37°C for 15 min, the mixture was transferred to an ice bath for 3 min. The suspension was transferred into a 0.5 cm optically clear colorimetric dish for transmittance (T15) determination at 530 nm. Transmittance of the distilled water at 530 nm was adjusted to 100% before measurements. Lysozyme content of the samples was calculated according to the following formula: Lysozyme content (U/ml) = (UT15-OT15)/ (ST15-OT15) × standard concentration (200 U/ml) × sample dilution factor where UT15 is test tube transmittance, OT15 is blank tube transmittance, and ST15 is standard tube transmittance.
Statistical analysis
Data are expressed as mean ± standard error. Significance of differences between samples was determined using Duncan’s test. The level of statistical significance was set at P < 0.05.
Ethical considerations
The authors agree upon standards of expected ethical behavior.
Identification of bacterium
Bacteria were isolated from the tissues of dying catfish, and the bacterial colonies were orbicular, smooth and white on blood agar, or green on TCBS agar (Figure 1B and C). Isolated bacteria were Gram-negative, curved rod-shaped, and had a single polar flagella (Figure 1D and F). BioMerieux VITEK system identified the bacterial isolates as V. mimicus, with 99% probability (Table 1). In addition, phylogenetic analysis of the 16S rDNA sequence showed that the NH strain clustered with V. cholerae CECT514T and V. mimicus ATCC33653T into the same group (Figure 2A). However, the phylogenetic analysis, based on the concatenated sequence of three housekeeping genes rpoA, recA, and pyrH, showed that the NH strain had a closer relationship to V. mimicus ATCC33653T than to V. cholerae CECT514T (Figure 2B).
Virulence of the isolate
The NH strain caused death of healthy channel catfish from day 1 post infection. Most deaths were observed at day 3 (~40%) and 4 (~100%) post infection. Infected fish exhibited the typical symptom of skin ulcers. Moreover, V. mimicus could be recovered from dead fish. The LD50 value of the isolate to catfish was 3.42×105 CFU per fish.
Vaccine protection
After V. mimicus infection, 56 fish (100%) died in the control group. However only one (Group I) and three (Group II) fish died in the vaccinated groups. The RPS of group I and group II were 98.2 and 94.6%, respectively.
Respiratory burst activity
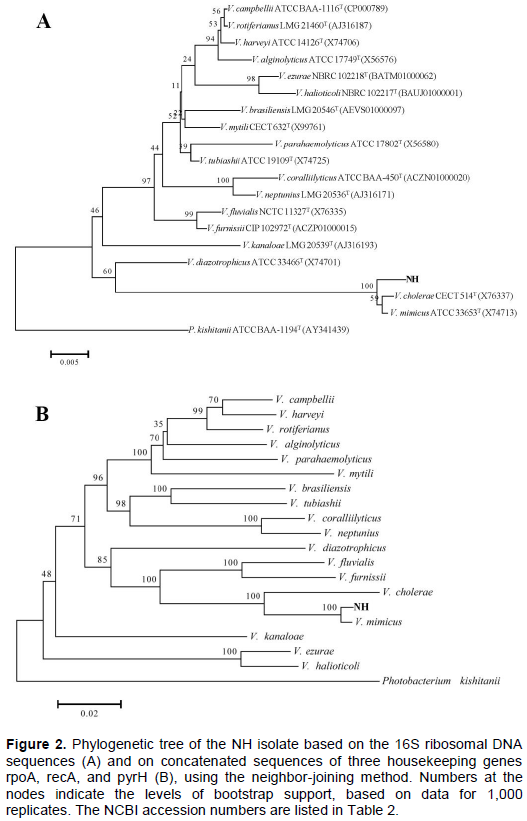
After immunization, the respiratory burst activity of blood leukocyte increased slightly, but there were no significant differences between the immunization groups and the control group at each time point (Figure 3).
Agglutination antibody titer
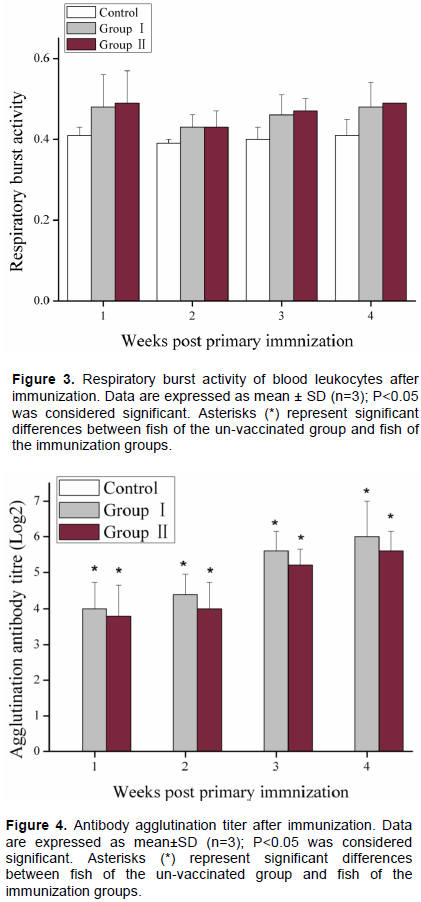
As shown in Figure 4, the serum agglutination antibody titer was significantly increased at week 1 post immunization, and increased continuously until 4 weeks after immunization. The agglutination antibody titer was not significantly different between vaccinated fish of Group I or II. In un-vaccinated fish no agglutination was detected during the course of experiment.
Lysozyme activity
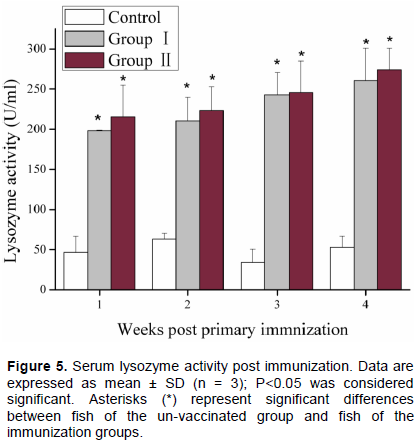
Similar to serum agglutination, serum lysozyme activity was significantly up-regulated at week 1 post immunization, and it continuously increased until the end of the experiment (Figure 5). The lysozyme activity did not change significantly between the two immunization groups (that is, Group I and II). There were small changes in lysozyme activity in the control group during the experiment, but differences were not significant.
V. mimicus can cause human gastroenteritis, ear infections, and severe cholera-like diarrhea (Austin 2010; Hasan et al., 2010). V. mimicusis is also associated with disease in aquatic animals, which are potential sources of infection to humans after ingestion (Miyoshi et al., 2014). Effective prophylaxis methods are needed to control V. mimicus epidemics. In this study, V. mimicus was isolated from dying channel catfish, and an effective prophylaxis method, a formalin-inactivated V. mimicus vaccine was developed.
According to the bacterial morphology and the character of gene sequences, the bacteria isolates were identified as V. mimicus. Subsequently, the virulence of the isolates obtained: V. mimicus exhibited high virulence to healthy channel catfish with a LD50 value of 3.42×105 CFU per fish. Then, we used formalin-inactivated V. mimicus as vaccine, which provided catfish with over 90% RPS. Similarly, in carp Cyprinus carpio, an inactivated V. mimicus vaccine (Zhang et al., 2014), and an OmpU-based vaccine (Cen et al., 2013), was also shown to protect against V. mimicus infection. These results indicate that immune prophylaxis is an effective method to control the disease caused by V. mimicus in both fish species, and maybe in other siluriformes species, such as yellow catfish P. fulvidraco (Geng et al., 2014), southern catfish S. soldatovi meridionalis (Geng et al., 2014), and Zhengchuan catfish S. soldatovi meridionalis (Zhang et al., 2014), which are also susceptible species.
Compared to the control group, the respiratory burst activity of blood leukocyte in the immunization groups increased slightly, but there were no significant differences between (P>0.05); the serum agglutination antibody titer significantly increased since immunization. After immunization with inactivated V. mimicus, serum agglutination antibody titer and lysozyme activity (but not the respiratory burst) increased significantly. Adaptive immunity is the basis for vaccine development, and antibodies play a crucial role in it. Fish can produce specific antibodies after immunization with inactivated V. mimicus (Zhang et al., 2014), or with a vaccine based on OmpU subunit (Cen et al., 2013). Li et al. (2016) observed a positive correlation between the antibody titer and immune protection. Similar results were also reported in grouper immunized with V. harveyi (Nguyen et al., 2017), in tilapia vaccinated with Streptococcus iniae or with S. agalactiae (Li et al., 2016; Zou et al., 2011), and in rainbow trout Oncorhynchus mykiss immunized with live attenuated Flavobacterium psychrophilum (Sudheesh and Cain, 2016), to name a few examples.
Lysozyme is an important component of innate immunity involved in host protection against microbial infection (Saurabh and Sahoo, 2008). Lysozyme is normally used as an indicator to evaluate the effect of vaccination on the humoral immune responses of fish. In the present study, serum lysozyme activity of immune I. punctatus was significantly up-regulated post immunization, and similar results were found in Scortum barcoo (Liu et al., 2014), and Anguilla rostrata (SongLin et al., 2015), indicating that lysozyme activity is up-regulated after immunization.
In conclusion, our results indicate that the RPS observed after vaccination is highly correlated to the agglutination titer of serum antibodies and lysozyme activity, but not to respiratory burst activity of blood leukocytes. Serum antibodies and lysozyme activity may play an important role in catfish immunity against V. mimicus infection.
The authors have not declared any conflict of interests.
The authors appreciate the support of the Special Funds for Marine Fishery Science and Technology Promotion Project of Guangdong Province (Grant No. YCN (2017)17H) given to Dr. Hong-Yan Sun.
REFERENCES
|
Adeleye IA, Daniels FV, Enyinnia VA (2010). Characterization And Pathogenicity of Vibrio spp. Contaminating Seafoods In Lagos, Nigeria. Internet Journal of Food Safety 12:1-9.
|
|
|
|
Ambas I, Suriawan A, Fotedar R (2013). Immunological responses of customised probiotics-fed marron, Cherax tenuimanus, (Smith 1912) when challenged with Vibrio mimicus. Fish and Shellfish Immunology 35:262-270.
|
|
|
|
|
Anderson DP, AK S (1995). Basic hematology and serology for fish health programmes. In: Shariff M, Arthur JR, Subasinghe RP, editors. Disease in Asian aquaculture. Manila: Fish Health Section of Asian Fisheries Society, pp. 185-202.
|
|
|
|
|
Austin B (2010). Vibrios as causal agents of zoonoses. Veterinary Microbiology 140:310-317.
|
|
|
|
|
Campos E, Bolaños H, Acuña MT, Díaz G, Matamoros MC, Raventós H, Sánchez LM, Sánchez O, Barquero C (1996). Vibrio mimicus diarrhea following ingestion of raw turtle eggs. Applied and Environmental Microbiology 62:1141-1144.
|
|
|
|
|
Cen J, Liu X, Li J, Zhang M, Wang W (2013). Identification and immunogenicity of immunodominant mimotopes of outer membrane protein U (OmpU) of Vibrio mimicus from phage display peptide library. Fish and Shellfish Immunology 34:291-295.
|
|
|
|
|
Chowdhury MA, Yamanaka H, Miyoshi S, Aziz KM, Shinoda S (1989). Ecology of Vibrio mimicus in aquatic environments. Applied and Environmental Microbiology 55:2073-2078.
|
|
|
|
|
Davis BR, Fanning GR, Madden .M, Steigerwalt AG, Bradford HB, Jr Smith HL, Jr Brenner DJ (1981). Characterization of biochemically atypical Vibrio cholerae strains and designation of a new pathogenic species, Vibrio mimicus. Journal of Clinical Microbiology 14: 631-639.
|
|
|
|
|
Eaves LE, Ketterer PJ (1994). Mortalities in red claw crayfish Cherax quadricarinatus associated with systemic Vibrio mimicus infection. Diseases of Aquatic Organisms 19:233-237.
|
|
|
|
|
Geng Y, Liu D, Han S, Zhou Y, Wang KY, Huang XL, Chen DF, Peng X, Lai WM (2014) Outbreaks of vibriosis associated with Vibrio mimicus in freshwater catfish in China. Aquaculture 433:82-84.
Crossref
|
|
|
|
|
Guardiola-Avila I, Acedo-Felix E, Sifuentes-Romero I, Yepiz-Plascencia G, Gomez-Gil B, Noriega-Orozco L (2016). Molecular and genomic characterization of Vibrio mimicus isolated from a frozen shrimp processing facility in Mexico. PloS One 11:e0144885.
|
|
|
|
|
Hamilton MA, Russo RC, Thurston RV (1977). Trimmed Spearman-Karber method for estimating median lethal concentrations in toxicity bioassays. Environmental Science and Technology 11:714-719.
|
|
|
|
|
Hasan NA, Grim CJ, Haley BJ, Chun J, Alam M, Taviani E, Hoq M, Munk AC, Saunders E, Brettin TS, Bruce DC, Challacombe JF, Detter JC, Han CS, Xie G, Nair GB, Huq A, Colwell RR (2010). Comparative genomics of clinical and environmental Vibrio mimicus. PNAS 107:21134-21139.
|
|
|
|
|
Li JN, Li YY, Hu SK, Li L, Fang B, Yu WY, Zhang XH (2005). Analysis on pathogen producing ascitic fluid disease of Eriocher sinensis. Journal of Fishery Sciences of China 12:267-274.
|
|
|
|
|
Li W, Wang HQ, He RZ, Li YW, Su YL, Li AX (2016). Major surfome and secretome profile of Streptococcus agalactiae from Nile tilapia (Oreochromis niloticus): Insight into vaccine development. Fish and Shellfish Immunology 55:737-746.
|
|
|
|
|
Liu L, Li YW, He RZ, Xiao XX, Zhang X, Su YL, Wang J, Li AX (2014). Outbreak of Streptococcus agalactiae infection in barcoo grunter, Scortum barcoo (McCulloch & Waite), in an intensive fish farm in China. Journal of Fish Diseases 37:1067-1072.
|
|
|
|
|
Liu X, Gao H, Xiao N, Liu Y, Li J, Li L (2015). Outer membrane protein U (OmpU) mediates adhesion of Vibrio mimicus to host cells via two novel N-terminal motifs. PloS One 10:e0119026.
|
|
|
|
|
Miyoshi SI, Ikehara H, Kumagai M, Mizuno T, Kawase T, Maehara Y (2014). Defensive effects of human intestinal antimicrobial peptides against infectious diseases caused by Vibrio mimicus and V. vulnificus. Biocontrol Science 19:199-203.
|
|
|
|
|
Nguyen HT, Thu Nguyen TT, Tsai MA, Ya-Zhen E, Wang PC, Chen SC (2017). A formalin-inactivated vaccine provides good protection against Vibrio harveyi infection in orange-spotted grouper (Epinephelus coioides). Fish and Shellfish Immunology 65:118-126.
|
|
|
|
|
Nugroho RA, Fotedar R (2013). Dietary organic selenium improves growth, survival and resistance to Vibrio mimicus in cultured marron, Cherax cainii (Austin, 2002). Fish and Shellfish Immunology 35:79-85.
|
|
|
|
|
Sang HM, Ky LT, Fotedar R (2009). Dietary supplementation of mannan oligosaccharide improves the immune responses and survival of marron, Cherax tenuimanus (Smith, 1912) when challenged with different stressors. Fish and Shellfish Immunology 27: 341-348.
|
|
|
|
|
Saurabh S, Sahoo PK (2008). Lysozyme: an important defence molecule of fish innate immune system. Aquaculture Research 39:223-239.
|
|
|
|
|
SongLin G, PanPan L, JianJun F, JinPing Z, Peng L, LiHua D (2015). A novel recombinant bivalent outer membrane protein of Vibrio vulnificus and Aeromonas hydrophila as a vaccine antigen of American eel (Anguilla rostrata). Fish and Shellfish Immunology 43:477-484.
|
|
|
|
|
Sudheesh PS, Cain KD (2016). Optimization of efficacy of a live attenuated Flavobacterium psychrophilum immersion vaccine. Fish and Shellfish Immunology 56:169-180.
|
|
|
|
|
Sun HY, Noe J, Barber J, Coyne RS, Cassidy-Hanley D, Clark TG, Findly RC, Dickerson HW (2009). Endosymbiotic Bacteria in the Parasitic Ciliate Ichthyophthirius multifiliis. Applied and Environmental Microbiology 75:7445-7452.
|
|
|
|
|
Takahashi A, Miyoshi S, Takata N, Nakano M, Hamamoto A, Mawatari K, Harada N, Shinoda S, Nakaya Y (2007). Haemolysin produced by Vibrio mimicus activates two Cl- secretory pathways in cultured intestinal-like Caco-2 cells. Cell Microbiology 9:583-595.
|
|
|
|
|
Thune RL, Hawke JP, Siebeling RJ (1991). Vibriosis in the red swamp crawfish. Journal of Aquatic Animal Health 3:188-191.
|
|
|
|
|
Wang GL, Zheng TL, Jin S, Lu TX (2003). Preliminary studies on pathogen of ascites disease of cultured Sparus macrocephelus. Chinese Journal of Veterinary Science 23:33-35.
|
|
|
|
|
Wong FYK, Fowler K, Desmarchelier PM (1995). Vibriosis due to Vibrio mimicusin Australian freshwater Crayfish. Journal of Aquatic Animal Health 7:284-291.
|
|
|
|
|
Zhang X, Li YW, Mo ZQ, Luo XC, Sun HY, Liu P, Li AX, Zhou SM, Dan XM (2014). Outbreak of a novel disease associated with Vibrio mimicus infection in fresh water cultured yellow catfish, Pelteobagrus fulvidraco. Aquaculture 432:119-124.
|
|
|
|
|
Zhang YQ, Zhang TT, Li JN, Liu XL, Li L (2014). Design and evaluation of a tandemly arranged outer membrane protein U (OmpU) multi-epitope as a potential vaccine antigen against Vibrio mimicus in grass carps (Ctenopharyngodon idella). Veterinary Immunology and Immunopathology 160:61-69.
|
|
|
|
|
Zou LL, Wang J, Huang BF, Xie MQ, Li AX (2011). MtsB, a hydrophobic membrane protein of Streptococcus iniae, is an effective subunit vaccine candidate. Vaccine 29:391-394.
|
|