Review

ABSTRACT
Medicinal and edible mushrooms have biological properties that are important in promoting health. In the last decades, an increasing number of studies have explored various aspects of the cultivation and metabolism of various species. Many isolated substances have potential for use in medicine and among them are polysaccharides, which have been shown to have physiological properties such as anti-tumor, immunomodulatory, anti-hypercholesterolemia, antiviral and antiinflammatory. The most common way of obtaining products from mushrooms is through cultivation in a solid medium. However, in this type of cultivation there is a difficulty in controlling the physical and chemical parameters and, consequently, the quality of the product. In this context, the submerged fermentation of the mycelial form of the mushroom has received attention as an alternative for the efficient production of biomass and, therefore, of polysaccharides. Technical problems and forms of cultivation are studied so that the standardization of cultivation is more effective. This review describes some aspects about the submerged fermentation technique in bottles and bioreactors, exploring the two main ones, which are the stirred tank fermentator and the air lift reactors and the species that are in these cultivated systems.
Key words: Mushrooms, polysaccharides, submersed fermentation, biorreactor.
INTRODUCTION
CULTIVATION IN SUBMERGED FERMENTATION: A NEW PERSPECTIVE OF CULTIVATION
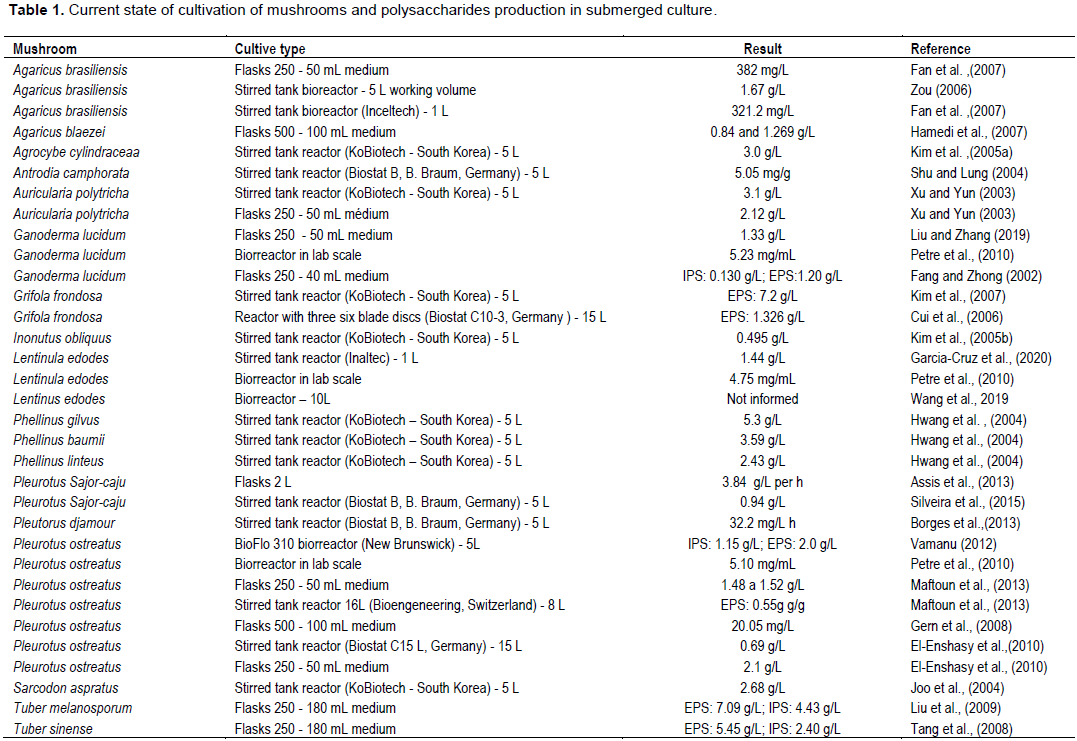
CONCLUSION AND PERSPECTIVES
CONFLICT OF INTERESTS
ACKNOWLEDGEMENT
REFERENCES
|
Assis IS, Chaves MB, Silveira MLL, Gern RMM, Wisbeck E, Furigo Junior A, Furlan SA (2013). Production of bioactive compounds with antitumor activity against Sarcoma 180 by Pleurotus sajor-caju. Journal of Medicinal Food 16(11):1004-1012. |
|
|
Bai J, Jia X, Chen Y, Ning Z, Liu, S, Xu C (2020).. Effect of carbon source on properties and bioactivities of exopolysaccharide produced by Trametes ochracea (Agaricomycetes). International Journal of Mushrooms 23(3):289-297. |
|
|
Borges GM, de Barba FFM, Schiebelbein AP, Pereira BP, Chaves MB, Silveira MLL, Pinho, MLS, Furlan SA, Wisbeck E (2013). Extracellular polysaccharide production by a strain of Pleurotus djamor isolated in the south of Brazil and antitumor activity on sarcoma 180. Brazilian Journal of Microbiology 44(4):1059-1065. |
|
|
Burns PJ, Yeo P, Keshavarz T, Roller S, Evans CS (1994). Physiological studies of exopolysaccharide production from basidiomycetes Pleurotus sp. Florida. Enzyme Microbiology Technology 16(7):566-572. |
|
|
Camelini CM, Rezzadori K, Benedetti S, Proner MC, Fogaça L, Azambuja AA, Giachini AJ, Rossi MJ, Petrus JCC (2013). Nanofiltration of polysaccharides from Agaricus subrufescens. Applied Microbiology and Biotechnology 97:9993-10002. |
|
|
Camelini CM, Rossi MJ. Cardozo, FTGS, Gomes A, Sales-Campos C, Giachini AJ (2014). Fungal cultivation and production of polysaccharides. In: Ramawat K, Mérillon JM (eds) Polysaccharides pp. 1-34. |
|
|
Chisti MY (1989). Airlift Bioreactors. London: Elsevier Applied Sciences 345 p. |
|
|
Confortin FG, Marchetto R, Bettin F, Camassola M, Salvador M, Dillon, AJP (2008). Production of Pleurotus sajor-caju strain PS-2001 biomass in submerged culture. Journal of Industrial Microbiology and Biotechnology 35(10):1149-1155. |
|
|
Crueger W, Crueger A (1990). Substrates for industrial fermentation. In: Biotechnology: A textbook of industrial microbiology. Wisconsin: Science Tech Publishers 59-63. |
|
|
Cui, FJ, Xu ZH, Xu HY, Sun K, Tao WY (2006). Optimization of the medium composition for production of mycelial biomass and exo-polymer by Grifola frondosa GF9801 using response surface methodology. Bioresource Technology 97(10):1209-1216. |
|
|
Duobin M, Yupping MA, Lujing G, Aijing Z, Jianqiang Z, Chunping X (2013). Fermentation characteristics in stirred-tank reactor of exopolysaccharides with hypolipidemic activity produced by Pleurotus geesteranus 5#. Anais da Academia Brasileira de Ciências 85(4):1473-1481. |
|
|
El-Enshasy H, Daba A, El Demellawy M, Ibrahim A, El Sayed S, Elabary I (2010). Bioprocess development for large scale production of anticancer exopolysaccharide by Pleurotus ostreatus in submerged culture. Journal of Applied Sciences 10(21):2523-2529. |
|
|
Elisashvili V (2012). Submerged cultivation of medicinal mushrooms: bioprocesses and products. International Journal of Medicinal Mushrooms 14(3):211-239. |
|
|
Fan L, Soccol AT, Pandey A, Soccol CR (2007). Effect of nutritional and environmental conditions on the production of exo-polysaccharide of Agaricus brasiliensis by submerged fermentation and its antitumor activity. LWT 40(1):30-35. |
|
|
Fang, QH, Zhong JJ (2002). Effect of initial pH on production of ganoderic acid and polysaccharide by submerged fermentation of Ganoderma lucidum. Process Biochemistry 37(7):769-774. |
|
|
Furnali, RPZ, Godoy HT (2007). Valor nutricional de cogumelos comestíveis. Ciencia e Tecnologia de Alimentos 27(1):154-157. |
|
|
García-Cruz F, Durán-Páramo E, Garín-Aguilar MA, del Toro GV, Chairez I (2020). Parametric characterization of the initial pH effect on the polysaccharides production by Lentinula edodes in submerged culture. Food and Bioproducts Processing 119:170-178. |
|
|
Garcia-Ochoa F, Gomez E (2009). Bioreactor scale-up and oxygen transfer rate in microbial processes: an overview. Biotechnology Advance 27(2):153-176. |
|
|
Gern RMM, Wisbeck E, Rampinelli J, Ninow JL, Furlan SA (2008). Alternative medium for production of Pleurotus ostreatus biomass and potential antitumor polysaccharides. Bioresource Technology 99(1):76-82. |
|
|
Ghorai S, Banik SP, Verma D, Chowdhury S, Mukherjee S, Khowala S |
|
|
(2009). Fungal biotechnology in food and feed processing. Food Research International 42(5-6):577-587. |
|
|
Guillén-Navarro GK, Márquez-Rocha FJ, Sanchez-Vázquez JE (1998). Producción de biomasa y enzimas ligninolíticas por Pleurotus ostreatus en cultivo sumergido. Revista Iberoamericana de Mycologia 15:302-306. |
|
|
Hamedi A, Vahid H, Ghanati F (2007) Optimization of the medium composition for production of mycelia biomass and exo- polysaccharide by Agaricus blazei Murill DPPh 131 using response surface methodology. Biotechnology 6(4):456-464. |
|
|
Hwang HJ, Kim SW, XU CP, Choi JW, Yum JW (2004). Morphological and rheological properties of the three different species of basidiomycetes Phellinus in submerged cultures. Journal of Applied Microbiology 96(6):1296-1305. |
|
|
Jong SC, Donovick R (1989). Antitumor and antiviral substances from fungi. Advances in Applied Microbiology 34:183-261. |
|
|
Joo JH, Lim JM, Kim HO, Kim SW, Hwang HJ, Choi, JW, Yun JW (2004). Optimization of submerged culture conditions for exopolysaccharide production in Sarcodon aspratus (Berk) S. Ito TG-3. World Journal of Microbiology and Biotechnology 20:767-773. |
|
|
Kavanagh K (2005). Fungal fermentation systems and products. In: Kavanagh K. Fungi: Biology and Applications. West Sussex: Wiley pp. 89-111. |
|
|
Kawagoe M, Nagaoka Y, Araki M, Yamagaki K, Naoe K, Noda H (2004) Submerged culture of Agaricus blazei mycelium in a bubble column fermentor. Journal of Chemical Engineering of Japan 37(8):1056-1061. |
|
|
Kim HO, Lim JM, Joo JH, Kim SW, Hwang HJ, Choi JW, Youn JW (2005a). Optimization of submerged culture condition for the production of mycelial biomass and exopolysaccharides by Agrocybe cylindracea. Bioresource Technology 96:1175-1182. |
|
|
Kim YO, Han SB, Lee HW, Ahn HJ, Yoon YD, Jung JK, Kim HM, Shin CS (2005b). Immuno-stimulating effect of the endo-polysaccharide produced by submerged culture of Inonotus obliquus. Life Science 77(19):2438-2456. |
|
|
Kim SW, Hwang HJ, Lee BC, Yun JW (2007). Grifola frondosa polysaccharides as cosmeceuticals, Food Technology and Biotechnology 45(3):295-305. |
|
|
Kurosumi A, Kobayasi F, Mtui G, Nakamura Y (2006). Development of optimal culture method of Sparassis crispa mycelia and a new extraction method of antineoplastic constituent. Biochemical. Engineering Journal 30(1):109-113. |
|
|
Leonowicz A, Wojias-Wasilewska M, Rogalski J, Luterek J (1991). Higher fungi as a potential feed and food source from lignocellulosic wastes. Studies in Environmetal Science 42:229-255. |
|
|
Li J, Jia Z, Yao Y, Bai Y (2019). Characterization of Exopolysaccharide Produced by Phellinus vaninii (Agaricomycetes) and Antioxidant Potential for Meat Batter. International Journal of Mushrooms 21(5):459-467. |
|
|
Liu QN, Liu RS, Wang YH, Mi ZY, Li DS, Zhong JJ (2009). Fed-batch fermentation of Tuber melanosporum for the hyperproduction of mycelia and bioactive Tuber polysaccharides. Bioresource Technology 100:3644-3649. |
|
|
Liu SR, Zhang WR (2019). Optimization of submerged culture conditions involving a developed fine powder solid seed for exopolysaccharide production by the medicinal mushroom Ganoderma lucidum. Food Science and Biotechnology 28:1135-1145. |
|
|
Maftoun P, Malek R, Mahmoud AS, Abdel-Sadek M, Aziz R, El Enshasy H (2013). Bioprocess for semi-industrial production of immunomodulator polysaccharide Pleuran by Pleurotus ostreatus in submerged culture. Journal of Scientific and Industrial Research 72(11):655-662. |
|
|
Maziero R, Cavazzoni V, Bononi VLR (1999). Screening of basidiomycetes for the production of exopolysaccharide and biomass in submerged culture. Revista de Microbiologia 30:77-84. |
|
|
Merchuk JC, Yunger R (1990). The role of the gas-liquid separator of airlift reactors in the mixing process. Chemical Engineering Science 45(9):2973-2976. |
|
|
Merchuk JC (2003). Airlift Bioreactors: Review of Recent Advances. The Canadian Journal of Chemical Engineering 81(3):324-337. |
|
|
Mohamad SA, Awang MR, Ibrahim R, Keong CY, Hamzah MY, Rashid A, Hussein S, Rahim KA, Daud F, Hamid AB, Yusoff MW (2015). Production of endopolysaccharides from Malaysia's local mushrooms in air-lift bioreactor. Advances in Bioscience and Biotechnology 6:456-462. |
|
|
Nienow AW (2014). Stirring and Stirred-Tank Reactors. Chemie Ingenieur Technik 86(12):1-13. |
|
|
Oosterhuis NMG, Neubauer P, Junne S (2013). Single-use bioreactors for microbial cultivation. Pharmeutical Bioprocess 1(2):167-177. |
|
|
Petre M, Teodorescu A, Tuluca E, Bejan C, Andronescu A (2010). Biotechnology of mushroom pellets producing by controlled submerged fermentation. Romanian Biotechnological Letters 15(2):50-55. |
|
|
Prosser JI, Tough (1991). Growth mechanisms and growth kinetics of filamentous microorganisms. Critical Reviews in Biotechnology 10(4):253-274. |
|
|
Rosado FR, Carbonero ER, Kemmelmeier C, Tischer CA, Gorin PAJ, Iacomini M (2002). A partially 3-O-methylated (14)-linked-D-galactan and alpha-D-mannan from Pleurotus ostreatoroseus Sing. FEMS Microbiology Letters 212:261-265. |
|
|
Rosado FR, Germano S, Carbonero, ER, Da Costa SM, Iacomini M, Kemmelmeier C (2003). Biomass and exopysaccharide production in submerged cultures of Pleurotus ostreatoroseus Sing. and Pleurotus ostreatus florida (Jack.:Fr.) Kummer. Journal of Basic Microbiology 43(3):230-237. |
|
|
Rossi MJ, Souza JAR, Oliveira VL (2002). Inoculum production of the ectomycorrhizal fungus Pisolithus microcarpus in an airlift bioreactor. Applied Microbiology and Biotechnology 59:175-181. |
|
|
Ruiz-Herrera J, Ortiz-Castellanos L (2019). Cell wall glucans of fungi. A review. The Cell Surface 5:100022. |
|
|
Saeki, N, Takeda, H, Tanesaka E, Yoshida M, Takeda H (2011). Induction of manganese peroxidase and laccase by Lentinula edodes under liquid culture conditions and their isozyme detection by enzymatic staining on native-PAGE. Mycoscience 52(2):132-136. |
|
|
Sales-Campos C, Vianez BF, de Abreu RLS (2011a). Productivity and nutritional composition of Lentinus strigosus (Svhwinitz) fries mushroom from the Amazon region cultivated in sawdust supplemented with soy bran. Recent Trends for Enhancing the Diversity and Quality of Soybean Products pp. 227-250. |
|
|
Sales-Campos C, Araújo LM, Minhoni MTDA, Andrade MCND (2011b). Physiochemical analysis and centesimal composition of Pleurotus ostreatus mushroom grown in residues from the Amazon. Food Science and Technology 31(2):456-461. |
|
|
Sales-Campos C, Araujo LM, Minhoni MTA, Andrade MCN (2013). Centesimal composition and physical-chemistry analysis of the edible mushroom Lentinus strigosus occurring in the Brazilian Amaz. Anais da Academia Brasileira de Ciências 85(4):1537-1544. |
|
|
Schmidell W (2001). Agitação e aeração em biorreatores. In: Schmidell W. Biotecnologia Industrial Vol II: Engenharia Bioquímica. São Paulo: Edgard Blücher Ltda. pp. 277-331. |
|
|
Seo YR, Patel DK, Shin WC, Sim WS, Lee OH, Lim KT (2019). Structural Elucidation and Immune-Enhancing Effects of Novel Polysaccharide from Grifola frondosa. BioMed Research International 2019:1-7. |
|
|
Shu CH, Lung MY (2004). Effect of pH on the production and molecular weight distribution of exopolysaccharide by Antrodia camphorata in batch cultures. Process Biochemistry 39(8):931-937. |
|
|
Silveira MLL, Smiderle FR, Agostini F, Pereira EM, Bonatti-Chavesa M, Wisbeck E, Ruthes AC, Sassaki GL, Cipriani TR, Furlan, AS, Iacomini M (2015). Exopolysaccharide produced by Pleurotus sajor-caju: Its chemical structure and anti-inflammatory activity. International Journal of Biological Macromolecules 75:90-96. |
|
|
Singh MP, Vishwakarma SK, Srivastava (2013). Bioremediation of direct blue 14 and extracellular ligninolytic enzyme production by white rot fungi: Pleurotus spp. BioMed Research International 2013:1-4. |
|
|
Smith JE, Rowan NJ, Sullivan R (2002). Medicinal mushrooms: a rapidly developing area of biotechnology for cancer therapy and other bioactivities. Biotechnology Letters 24:1839-1845. |
|
|
Tang YJ, Zhong JJ (2002). Fed-batch fermentation of Ganoderma lucidum for hyperproduction of polysaccharides and ganoderic acid. Enzyme and Microbial Technology 31(1-2):20-28. |
|
|
Tang YJ, Zhu LW, Li HM, Li DS (2007). Submerged culture of mushrooms in bioreactors - Challenges, current state-of-the-art, and future prospects. Food Technology and Biotechnology 45(3):221-229. |
|
|
Tang YJ, Zhu LL, Liu RS, Li HM, Li DS, Mi ZY (2008). Quantitative response of cell growth and Tuber polysaccharides biosynthesis by medicinal mushroom chinese truffle Tuber sinense to metal ion in culture medium Bioresource Technology 99(16):7606-7615. |
|
|
Vamanu E (2012). Biological activities of the polysaccharides produced in submerged culture of two edible Pleurotus ostreatus mushrooms. Journal of Biomedicine and Biotechnology 2012:56597. |
|
|
Wang T, He H, Liu X, Liu C, Liag Y, Mei Y (2019). Mycelial polysaccharides of Lentinus edodes (shiitake mushroom) in submerged culture exert immunoenhancing effect on macrophage cells via MAPK pathway. International Journal of Biological Macromolecules 130:745-754. |
|
|
Wu JZ, Cheung PCK, Wong K-H, Huang N-L (2003) Studies on submerged fermentation of Pleurotus tuber-regium (Fr.) Singer. Part 1: physical and chemical factors affecting the rate of mycelial growth and bioconversion efficiency. Food Chemistry 81(3):389-393. |
|
|
Wu JZ, Cheung PCK, Wong KH, Huang N-L (2004) Studies on submerged fermentation of Pleurotus tuber-regium (Fr.) Singer. Part 2: effect of carbon-to-nitrogen ratio of the culture medium on the content and composition of the mycelial dietary fibre. Food Chemistry 85(1):101-105. |
|
|
Xiao JH, Chen DX, Wan WH, Hu XJ, Qi Y, Liang ZQ (2006). Enhanced simultaneous production of mycelia and intracellular polysaccharide in submerged cultivation of Cordyceps jiangxiensis using desirability functions. Process Biochemistry 41(8):1887-1893. |
|
|
Xu CP, Yun JW (2003). Optimization of submerged-culture conditions for mycelial growth and exo-biopolymer production by Auricularia polytricha (wood ears fungus) using the methods of uniform design and regression analysis. Biotechnology and Applied Biochemistry. 38(2):193-199. |
|
|
Xu C, Geng L, Zhang W (2013). Production of Extracellular Polysaccharides by the Medicinal Mushroom Trametes trogii (Higher Basidiomycetes) in Stirred-Tank and Airlift Reactors. International Journal of Medicinal Mushrooms 15(2):183-189. |
|
|
Znad HV, Báleš VJ, Markoš J, Kawaseb Y (2004). Modeling and simulation of airlift bioreactors. Biochemical Engineering Journal 21(1):73-81. |
|
|
Zhang M, Zhang L, Cheung PC (2003). Molecular mass and chain conformation of carboxymethylate derivatives of β-glucan from sclerotia of Pleurotus tuber-regium. Biopolymers 68(2):150-159. |
|
|
Zhang Z, Guoying LV, Pan H, dos Santos LF, Fan M, Soccol CR, Soccol, VT, Lacerda LG, Carvalho Filho MAS, Fan L (2011). Antidiabetic activities of ethanol extract of dry matters of culture broth of Coriolus versiolor in submerged culture. Brazilian Archives of Biology and Technology 54(4):701-708. |
|
|
Zou X (2006). Fed-batch fermentation for hyperproduction of polysaccharide and ergosterol by medicinal mushroom Agaricus brasiliensis, Process Biochemistry 41(4):970-974. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


