Review

ABSTRACT
Detection and identification of plant pathogens is one of the most important strategies for sustainable plant diseases management. For this reason, the availability of fast, sensitive and accurate methods for detection and identification of plant pathogens is increasingly necessary to improve disease control decision making process. In other words, new technologies and improved methods with reduced/fair cost and improved speed, throughput, multiplexing, accuracy and sensitivity have emerged as an essential strategy for the control of both fungal and bacterial diseases. The development of recombinant DNA technology is also possible to isolate individual genes and incorporate resistance genes into otherwise agronomically acceptable cultivars to develop genetically resistance variety for a particular disease. These advances have been complemented by the development of new nucleic acids extraction methods, increased automation, reliable internal controls, multiplexing assays, online information and on site molecular diagnostics. The different types of polymerase chain reaction (PCR) are the most common DNA amplification technology used for detecting various plant pathogens. With the applications of bioinformatics as a modern technology in plant pathology, identification of specific motifs, DNA sequences has become possible, which ultimately increase the accuracy of modern techniques in plant disease diagnosis. The newly emerged proteomic technology is also a promising tool for providing information about pathogenicity and virulence factors that will open up new possibilities for plant disease diagnosis and appropriate protection measures.
Key words: Biotechnology, molecular markers, marker assisted selection, quantitative trait loci (QTL), polymerase chain reaction (PCR), proteomics.
INTRODUCTION
ADVANCED TOOLS FOR DETECTION AND IDENTIFICATION OF PATHOGENS
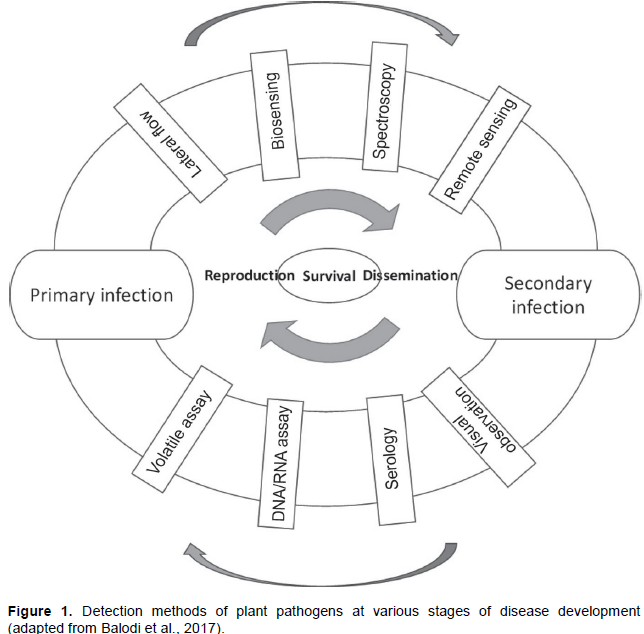
APPLICATION OF MOLECULAR MARKERS FOR RESISTANCE BREEDING
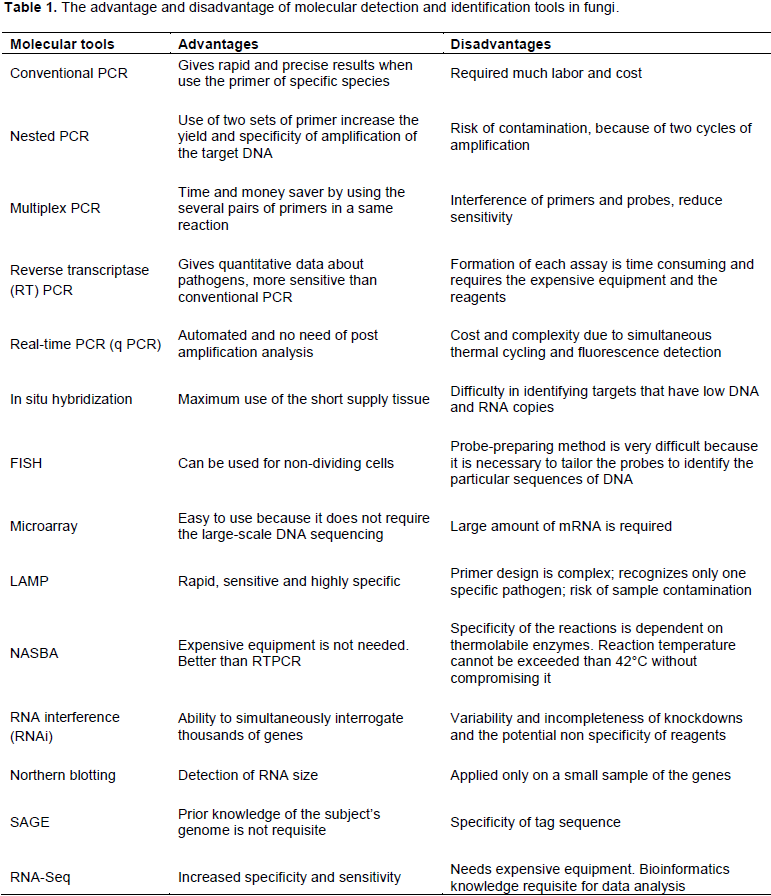
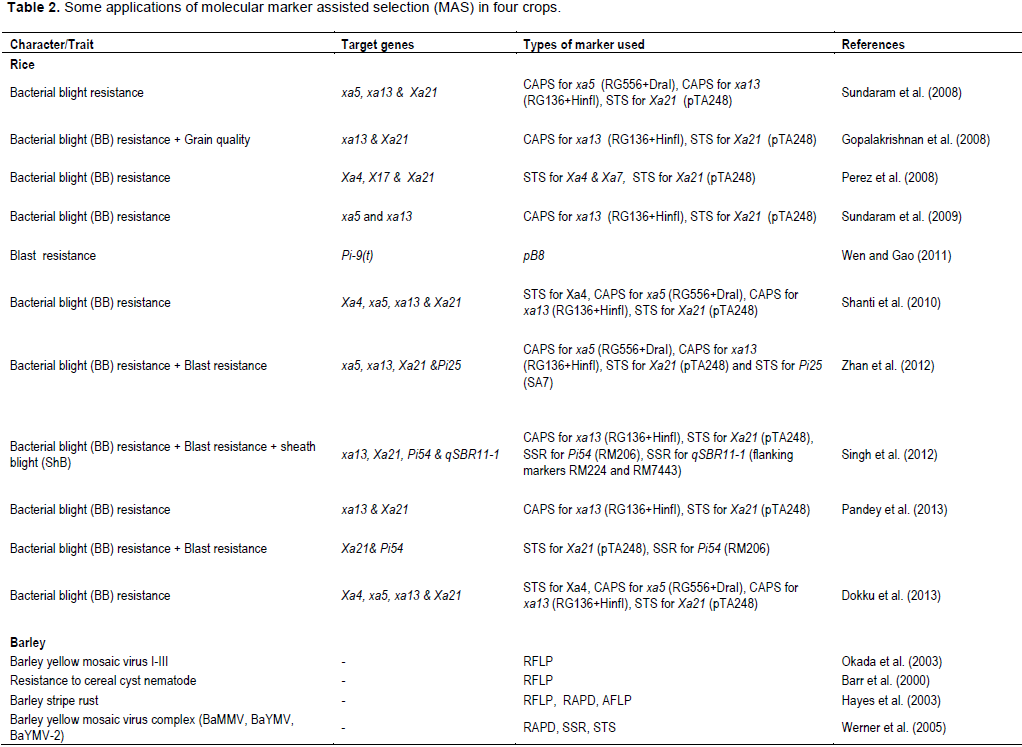
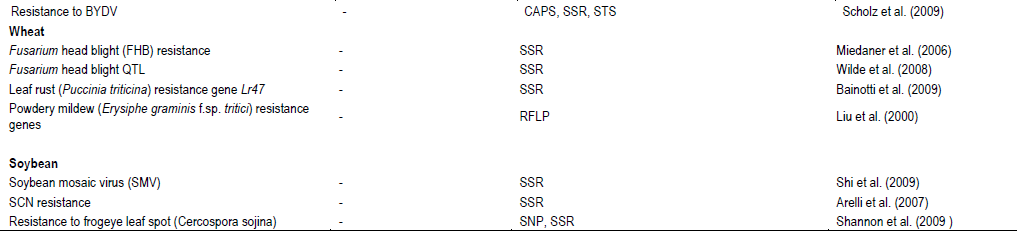
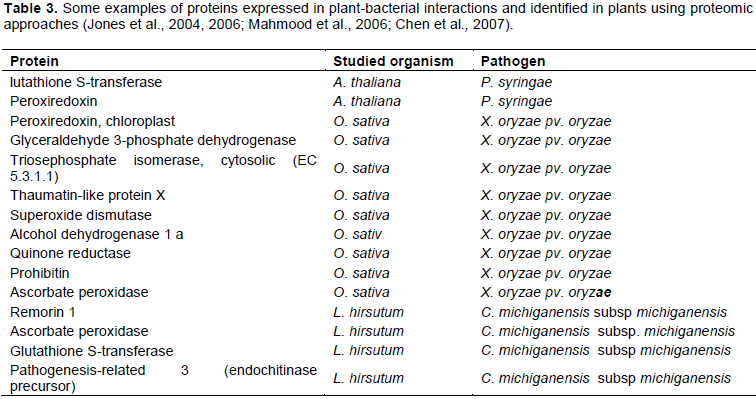
CONCLUSIONS
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENT
The authors would like to thank the staff members of Plant Protection Department, Selcuk University, Turkey for giving moral to develop this manuscript and helpful review.
REFERENCES
|
Agrios GN (2005). Plant Pathology. 5 th Ed., Elsevier Academic Press. New York pp. 213-223 |
|
|
Alemu K (2014). Real-time PCR and its application in plant disease diagnostics. Advances in Life Science and Technology 27:29-49. |
|
|
Alvarez E, Llano G, Mejia J (2008). Applying biotechnology tools to improve control diseases of some tropical crops. Ftopatologia Colombiana 28(2):93-97. |
|
|
Arelli PR, Pantalone VR, Allen FL, Mengistu A (2007). Registration of soybean germplasm JTN-5303. Journal Plant Registrations 1:69-70. |
|
|
Aslam S, Tahir A, Aslam MF, Alam MW, Shedayi AA, Sadia S (2017). Recent advances in molecular techniques for the identification of phytopathogenic fungi - a mini review. Journal of Plant Interactions 12(1):493-504. |
|
|
Audy P, Laroche A, Saindon G, Huang HC, Gilbertson RC (1994). Detection of bean common blight bacteria Xanthomonas campestris pv. phaseoli and X. campestris pv. phaseoli var. fuscans using polymerase chain reaction. Phytopathology 84:1185-1192. |
|
|
Badali H, Nabili M (2012). Molecular tools in medical mycology; where we are! Jundishapur Journal of Microbiology 6(1):1-3. |
|
|
Bainotti C, Fraschina J, Salines JH, Nisi JE, Dubcovsky J, Lewis SM, Bullrich L, Vanzetti L, Cuniberti M, Campos P, Formica MB, Masiero B, Alberione E, Helguera M (2009). Registration of 'BIOINTA 2004 wheat. Journal of Plant Registrations 3:165-169. |
|
|
Balodi R, Bisht S, Ghatak A, Rao KH (2017). Plant disease diagnosis: technological advancements and challenges. Indian Phytopathology 70(3):275-281. |
|
|
Barnes CW, Szabo LJ (2007). Detection and identification of four common rust pathogens of cereals and grasses using real-time polymerase chain reaction. Phytopathology 97(6):717-727. |
|
|
Barr AR, Jefferies SP, Warner P, Moody DB, Chalmers KJ, Langridge P (2000). Marker- assisted selection in theory and practice. Prodeedings of the 8th International Barley Genetics Symposium, Vol I., Adelaide, Australia pp. 167-178. |
|
|
Billard A, Laval V, Fillinger S, Leroux P, Lachaise H, Beffa R, Debiue D (2012). The allele specific probe and primer amplification assay, a new real time PCR method for fine quantification of single nucleotide Polymorphism in pooled DNA. Applied and Environmental Microbiolgy 78(04):1063-1068. |
|
|
Boora KS, Frederiksen RA, Magill CW (1999). A molecular marker that segregates with sorghum leaf blight resistance in one cross is maternally inherited in another. Molecular and General Genetics 261:317-322. |
|
|
Capote N, Pastrana AM, Aguado A, Sánchez-Torres P (2012). Molecular tools for detection of plant pathogenic fungi and fungicide resistance. In Cumagun CJ (Ed.), Plant Pathology, InTech, Rijeka, Croatia 362 p. |
|
|
Chen RS, Chu C, Cheng CW, Chen WY, Tsay JG (2008). Differentiation of two powdery mildews of sunflower (Helianthus annuus) by a PCR-mediated method based on ITS sequences. European Journal of Plant Pathology 121(1):1-8. |
|
|
Chen F, Yuan Y, Li Q, He Z (2007). Proteomic analysis of rice plasma membrane reveals proteins involved in early defense response to bacterial blight. Proteomics 7:1529-1539. |
|
|
Dokku P, Das KM, Rao GJN (2013). Pyramiding of four resistance genes of bacterial blight in Tapaswini, an elite rice cultivar, through marker-assisted selection. Euphytica 192:87-96. |
|
|
Doveri S, Maheswaran M, Powell W (2008). Molecular markers - history, features and applications. In: Kole C, Abbott AG (eds) Principles and Practices of Plant Genomics. Vol. 1: Genome Mapping. Science Publishers, Enfield, Jersey, Plymouth, England. |
|
|
Fagwalawa LD, Kutama AS, Yakasai MT (2013). Current issues in plant disease control: Biotechnology and plant disease. Bayero Journal of Pure and Applied Sciences 6(2):121-12. |
|
|
Gopalakrishnan S, Sharma RK, Rajkumar KA, Joseph M, Singh VP, Singh AK, Bhat KV, Singh NK, Mohapatra T (2008). Integrating marker assisted background analysis with foreground selection for identification of superior bacterial blight resistant recombinants in Basmati rice. Plant Breeding 127:131-139. |
|
|
Guglielmo F, Bergemann SE, Gonthier P, Nicolotti G, Garbelotto M (2007). A multiplex PCR-based method for the detection and early identification of wood rotting fungi in standing trees. Journal of Applied Microbiology 103:1490-1507. |
|
|
Haggag WM (2008). Biotechnological aspects of plant resistant for fungal diseases management. American-Eurasian Journal of Sustainable Agriculture 2(1):1-18. |
|
|
Hayes PM, Corey AE, Mundt C, Toojinda T, Vivar H (2003). Registration of 'Tango' Barley. Crop Science 43:729-731. |
|
|
James C (2003). Preview: Global status of commercialized transgenic crops: 2003. ISAAA Briefs No. 30. Ithaca, NY. |
|
|
Jeeva ML, Mishra AK, Vidyadharan P, Misra RS, Hegde V (2010). A species-specific polymerase chain reaction assay for rapid and sensitive detection of Sclerotium rolfsii. Australasian Plant Pathology 39(6):517-523. |
|
|
Jones AME, Thomas V, Truman B, Lilley K, Mansfield J, Grant M (2004). Specific changes in the Arabidopsis proteome in response to bacterial challenge: differentiating basal and R-gene mediated resistance. Phytochemistry 65:1805-1816. |
|
|
Jones AME, Thomas V, Bennett MH, Mansfield J, Grant M (2006). Modifications to the Arabidopsis defense proteome occur prior to significant transcriptional change in response to inoculation with Pseudomonas syringae. Plant Physiology 142:1603-1620. |
|
|
Kang S, Mansfield MA, Park B, Geiser DM, Ivors KL, Coffey MD, Grünwald NJ, Martin FN, Lévesque CA, Blair JE (2010). The promise and pitfalls of sequence-based identification of plant-pathogenic fungi and oomycetes. Phytopathology 100(8):732-737. |
|
|
Kaymak S, Boyraz N, Daniels J (2016). Molecular markers to evaluate genetic diversity among Venturia inaequalis isolates obtained from apple plantations in Isparta Province. Turkish Journal of Agriculture and Forestry 40:489-498. |
|
|
Liu J, Liu D, Tao W, Li W, Wang CP, Cheng S, Gao D (2000). Molecular marker-facilitated pyramiding of different genes for powdery mildew resistance in wheat. Plant Breeding 119:21-24. |
|
|
Lodha TD, Hembram P, Tep N, Basak J (2013). Proteomics: A successful approach to understand the molecular mechanism of plant-pathogen interaction. American Journal of Plant Sciences 4:1212-1226. |
|
|
Mahmood T, Jan A, Kakishima M, Komatsu S (2006). Proteomic analysis of bacterial-blight defenceresponsive proteins in rice leaf blades. Proteomics 6:6053-6065. |
|
|
Manulis S, Kogan N, Reuven M, Yephet YB (1994). Use of the RAPD technique for identification of Fusarium oxysporum f. sp. dianthi from carnation. Phytopathol 84:98-101. |
|
|
Martin RR, Constable F, Tzanetakis IE (2016). Quarantine regulations and the impact of modern detection methods. Annual Review of Phytopathology 54:189-205. |
|
|
Martins D, Astua-Monge G, Coletta-Filho HD, Winck FV, Baldasso PA, de Oliveira BM, Marangoni S, Machado MA, Novello JC, Smolka MB (2007). Absence of classical heat shock response in the citrus pathogen Xylella fastidiosa. Current Microbiology 54:119-123. |
|
|
Mathesius U, Mulders S, Gao M, Teplitski M, Caetano-Anolles G, Rolfe BG, Bauer WD (2003). Extensive and specific responses of a eukaryote to bacterial quorum-sensing signals. Proceedings of the National Academy of Sciences USA 100:1444-1449. |
|
|
McCartney HA, Foster SJ, Fraaije BA, Ward E (2003). Molecular diagnostics for fungal plant pathogens. Pest Management Science 59(2):129-142. |
|
|
Mehta A, Rosato YB (2001). Differentially expressed proteins in the interaction of Xanthomonas axonopodis pv. citri with leaf extract of the host plant. Proteomics 1:1111-1118. |
|
|
Miedaner T, Wilde F, Steiner B, Buerstmayr H, Korzun V, Ebmeyer E (2006). Stacking quantitative trait loci (QTL) for fusarium head blight resistance from non-adapted sources in a European elite spring wheat background and assessing their effects on deoxynivalenol (DON content and disease severity. Theoretical and Applied Genetics 112:562-569. |
|
|
Mittal M, Boora KS (2005). Molecular tagging of gene conferring leaf blight resistance using microsatellites in sorghum {Sorghum bicolour (L.) Moench}. Indian Journal of Experimental Biology 43(5):462-466. |
|
|
Narayanasamy P (2011). Microbial plant pathogens-detection and disease diagnosis, Springer Dordrecht Heidelberg London New York, 2:280. |
|
|
Nilsson RH, Abarenkov K, Larsson KH, Kõljalg U (2011). Molecular identification of fungi: rationale, philosophical concerns, and the UNITE database. The Open Applied Informatics Journal 5:81-86. |
|
|
Okada Y, Kanatani R, Arai S, Asakura T, Ito K (2003). Production of a novel virus-resistant barley line introgression to the rym1 locus with high malting quality using DNA marker assisted selection. Journal of the Institute of Brewing 109:99-102. |
|
|
Pandey MK, Shobha Rani N, Sundaram RM, Laha GS, Madhav MS, Srinivasa Rao K, Injey Sudharshan, Yadla Hari, Varaprasad GS, Subba Rao LV, Kota Suneetha, Sivaranjani AKP, Viraktamath BC (2013). Improvement of two traditional Basmati rice varieties for bacterial blight resistance and plant stature through morphological and marker-assisted selection. Molecular Breeding 31:239-246. |
|
|
Perez DR, Lopez GE, Rivera J, Ferreira A, Fontan G (2008). Naturally occurring Bruton's tyrosine kinase mutations have no dominant negative effect in an Xlinked agammaglobulinaemia cellular model. Clinical and Experimental Immunology 152:33-38. |
|
|
Punja ZK, De Boer SH, Sanfaçon H (2008). Biotechnology and plant disease management. CAB International, Canada 590 p. |
|
|
Ragimekula N, Varadarajula NN, Mallapuram SP, Gangimeni G, Reddy RK, Kondreddy HR (2013). Marker assisted selection in disease resistance breeding. Journal of Plant Breeding and Genetics 1(2):90-109. |
|
|
Scholz M, Ruge-Wehling B, Habekub A, Schrader O, Pendinen G, Fischer K, Wehling P (2009). Ryd4Hb: A novel resistance gene introgressed from Hordeum bulbosum into barley and conferring complete and dominant resistance to the barley yellow dwarf virus. Theoretical and Applied Genetics 119:837-849. |
|
|
Shamim M, Kumar P, Kumar RR, Kumar M, Kumar RR, Singh KN (2017). Assessing fungal biodiversity using molecular markers. In: Singh B., Gupta V. (eds) Molecular markers in mycology. Fungal biology. Springer, Cham, Switzerland. |
|
|
Shannon JG, Lee JD, Wrather JA, Sleper DA, Mian MAR, Bond JP, Robbins RT (2009). Registration of S99-2281 soybean germplasm line with resistance to frogeye leaf spot and three nematode species. Journal of Plant Registrations 3:94-98. |
|
|
Shanti ML, Shenoy VV, Lalitha Devi G, Mohan Kumar V, Premalatha P, Naveen Kumar G, Shashidhar HE, Zehr UB, Freeman WH (2010). Marker-assisted breeding for resistance to bacterial leaf blight in popular cultivar and parental lines of hybrid rice. Journal of Plant Pathology 92(2):495-501. |
|
|
Shi A, Chen P, Li D, Zheng C, Zhang B, Hou A (2009). Pyramiding multiple genes for resistance to soybean. Mosaic virus using molecular markers. Molecular Breeding 23:113-124. |
|
|
Singh A, Singh KV, Singh SP, Pandian RTP, Ranjith EK, Singh D, Bhowmick KP, Gopala Krishnan S, Nagarajan M, Vinod KK, Singh UD, Prabhu KV, Sharma TR, Mohapatra T, Singh AK (2012). Molecular breeding for the development of multiple disease resistance in Basmati rice. |
|
|
Singh Y, Singh J, d Pandey AK (2013). Molecular markers in diagnosis and management of fungal pathogens: A Review. International Journal of Advanced Biotechnology and Research 4(2):180-188. |
|
|
Sundaram RM, Vishnupriya MR, Biradar SK, Laha GS, Reddy GA, Shoba Rani N, Sarma NP, Sonti RV (2008). Marker assisted introgression of bacterial blight resistance in Samba Mahsuri, an elite indica rice variety. Euphytica 160:411-422. |
|
|
Sundaram RM, Priya MRV, Laha GS, Shobha Rani N, Srinivasa Rao P, Balachandran SM, Ashok Reddy G, Sarma NP, Sonti RV (2009). Introduction of bacterial blight resistance into Triguna, a high yielding, mid-early duration rice variety by molecular marker assisted breeding. Biotechnology Journal 4:400-407. |
|
|
Suarez -Moreno ZR, Caballero-Mellado J, Coutinho BG, Mendonça-Previato L, James EK, Venturi V (2012). Common features of environmental and potentially beneficial plant-associated Burkholderia. Microbial Ecology 63:249-66. |
|
|
Tahara ST, Mehta A, Rosato YB (2003). Proteins induced by Xanthomonas axonopodis pv. passiflorae with leaf extract of the host plant (Passiflorae edulis). Proteomics 3:95-102. |
|
|
Torres AM (2010). Application of molecular markers for breeding disease resistant varieties in crop plants. In Molecular techniques in crop improvement. Springer, Dordrecht pp. 185-205. |
|
|
Torres-Calzada C, Tapia-Tussell R, Quijano-Ramayo A, Martin-Mex R, Rojas-Herrera R, Higuera-Ciapara I, Perez-Brito D (2011). A species-specific polymerase chain reaction assay for rapid and sensitive detection of Colletotrichum capsici. Molecular Biotechnology 49(1):48-55. |
|
|
Van Doorn R, Slawiak M, Szemes M, Dullemans AM, Bonants P, Kowalchuk GA, Schoen CD (2009). Robust detection and identification of multiple oomycetes and fungi in environmental samples by using a novel cleavable padlock probe-based ligation detection assay. Applied and Environmental Microbiology 75(12):4185-4193. |
|
|
Wen S, Gao B (2011). Introgressing blast resistant gene Pi-9(t) into elite rice restorer luhui17 by marker-assisted selection. Rice Genomics and Genetics 2(4):31-36. |
|
|
Werner K, Friedt W, Ordon F (2005). Strategies for pyramiding resistance genes against the barley yellow mosaic virus complex (BaMMV, BaYMV, BaYMV-2). Molecular Breeding 16:45-55. |
|
|
Wilde F, Schon C, Korzun V, Ebmeyer E, Schmolke M, Hartl L, Miedaner T (2008). Marker-based introduction of three quantitative-trait loci conferring resistance to fusarium head blight into an independent elite winter wheat breeding population. Theoretical and Applied Genetics 117:29-35. |
|
|
Zhan XD, Zhou HP, Chai RY, Zhuang JY, Cheng SH, Cao LY (2012). Breeding of R8012, a rice restorer line resistant to blast and bacterial blight through marker-assisted selection. Rice Science 19 (1):29-35. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


