Full Length Research Paper

ABSTRACT
The mangrove sediments are rich in organic matter and humic substances, responsible for important functions such as reducing the toxicity of heavy metals, nutrient stabilization, serving as drain to atmospheric carbon and increasing the plant growth. In this study, we observed the effect of humic substances, humic acids and fulvic acids isolated from sediments of a mangrove forest, sampled from the Municipal Ecological Station Ilha do Lameirão (EEMIL, located in Vitória, Espírito Santo State, Brazil), on the growth and root acidification of Rizophora mangle and Laguncularia racemosa seedlings. For this, R. mangle and L. racemosa propagules were transferred to pots and then treated with different humic materials. The evaluation in root architecture change was performed by analyzing the main root axis length, lateral root length, density of lateral roots, fresh and dry mass weight of roots and estimated H+-ATPase activity by specific root acidification measurement. The results indicate that all humic materials extracted from mangrove organic matter were able to modify the root architecture systems of the studied plants. In addition to inducing an increase in the number of lateral roots and root branching, it also stimulated specific root acidification when compared to control groups. There is a good potential in developing technologies for the production of seedlings of mangrove plant species treated with biostimulants based in humic materials isolated from the mangrove ecosystem itself.
Key words: Humic substances, humic acids, fulvic acids, mangrove.
Abbreviation: EEMIL, Estação Ecológica Municipal Ilha do Lameirão (Municipal Ecological Station Ilha do Lameirão); HS, humic substances; HA, humic acids; FA, fulvic acids.INTRODUCTION
Periodical flooding from fluvial or marine origin that may occur in mangroves generates accumulation of organic matter ecosystem, leading to a carbon fixation amount, around 0.57 Mg per ha per year (Ferreira et al., 2007), which highlights the mangrove’s importance in the cycling of atmospheric C. The biggest amount of the organic matter from soils, water and sediments is composed of humic substances (HS) (Baldotto, et al., 2013). According to Stevenson (1994), the HS covers heterogeneous organic compounds with no clear biochemical classi-fication, produced as by-products of microbial metabolism. Despite the long period of study and experimentation, only recently we came to an understanding of what would these substances. Piccolo (2002) showed that HS are aggregates of organic compounds that are held together by weak interactions in a supramolecular conformation of apparently high mass. These substances promote direct effects on plant metabolism, stimulating plant growth as a whole, stimulating flowering and by consequence improving plantation yield (Vaz and Gonçalves, 2002; Rocha et al., 2004; Benites et al., 2006). They are also able to indirectly enhance soil resistance to water and nutritional deficiencies, providing better fertility and improving the system’s physical and biological conditions (Guminski, 1968; Busato et al., 2009).
Many studies support the benefit generated by the use of products based on humic substances (HS) as biostimulant to different plant species incrementing, especially, root growth (Canellas et al., 2002; Dobbss et al., 2010; Aguiar et al., 2013; Canellas et al. 2012; Amorim et al., 2015; Silva et al., 2015; Ramos et al., 2015). However, further studies to quantify and characterize the specific amounts to be applied in certain species of mangrove as Rizophora mangle and Laguncularia racemosa remain scarce.
The fraction of HS originates from three products, classified according to their solubility in acid or alkaline, they are: Humin, fulvic acid (FA) and humic acid (HA) (Kononova, 1966; Canellas and Santos, 2005). The first fraction, humin, is less evolved and more stable being strongly linked to the mineral fraction of the soil, and thus is the insoluble portion of the organic matter. The second fraction (FA) constitutes the water-soluble portion, consisting of low apparent molecular mass molecules and higher amount of acid functional groups. The last, fraction (HA) is composed by molecules soluble only in alkaline medium with high apparent molecular mass (Schnitzer, 1982; Canellas et al., 2001).
According to Façanha et al. (2002), better plant development promoted by treatment with HS can be attributed to increased permeability of the plasmatic membrane, and activation of the transmembrane enzyme H+-ATPase, which pumps H+ into intracellular space. The acidification of the cell environment increases the plasticity of the membrane, allowing the cell to stretch wider and thus promoting the mechanism know as "acid growth" (Hager, 2003).
The aim of this study is to evaluate the effect of different humic fractions extracted from mangrove sediment on the root system, and specific root acidification in seedlings of Rizophora mangle and Laguncularia racemosa.
MATERIALS AND METHODS
Sampling area, chemical and physical composition of the sediments, location and history
Sediment samples from a mangrove ecosystem were collected at the surface of an area with very few anthropic impacts (Escravos canal region) in the EEMIL, Vitória, Espírito Santo State. The chemical and physical composition of the sediment used to extract the humic materials is shown in Table 1. The station is located between the latitudes 20° 14’ S to 20° 17’ S and longitudes 40° 16’ W to 40° 20’ W (Tulli, 2007), and covers an area of 891.8 ha, where 92.7% are represented by the mangrove. Such area was, initially, made into a preservation unit of Vitória-ES as a Municipal Biological Reserve, by publishing of the Municipal Law No. 3326 of May 27, 1986. It was later transformed into the EEMIL by the Municipal Law No. 3377 of September 12, 1986 (Tulli, 2007).

Humic substance extraction from organic matter of mangrove sediment and elemental composition of humic materials utilized
The extraction of HS, to obtain the FA and HA fractions were made in accordance to the classical methodology adopted by the International Humic Substances Society (IHSS) (Schnitzer, 1982). Explaining it briefly, 200 g of sediment samples (in triplicate) were air dried and sieved (2 mm mesh sieve), extraction of HS was done using NaOH 0.5 mol L-1, in a sediment: solvent ratio of 1:10 (m:v). The separation of HA was achieved by lowering the solution’s pH to 1.0 to 1.5 with HCl 6.0 mol L-1 followed by centrifugation (890 rcf / 20 min). Dissolutions and precipitations were repeated three times; all humic materials were adjusted to pH 7.0 utilizing NaOH 0.5 mol L-1 or HCl 6.0 mol L-1, properly purified according to Canellas et al. (2005).
After purification and subsequent lyophilization, the humic materials (HS, FA and HA) had their elemental composition analyzed (CHNO) through an elemental analyzer device (CHNS - 932, Leco, Germany) with 4.0 mg samples (in triplicate). Oxygen content was determined by oxygen-difference and ash from incineration of 50 mg of the humic materials at 700°C during 8 h. The CHNO analysis is often used to relate chemical properties of humic substances with the genesis or properties of the extraction origin and this present work was basically used for the calculations of doses (in mmol C L-1) were used in preliminary experiments concentration response. The values obtained for CHNO humic materials were as follows: HS (C: 21.61% H: 2.02% N: 2.06% and O: 74.31%); HA (C: 48.93% H: 5.76% N: 4.09% and O: 41.21%) and AF (C: 34.30% H: 4.63% N: 6, 35% and O: 65.53%).
Concentration response test with humic materials used on treatment of Laguncularia racemosa and Rizophora mangle seedlings and experiment with the best concentration
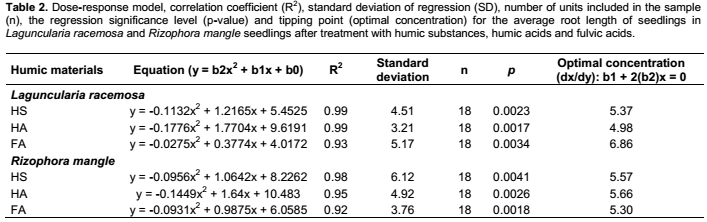
Seedlings Laguncularia and Rizophora were exposed to all the humic materials (HS, FA and HA) extracted from the mangrove ecosystem. In order to obtain the best doses, at witch plant growth is greatly improved, the following concentrations were used for each humic material: 0.0 (control); 1.5; 3.0; 6.0 and 12.0 mmol C L-1. Following a regression analysis, a new experiment was performed, this time using the optimal dose of each humic material (Table 2), for comparing various treatments and the control.
Propagules of L. racemosa and R. mangle brought from the study area (Escravos channel region) developed during 30 days were treated with the best concentrations of the humic materials found (Table 1). Following exposure to the treatments, the number of lateral roots was assessed using a computer program to analyze digital images (ImageJ® v.1.45) and the root main axis length. Also, fresh and dry root mass were measured using a scale with analytical precision, dry weight was measured after 72 h on a stove at 60ºC. Each treatment with the best concentrations of humic material had 8 pots and each pot had 2 propagules, giving a total sample size of 16 plants per treatment for each humic material (HS, FA and HA).
The experiment was completely randomized, with 3 replicates per treatment, whose mathematical model can be described as: Yij = µ + ti + eij. As it follows: Yij = experimental response, measured in the experimental unit j, submitted to the treatment i; µ = overall average; ti = relative effect by the treatment i; eij = random error. An analysis of variance was performed and means compared by the Tukey test (P <0.05) using the software SAEG v.9.1.
Measure of acidity in the growing solution - estimate of the activity H+ -ATPase
Estimation of H+-ATPase activity was assessed by measuring the acidity in a solution containing seedlings of L. racemosa and R. mangle treated with the best concentration of HS obtained in preliminary test concentration response test. In this experiment, only minimal medium (CaCl2 2 mmol) was used to avoid any influence of nutrients which could act in synergy with the HS, stimulating root growth and/or overall plant metabolism. A total of 5 plants (in triplicate) with similar age and height were treated with HS (along with a control) for 48 h and after this period, they were transferred to a recipient containing 50 mL of a CaCl2 2.0 mmol solution at pH 7.0. The solution’s pH was measured using a potentiometer able to measure pH with a glass electrode, during 140 minutes. The roots were digitalized for latter analysis and then dried in a forced air stove, after that the amount of H+ was expressed in moles of H+ per gram of dry roots (mmol H+ g of roots1) (Aguiar et al., 2013).
RESULTS AND DISCUSSION
Effects of the humic materials on root growth
Changes in root system architecture can be variable in response to hormonal and environmental stimulations (Torrey, 1986; López-Bucio et al., 2003; Sorin, 2005). All the humic products (HS, HA and FA) isolated from the mangrove ecosystem sediment showed the ability to induce these changes in rooting pattern in seedlings of L. racemosa and R. mangle (Figures 2 and 3).
The quadratic regression curves for the mean root length of L. racemosa and R. mangle seedlings treated with the different humic materials are illustrated in Figure 1.
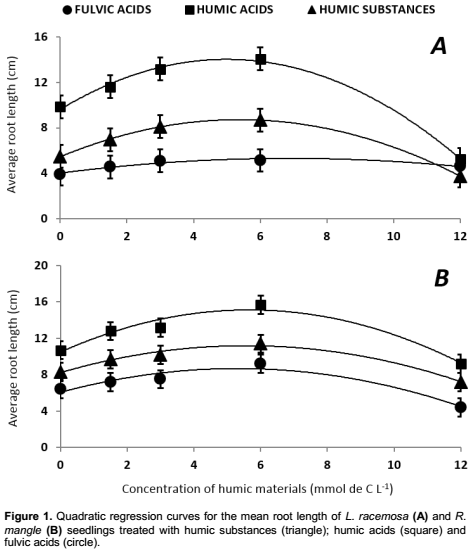
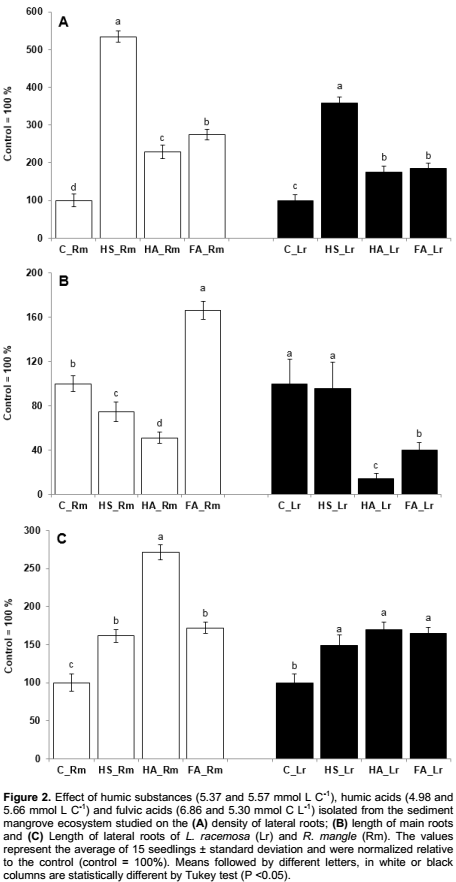
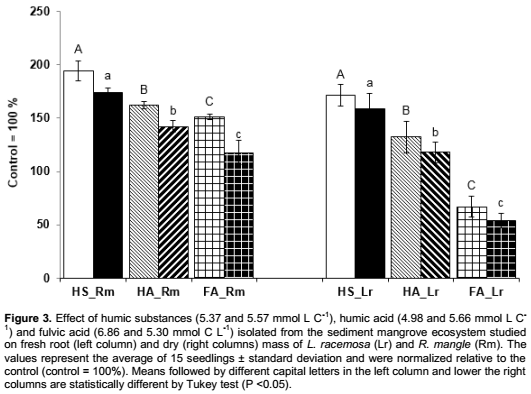
Table 2 and Figure 1 show the concentration-response model and the optimum concentrations for each humic material for seedlings of L. racemosa and R. mangle. The results of the average increase in root length, obtained by the first derivative of a quadratic regression, for the effect of concentration in seedlings of L. racemosa and R. mangle respectively, the results were 5.37 and 5.57 mmol C L-1 for HS; 4.98 and 5.66 mmol C L-1 for HA; 6.86 and 5.30 mmol C L-1 for FA (Table 1).
Roots architecture of the seedlings from both species were significantly altered by the addition of the HS to the culture, which results are shown in Figures 2 and 3. Also, the density of lateral roots of both species was significantly altered by the presence of humic substances. Similar results were observed by Dobbss et al. (2007, 2010), Aguiar et al. (2009), Aguiar et al. (2013). However, by comparing the two species, it was observed that seedlings of R. mangle showed a greater number of lateral roots per unit of primary root length (lateral roots density). Seedlings of L. racemosa also showed statistically higher values compared to the control plants (Figure 2A). Such results are probably related to the specific development characteristics of each studied species.
Regarding the length of the main roots, results showed a shortening caused by treatment with HS. This result is very similar to the behavior of plants treated with high concentrations of auxin (Peres et al., 2009), as auxin has already been found in the supramolecular structure of HS (Muscolo et al., 1998). However this “auxinic” effect was significant for seedlings of R. mangle and not significant for seedlings of L. racemosa (Figure 2B).
The increase in length of emerged lateral roots was 48 and 72% higher for L. racemosa and R. mangle respectively for seedlings treated with HS when compared to control seedlings (Figure 2C). Induction in lateral roots growth has been observed in other plant species as reported, indicated, highlighted by Canellas et al. (2002, 2008, 2009 and 2010), Dobbss et al. (2007, 2010), Baldotto et al. (2011), Amorim et al. (2015), Silva et al. (2015) and Ramos et al. (2015), suggesting an occupation and soil exploitation strategy by plants treated with HS. Lateral roots explore more soil around them and compete less with each other due to the distance between them (Silva and Delatorre, 2009).
The results of fresh and dry root mass (Figure 3) were significantly higher in treatments with HS for both plant species, when compared to their respective control treatment, which corroborates similar to the findings of Canellas et al. (2009), where the observed increases in root mass were 100% higher in seedlings treated with HS.
The HA stimulated lateral rooting significantly for both species, reflecting in increases to root density, fresh and dry masses (Figures 2A and 3). The growth of the main roots was strongly inhibited by the HA in a behavior typical to plants treated with auxins (Dobbss et al., 2007; Zandonadi et al., 2007) (Figure 2B). Auxin is a key hormone for the regulation of lateral root emission in plants (Blakely et al., 1982). As expected for effects of exogenously applied hormones, auxin action on the root development is dependent on its concentration as reported by Mulkey et al. (1982). Casimiro et al. (2001) demonstrated that polar transport of auxin in both directions, basipetal (from the tip towards the base of the root) and acropetal (from the base towards the tip of the root) is necessary for the initiation and emergence of lateral roots. Stimulation of lateral roots emission and shortening of the main root, at the same time, resulted in an increase in the average density of lateral roots of 75 and 111% for L. racemosa and R. mangle respectively, when compared to their control groups (Figure 2A).
The greater effect of HA on fresh and dry weight of roots was observed in seedlings R. mangle (Figure 3). The promoted HA increases in the length of lateral roots in the order of 69 and 183% in L. racemosa and R. mangle respectively, when compared to their control groups (Figure 2C). According to Conceição et al. (2008), root growth occurs in two stages: Meristematic growth and vacuolated growth; the latter occurs in the stretching zone and is characterized by rapid expansion of cells, sustained by increased water uptake by the vacuoles through turgid pressure. Moreover, Cosgrove (1998) and Zandonadi et al. (2007) found that HA isolated from various sources of organic matter strongly stimulates the pumping of H+ by V-ATPase and H+-pirofosfatase. Also, Canellas et al. (2010) and Dobbss et al. (2010) observed that different humified materials are capable of promoting growth and various changes in the geometry of the root system, improving uptake of water and nutrients and resulting, as observed, in higher fresh plant mass and root growth.
The FA, extracted from the HS obtained from the mangrove sediment, also stimulated plant rooting significantly, markedly changing the root architecture of L. racemosa and R. mangle seedlings (Figures 2 and 3). Dobbss et al. (2007) observed similar results, where the authors found significant stimuli of the fraction FA to rooting Arabidopsis seedlings both in number and in length of lateral roots. The FA (that corresponds to the soluble humic fraction of the HS), at any pH value, caused an increase in lateral roots density of 196% in R. mangle seedlings and 84% increase in L. racemosa seedlings, when compared to their respective control groups (Figure 2A).
Results obtained regarding main root length were quite contrasting. An increase in the length of the main root in R. mangle and a reduction of main root length in L. racemosa seedlings after treatment with FA (Figure 2B) was observed. According to Zandonadi (2010), this difference between species is due to the fact that, the regulation of root architecture is quite complex and varies, both, in different species and within the same species.
Regarding dry and fresh root masses, it could be observed the same contrasting tendency as in main root length, an increase in plant mass to R. mangle and a decrease in plant mass to L. racemosa (Figure 3). In relation to lateral root length, significant increases were observed in both species when treated with FA (Figure 2C). According to the traditional view of the chemical structure of HS, it was postulated that "in the rhizosphere, the interaction between roots and the organic matter is possible when the humic molecules present in the soil solution are small enough to flow through the apoplast and reach the plasmatic membrane" (Muscolo et al., 2007). Thus, other works described in literature (Nardi et al., 2002 and Quaggiotti et al., 2004) consider that only the FA, known as the fraction of organic matter with lowest molecular mass, could promote increments to growth and changes in the energetic metabolism of plants. However, according to our results, we can reinforce the idea that regardless of molecular size (Canellas et al., 2010) different humidified fractions can stimulate plant growth in various ways and for different plant species. The findings of this study also collaborate with the data previously obtained by Aguiar et al. (2009) using exclusion chromatography on sephadex gel, that did not found any relationship between the molecular mass distribution and the bioactivity of HS.
Measure of acidity in the growing solution - estimate of the activity H+ -ATPase
Figure 4 shows the pH measures of the medium containing L. racemosa and R. mangle seedlings treated with and without (control) HS extracted from the mangrove ecosystem. The increase in acidity of the solution was observed in treatments in which HS exposed the plants for 48 h. These results corroborate with those obtained by Dobbss et al. (2008) and Silva et al. (2015) which also observed increases in acidity of solutions containing Arabidopsis thaliana seedlings treated with humic materials extracted from the Paraiba do Sul River and solutions with seedlings of Cattleya warneri treated with HA from vermicompost, respectively. Zandonadi et al. (2010) directly related root acidification with specific activity of H+-ATPase in seedlings roots treated with HA and recently Aguiar et al. (2013) positively correlated proton extrusion in seedlings roots of corn plants treated with the HA isolated from many vermicomposts with the activity of H+-ATPase of plasmatic membrane. According to Aguiar (2011) increased acidity of the medium may be primarily associated with two main factors: (i) Production of CO2 by root respiration, as the breathing process produces CO2, that dissolves in to the medium and causes a reduction of pH; and (ii) An increase in H+ extrusion, possibly associated with the activity of HS on H+-ATPase.
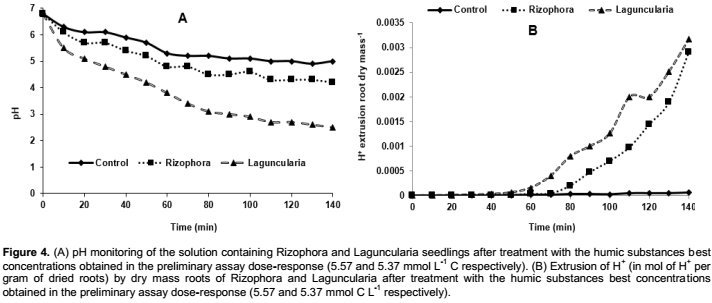
As recently reported by Aguiar et al. (2013), despite the acidity alterations provided by exposing the seedlings to HS not being exclusively related to stimulation of H+ pumps, it may be suggested that this simplified method may be used in the study of physiologically active HS. These results were expected since the cell growth promoting mechanism is mediated by the H+-ATPases in a process known as "acid growth theory." The process of H+-ATPases activation that culminates in cell expansion starts with the generation of the H+ gradient and lowering of the pH, provided by the accumulation of H+ in the extracellular side (Hager, 2003).
Therefore, based on the data obtained one can estimate that, at least in part, the measures of acidity in solutions containing plants treated HS is probably related to the activity of H+-ATPase (increase in H+ extrusion) in the plasmatic membrane.
CONCLUSIONS
Different humic materials (HS, HA and FA) isolated from mangrove forest sediment, from the EEMIL region, presented the ability to modify the architecture of root systems in R. mangle and L. racemosa, leading to increases in the number and length of lateral roots, and also the fresh and dry mass weight of roots. The humic substances, when in solution, acted as regulators of plant growth, since, possibly, were able to stimulate the activity of H+-ATPase by acidity measurements in solution assays.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
The authors thank Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (Capes), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and Fundação Nacional de Desenvolvimento do Ensino Superior Particular (FUNADESP) for financial support. The Secretaria Municipal de Meio Ambiente (SEMMAM) from Prefeitura Municipal de Vitória (PMV) is also appreciated for the fundamental assistance towards the development of the work within the Estação Ecológica Municipal Ilha do Lameirão (EEMIL).
ABBREVIATIONS
EEMIL, Estação Ecológica Municipal Ilha do Lameirão (Municipal Ecological Station Ilha do Lameirão); HS, humic substances; HA, humic acids; FA, fulvic acids.
REFERENCES
|
Aguiar NO (2011). Características químicas e bioatividade de ácidos húmicos isolados de vermicompostos em diferentes estádios de maturação. Dissertação de mestrado. Universidade Estadual do Norte Fluminense (UENF), Darcy Ribeiro, Campos dos Goytacazes, RJ, Brazil. 86 p. |
|
|
Aguiar NO, Canellas LP, Dobbss LB, Zandonadi DB, Olivares FL, Façanha AR (2009). Distribuição de massa molecular de ácidos húmicos e promoção de crescimento radicular. Rev. Bras. Ciênc. Solo. 33:1613-1623. |
|
|
Aguiar NO, Olivares FL, Novotny EH, Dobbss LB, Balmori DM, Santos-Júnior LG, Chagas JG, Façanha AR, Canellas LP (2013). Bioactivity of humic acids isolated from vermicomposts at different maturation stages. Plant Soil 362:161-174. |
|
|
Amorim MM, Vieira HD, Amorim IM, Dobbss LB, Deminicis BB, Xavier PB (2015). Effects of the humic acid extracted from vermicompost on the germination and initial growth of Brachiaria brizantha cv. MG5. Afr. J. Biotechnol. 14:1576-1583. |
|
|
Baldotto MA, Gobo AAR, Salomão MSMB, Rezende CE, Camargo PB (2013). Fractions of organic matter and redox properties of humic substances in sediments from deep oceans. Quím. Nova 36:1288-1295. |
|
|
Baldotto MA, Muniz RC, Baldotto LEB, Dobbss LB (2011). Root growth of Arabidopsis thaliana (L.) Heynh. treated with humic acids isolated from typical soils of Rio de Janeiro state, Brazil. Rev. Ceres 58:504-511. |
|
|
Benites VM, Polidoro JC, Menezes CC, Betta M (2006). Aplicação foliar de fertilizante organo mineral e soluções de ácido húmico em soja sob plantio direto. Circular técnica 35. Embrapa, 6 p. |
|
|
Blakely LM, Durham M, Evans TA, Blakely RM (1982). Experimental studies on lateral root-formation in radish seedling roots I. general-methods developmental stages, and spontaneous formation of laterals. Bot. Gaz. 143:341-352. |
|
|
Busato JG, Canellas LP, Dobbss LB, Baldoto MA, Aguiar NO, Rosa RCC, Schiavo JA, Marciano CR, Olivares FL (2009). Guia para adubação orgânica baseado na experiência com solos e resíduos do Norte Fluminense. Manual Técnico 14, Programa Rio Rural, Rio de Janeiro, 28 p. |
|
|
Canellas LP, Dobbss LB, Oliveira AL, Chagas JG, Aguiar NO, Rumjaneck VM, Novotny EH, Olivares FL, Spaccini R, Piccolo A (2012). Chemical properties of humic matter as related to induction of plant lateral roots. Eur. J. Soil Sci. 63:315-324. |
|
|
Canellas LP, Olivares FL, Okorokova-Façanha AL, Façanha AR (2002). Humic acids isolated from earthworm compost enhance root elongation, lateral root emergence, and plasma membrane H+-ATPase activity in maize roots. Plant Physiol. 130:1951-1957. |
|
|
Canellas LP, Piccolo A, Dobbss LB, Spaccini R, Olivares FL, Zandonadi DB, Façanha AR (2010). Chemical composition and bioactivity properties of size-fractions separated from a vermicompost humic acid. Chemosphere 78:457-466. |
|
|
Canellas LP, Santos GA (2005). Humosfera: tratado preliminar sobre a química das substâncias húmicas. Luciano Pasqualoto Canellas e Gabriel Araújo Santos, Campos dos Goytacazes, RJ. 309 p. |
|
|
Canellas LP, Santos GA, Rumjanek VM, Moraes AA, Guridi F (2001). Distribuição da matéria orgânica e características de ácidos húmicos em solos com adição de resíduos de origem urbana. Pesq. Agropec. Bras. 36:1529-1538. |
|
|
Canellas LP, Spaccini R, Piccolo A, Dobbss LB, Okorokova-Façanha AL, Santos GA, Olivares FL, Façanha AR (2009). Relationships between chemical characteristics and root growth promotion of humic acids isolated from brazilian oxisols. Soil Sci. 174:611-620. |
|
|
Canellas LP, Teixeira Junior LRL, Dobbss LB, Silva CA, Médici LO, Zandonadi DB, Façanha AR (2008). Humic acids cross interactions with root and organic acids. Ann. Appl. Biol. 153:157-166. |
|
|
Casimiro I, Marchant A, Bhalerao RP, Beeckman T, Dhooge S, Swarup R, Graham N, Inzé D, Sandber G, Casero PJ, Benett M (2001). Auxin Transport Promotes Arabidopsis Lateral Root Initiation. Plant Cell 13:843-852. |
|
|
Conceição PM, Vieira HD, Canellas LP, Júnior RBM, Olivares FL (2008). Recobrimento de sementes de milho com ácidos húmicos e bactérias diazotróficas endofíticas. Pesq. Agropec. Bras. 43:545-548. |
|
|
Cosgrove DJ (1998). Cell Wall Loosening by expansins. Plant Physiol. 118:333-339. |
|
|
Dobbss LB, Baldotto MA, Zandonadi DB, Canellas LP, Façanha AR, Rezende CE (2008). River organic matter affects Arabidopsis thaliana root architecture and proton extrusion. In. Sociedade Brasileira de Bioquímica e Biologia Molecular (SBBQ 2008). Águas de Lindóia, SP, Brasil. Anais, SBBQ 2008. CD-ROM. |
|
|
Dobbss LB, Canellas LP, Olivares FL, Aguiar NO, Peres LEP, Azevedo M, Spaccini R, Piccolo A, Façanha AR (2010). Bioactivity of chemically transformed humic matter from vermicompost on plant root growth. J. Agric. Food Chem. 58:3681-3688. |
|
|
Dobbss LB, Medici LO, Peres LEP, Pino-Nunes LE, Rumjanek VM, Façanha AR, Canellas LP (2007). Changes in root development of Arabidopsis promoted by organic matter from oxisols. Ann. Appl. Biol. 151:199-211. |
|
|
Façanha AR, Olivares FL, Velloso ACX, Braz-Filho R, Santos GA, Canellas LP (2002). Bioatividade de ácidos húmicos: efeitos sobre o desenvolvimento radicular e sobre a bomba de prótons da membrana plasmática. Pesq. Agropec. Bras. 37:1301-1310. |
|
|
Ferreira TO, Vidal-Torado P, Otero XL, Macías F (2007). Are mangrove Forest substrates sediments or soils? A case study in southeastern Brazil. Catena 70:79-91. |
|
|
Guminski S (1968). Present days view on physiological effects induced in plant organisms by humic compounds. Soviet Soil Sci. 9:1250-1255. |
|
|
Hager A (2003). Role of the plasma membrane H+-ATPase in auxin-induced elongation growth: historical and new aspects. J. Plant Res. 116:483-505. |
|
|
Kononova MM (1966). Soil organic matter, its nature, its role in soil formation and in soil fertility. Oxford, Pergamon Press, 543 p. |
|
|
López-Bucio J, Cruz-Ramírez A, Herrera-Estrella L (2003). The role of nutrient availability in regulating root architecture. Curr. Opin. Plant Biol. 6:280-287. |
|
|
Mulkey TJ, Kuzmanoff KM, Evans ML (1981). The agar-dye method for visualizing acid efflux patterns during tropistic curvatures. What's New in Plant Physiology 12:9-12. |
|
|
Muscolo A, Cutrupi S, Nardi S (1998). IAA detection in humic substances. Soil Biol. Biochem. 30:1199-1201. |
|
|
Muscolo A, Francioso O, Tugnoli V, Nardi S (2007). The auxin-like activity of humic substances is related to membrane interactions in carrot cell cultures. J. Chem. Ecol. 33:115-129. |
|
|
Nardi S, Pizzeghello D, Muscolo A, Vianello A (2002). Physiological effects of humic substances on higher plants. Soil Biol. Biochem. 34:1527-1536. |
|
|
Peres LEP, Zsögön A, Kerbauy GB (2009). Abscisic acid and auxin accumulation in Catasetum fimbriatum roots growing in vitro with high sucrose and mannitol content. Biol. Plant. 53:560-564. |
|
|
Piccolo A (2002). The Supramolecular Structure of Humic Substances: A Novel Understanding of Humus Chemistry and Implications in Soil Science. Adv. Agron. 75:57-134. |
|
|
Quaggiotti S, Ruperti B, Pizzeghello D, Francioso O, Tugnoli V, Nardi S (2004). Effect of low molecular size humic substances on nitrate uptake and expression of genes involved in nitrate transport in maize (Zea mays L.). J. Exp. Bot. 55:803-813. |
|
|
Ramos AC, Dobbss LB, Santos LA, Fernandes MF, Olivares FL, Aguiar NO, Canellas LP (2015). Humic matter elicits proton and calcium fluxes and signaling dependent on Ca2+-dependent protein kinase (CDPK) at early stages of lateral plant root development. Chem. Biol. Technol. Agric. 2:1-12. |
|
|
Rocha GN, Gonçalves JLM, Moura IM (2004). Mudanças da fertilidade do solo e crescimento de um povoamento de Eucalyptus grandis fertilizado com biossólido. Rev. Bras. Ciênc. Solo 28:623-639. |
|
|
Schnitzer M (1982). Organic matter characterization. In: Method of Soil Analysis, Part 2. A.L. Page; R.H. Miler & D. Keeney (eds.). Agronomy series no 9. American Society Agronomy and Soil Science Society of America, ASA/SSSA Publishers, Madison, USA. pp. 581-597. |
|
|
Silva AA, Delatorre CA (2009). Root architecture modification in response to phosphorus and nitrogen availability. Rev. Ciênc. Agrovet. 8:152-163. |
|
|
Silva MAC, Santos WO, Simoura NT, Tesch, JA, Ruas KF, Colodete CM, Tannure FP, Barbirato JO, Ramos AC, Dobbss LB (2015) Ácidos húmicos de vermicomposto estimulam o crescimento in vitro de plântulas de Cattleya warneri (Orchidaceae). Rodriguésia 66:759-768. |
|
|
Sorin C, Bussell JD, Camus I, Ljung K, Kowalczyk M, Geiss G (2005). Auxin and light control of adventitious rooting in Arabidopsis require argonaute. Plant Cell 17:1343-1359. |
|
|
Stevenson FJ (1994). Humus chemistry; genesis, composition, reactions. New York: John Wiley & Sons, 496 p. |
|
|
Torrey JG (1986). Endogenous and exogenous influences on the regulation of lateral root formation. Developments in plant and soil sciences. In. Jackson M.B. (Ed.) New Root Formation in Plants and Cuttings. Martinus Nijhoff. pp. 32-66. |
|
|
Tulli LMA (2007). Vulnerabilidade à ação antrópica e uso e ocupação do solo para a estação Ecológica Municipal Ilha do Lameirão, Vitória-ES. Dissertação de Mestrado. Universidade Federal do Espírito Santo, Vitória, ES, Brasil. 106 p. |
|
|
Vaz LMS, Gonçalves JLM (2002). Uso de biossólidos em povoamento de Eucalyptus grandis: Efeito em atributos químicos do solo, no crescimento e na absorção de nutrientes. Rev. Bras. Ciênc. Solo 26:747-758. |
|
|
Zandonadi DB (2010). Energização e sinalização dos mecanismos de regulação do desenvolvimento radicular via modulação das bombas de H+ por ácidos húmicos e fitormônios. Tese de Doutorado. Universidade Estadual do Norte Fluminense Darcy Ribeiro, Campos dos Goytacazes, RJ, Brasil. 156p. |
|
|
Zandonadi DB, Canellas LP, Façanha AR (2007). Indolacetic and humic acids induce lateral root development through a concerted plasmalemma and tonoplast H+ pumps activation. Planta 225:1583-1595. |
|
|
Zandonadi DB, Santos MP, Dobbss LB, Olivares FL, Canellas LP, Binzel ML, Okorokova-Façanha AL, Façanha AR (2010). Nitric oxide mediates humic acids-induced root development and plasma membrane H+-ATPase activation. Planta 231:1025-1036. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


