The elephant grass has gained prominence as one of the main forage species used for biomass production. Therefore, the aim of this study was to identify elephant grass genotypes with high energy biomass production potential by evaluating morpho-agronomic and biomass quality. The following traits were evaluated in this study: dry matter yield (DMY), percentage of whole-plant dry matter (%DM), percentage of neutral detergent fiber (%NDF), percentage of acid detergent fiber (%ADF); percentage of cellulose (%CEL), percentage of lignin (%LIG), percentage of carbon (%C), percentage of nitrogen (%N), and carbon: nitrogen ratio (C: N). Five different production ages were evaluated, and significant differences were observed for the variable DMY. The harvests performed at 20 and 24 weeks of age, provided the best response for biomass energy production (DMY) from elephant grass, averaging 20.50 and 23.77 t.ha−1. The genotypes that most stood out during the evaluation period at the five production ages were King Grass, Mole de Volta Grande, and Mercker 86 - México. Genotypes Mole de Volta Grande and King Grass are the most suitable for elephant grass breeding programs aimed at biomass energy production in the conditions of Campos dos Goytacazes - RJ, Brazil.
The increasing search for alternatives to fossil fuels has become a critical issue for the future of the economic development of the planet. Amidst the possible solutions, biomass stands out as a medium- and long-term choice (Goldemberg, 2009). Because biomass burning only recycles the CO2 taken from the atmosphere by photosynthesis, it appears that, in the long term, this will be one of the safest energy alternatives, provided that it is produced efficiently (Kalt and Kranzl, 2011).
The elephant grass (Pennisetum purpureum Schum.) is triploid tropical forage of African origin, which presents high production capacity and quality dry matter accumulation. In this regard, elephant grass has gained prominence as one of the main forage species used in energy production (Morais et al., 2011).
According to Mazzarella (2006), the comparative advantages of elephant grass for biomass production in relation to other sources include: greater yield (around 45 t/DM/ha/year), shorter production time, better cash flow, possibility of total mechanization, renewable energy, and greater carbon assimilation. Since its products are not directly present in the human diet and because it is usable in its entirety, elephant grass is ahead of other grasses used as energy source.
Elephant grass species feature is a large genetic variability (Meinerz et al., 2011). For this reason, materials adapted to the different ecosystems of Brazil should be selected to broaden the understanding of the interrelationships among traits. As a result, the employment of elephant grass as a bioenergy source can be improved and this plant can be elevated to a prominent level in the sustainable diversification of the Brazilian energy matrix. Therefore, aiming to investigate and understand the mechanisms to qualitatively and quantitatively increase the production of elephant grass for biomass energy generation, this study identified elephant grass genotypes with high biomass energy production by evaluating morpho-agronomic and biomass quality.
This experiment was implemented on April 26, 2010, at the State Center for Research in Agro-Energy and Waste Utilization (PESAGRO-Rio), located in Campos dos Goytacazes, Northern Rio de Janeiro State, Brazil. The climate in the region is a tropical hot and humid Aw type, according to the Köppen (1948) classification, with dry winters and rainy summers.
The soil in the experimental area is classified as a Typic Dystrophic Yellow Latosol, according to the Brazilian soil taxonomy. Chemical analysis revealed the following soil composition: pH - 6.3; phosphorus - 5.0 mg/dm3; potassium - 176 mg/dm3; calcium - 2.6 cmolc/dm3; magnesium - 1.4 cmolc/dm3; aluminum - 0.1 cmolc/dm3; hydrogen + aluminum - 0.0 cmolc/dm3 and carbon - 1.26%. Total monthly precipitation rates recorded during the experimental period are presented in Table 1. Six of the most productive genotypes of elephant grass, with the best biomass quality, were selected according to results obtained by Rossi (2010).
The experiment was undertaken in a randomized block statistical design with three replicates in an arrangement of plots subdivided into two factors: plots (genotypes) and subplots (number of harvests). Each plot consisted of 15-m row with 1-m spacing, and each subplot was composed of 3 m, in which only the two central meters of the row were considered for evaluation, disregarding half a meter from the borders of each row.
After the crop establishment phase in the field, the plot was leveled-off on 08/05/2010, followed by topdressing with 30 kg.ha−1 ammonium sulfate and 20 kg.ha−1 potassium chloride. Seedlings were re-planted to recompose the plots, followed by topdressing on 10/31/2010. After this process and the reestablishment of the crop in the field, a second plot-leveling cut was made on December 10, 2010. After that date, harvests were performed in plots chosen at random 8, 12, 16, 20, and 24 weeks of age, on 02/02/2011, 03/02/2011, 04/05/2011, 04/26/2011, and 05/23/2011, respectively.
Before the harvest, the traits were assessed in samples of whole plants collected at random. Prior to being analyzed and evaluated in the laboratory, these samples were dried in a forced-air oven at 65°C for 72 h. After drying, samples were ground through a Wiley mill with 1-mm sieves and packed in glass bottles.
Plant dry matter yield in t.ha−1 (DMY), was estimated as the product between whole-plant fresh matter yield and the percentage of whole-plant dry matter; the obtained value was then converted to t.ha−1.
The following biomass quality-related traits were evaluated: percentage of neutral detergent fiber (% NDF); percentage of acid detergent fiber (% ADF); percentage of cellulose (% CEL); and percentage of lignin (% LIG), were carried out at the Laboratory of Food Analysis of Embrapa Gado de Leite, in Juiz de Fora, MG, Brazil, by the method of near infrared reflectance (NIRS) (Van Soest, 1963), in a Perstorp analytical spectrometer, Silver Spring, MD, model 5000, coupled to a microcomputer equipped with ISI software version 4.1 (Infrasoft International, University, Park, PA). Preliminary evaluations were made for the calibration of the equipment using samples referring to the different cutting ages, and the final reading was performed using the wavelengths of 1100 to 2500 nanometers. Percentages of carbon (%C) and nitrogen (%N) and carbon:nitrogen (C:N) ratio were obtained using the CHNS/O Perkin Elmer (14.800) auto-analyzer.
Statistical analyses were performed using the GENES software (Computer Software for Genetics and Statistics) (Cruz, 2016), developed by the Federal University of Viçosa.
Variance analysis by production age
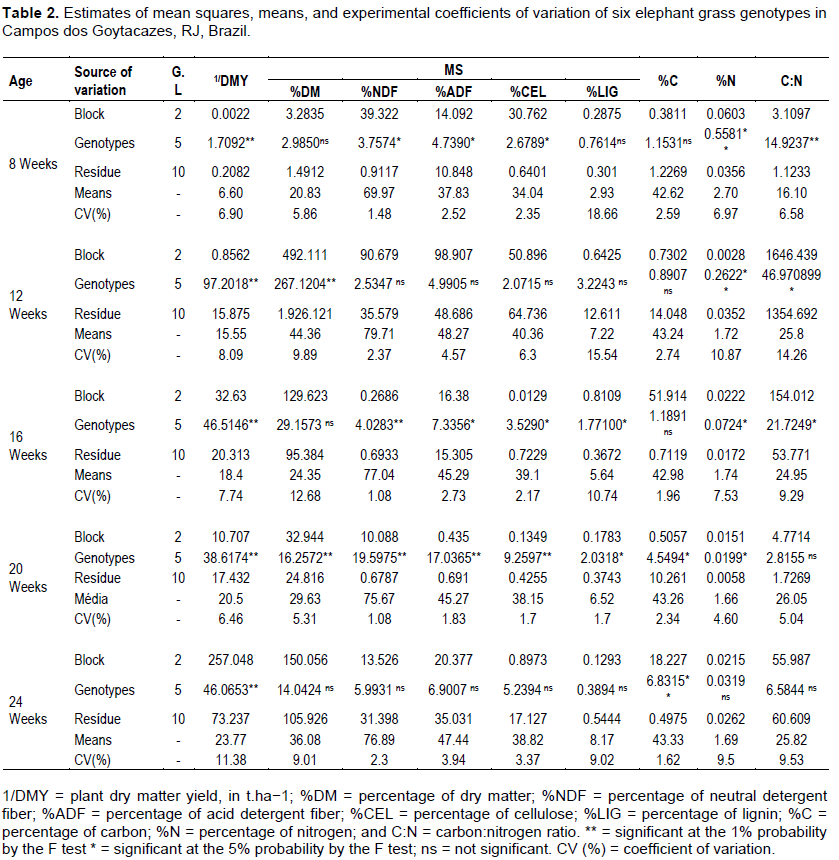
The variance analyses (P<0.01 and P<0.05) for the ages of the six genotypes evaluated (Table 2) revealed significant differences for almost all evaluated traits. This fact can be regarded as indicative of the phenotypic expression of the existing genetic variability, in the species and in the genotype collection evaluated (Pereira et al., 2008).
For the first production age with plants harvested at eight weeks, there were significant differences for the source of variation treatment (genotypes) and for most of the evaluated traits, except percentages of dry matter (%DM), %LIG, and %C. These findings confirm the existence of genetic variability between the treatments evaluated at this production age.
In the harvest performed at 12 weeks of age, there was an increase in relation to the previous harvest for the traits %DM, %LIG, and C: N, and also DMY, the trait of greatest importance for elephant grass, which averaged 6.60 and 15.55 t.ha−1 in the first and second harvests, respectively. Similar dry matter yields were obtained by Silva et al. (2014), who evaluated elephant grass hybrids with high production potential in Campos dos Goytacazes - RJ, Brazil.
These results can be explained by the climatic conditions of the period with highest incidence of rainfall (20.51 to 212.68 mm) (Table 1), followed by the topdressing applied in the restoration of the plots, which might have provided a greater development of the genotypes.
This result proves the existing difference between harvests performed in different periods, indicating that plants may undergo changes in their structure and morphology due to adverse environmental conditions. According to Faria et al. (2009), the genotype × environment interaction is of great importance in plant breeding; however, it compromises the identification of genotypes superior for different environments. At the harvest performed at 16 weeks of age, there were significant differences between genotypes for the majority of the evaluated traits, except %DM and %C.
For the fourth production age, at 20 weeks, there were significant differences between genotypes for most of the evaluated traits, except C: N ratio. The C: N ratio values found here were lower than the 80 to 156 obtained by Morais et al. (2009); the highest C: N ratio found in the present study was 26.05. This low ratio was due to the favorable climatic conditions for elephant grass growing (Flores et al., 2013).
At the fifth (and last) harvest performed at 24 weeks of age, the %DM, %NDF, %ADF, %CEL, %LIG, %N, and C:N traits showed a non-significant effect, demonstrating that the constitution of the walls of the tissues are similar across the genotypes.
In summary, at all five production ages, significant differences were observed for the variable DMY. Evaluating harvests individually by variance analysis, we observe that the fourth and fifth production ages (20 and 24 weeks of age) yielded the best responses for elephant grass biomass production (DMY), which averaged 20.50 and 23.77 t.ha−1.
These results are similar to those found by Morais et al. (2009), who evaluated the production potential of different elephant-grass genotypes at different production ages (9, 18, and 24 weeks) and observed an average DMY of 29.5 t.ha−1. Our results are also similar to the 11.15 to 23.08 t.ha−1 found by Rocha et al. (2015), who evaluated 73 elephant grass genotypes in harvests performed at 24 weeks.
This demonstrates that elephant grass can be harvested more than two times per year, as typical, for biomass production as long as ideal conditions of planting and preservation of the crop in the field are provided.
Comparison of genotype means by production age (Tukey’s test)
Based on Tukey’s mean comparison test at the 5% probability level (Table 3), the estimated means for the traits evaluated in six genotypes at five different production ages showed that, at the first production age for the trait DMY, the best response was observed for genotype Pusa Napier no. 1, whose average dry matter yield was 7.27 t.ha−1. For NDF, ADF, CEL, and %LIG traits, genotype Mole de Volta Grande had a better response than the others. At 12 weeks of age, King Grass was the genotype that most stood out for average DMY, %NDF, %ADF, %CEL, and %LIG, revealing 26.54 t.ha−1, 80.03%, 50.44%, 40.95%, and 8.80%, respectively. For the C: N trait, genotype Mercker 86 - México showed the best response: 31.25.
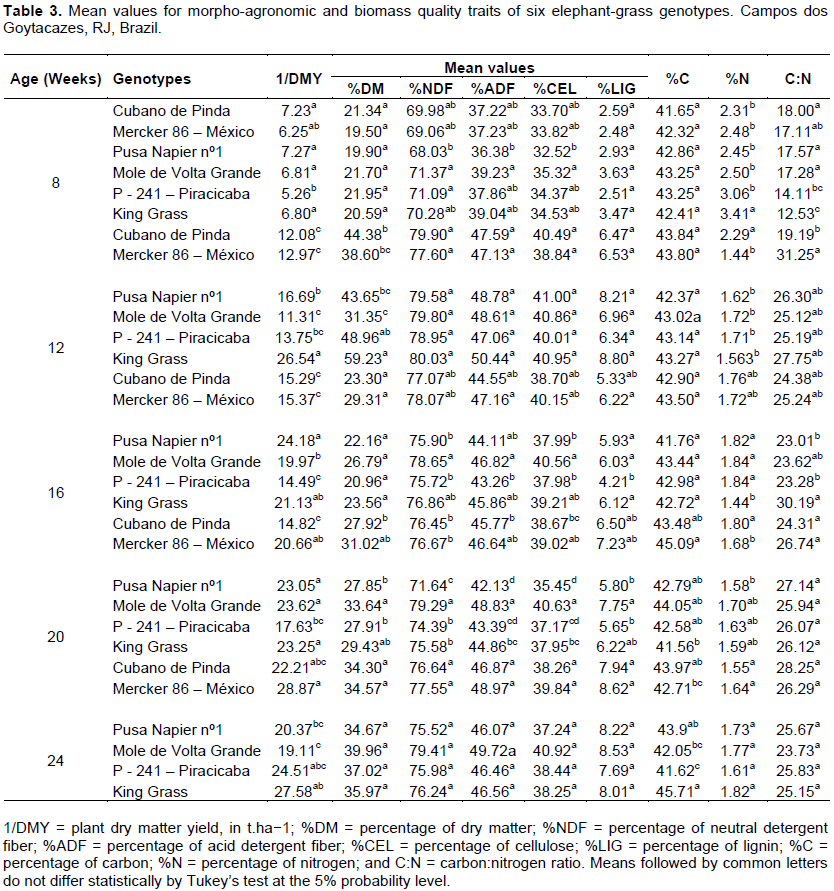
As in the first harvest, in the harvest performed at 16 weeks of age, genotype Pusa Napier no. 1 had the best average production, 24.18 t.ha−1. For the other traits of high importance for energy production, such as NDF and %CEL, genotype Mole de Volta Grande obtained 78.65% NDF and 40.56% CEL, respectively. As for the %ADF and %LIG traits, genotype Mercker 86-México averaged 47.16 and 6.22%, respectively. At the harvest performed at 20 weeks of age, genotype Mole de Volta Grande showed the best response, with a DMY of 23.62 t.ha−1, 79.29% NDF, and 48.83% ADF. Its cellulose percentage was 40.63%, corroborating Kannika et al. (2011), who reported an increase in %CEL as plants grew older.
At the fifth and last production harvest, which was performed at 24 weeks of age, the genotype that most stood out for DMY and %LIG was Mercker 86 - México, which averaged 28.87 t.ha−1 and 8.62% for the respective variables. For traits like NDF, ADF and %CEL, all genotypes except Pusa Napier no. 1 displayed good results. For C: N, as in the first harvest, genotype Cubano de Pinda showed a higher mean than the others, averaging 28.25.
Based on Tukey’s mean comparison test at the 5% probability level, the materials that most stood out at the five different production ages were genotypes King Grass, Mole de Volta Grande, and Mercker 86 - México. These results are much higher as compared with those found by Rossi (2010) for these same materials, considering that they were obtained at the production age of 10 months; higher than those reported by Morais et al. (2009).
Combined analysis of variance
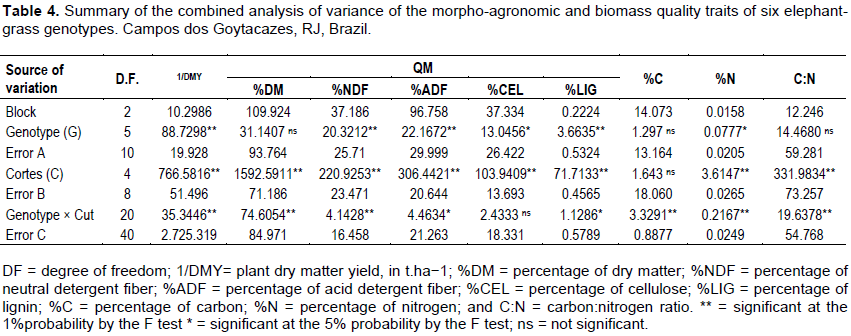
Table 4 depicts the mean squares for the effects of plot (genotypes), error A, associated with the plot effect; subplot (harvest), error B, associated with the subplot effect; interaction between plot and subplot; and error C, associated with the interaction effect, for the nine traits assessed in this study.
Most of the traits showed significant differences for the source of variation Genotype, except %DM, %C, and C: N ratio, indicating a difference between the genotypes. This result proves the distinction between the harvests performed in different periods, revealing that plants may undergo changes in their structure and morphology stemming from adverse environmental conditions. The source of variation harvests was also significant for most traits, except %C, indicating the presence of variability, which is paramount to prove the distinction between harvests performed during this study.
The only trait that did not show significance for the genotype × harvest interaction was %CEL. The significant interaction between genotypes and harvests indicates that there was a change in the classification of genotypes in different harvests. Sousa et al. (2016) evaluated elephant grass genotypes and also observed that, some were influenced by the evaluation harvest which is related to environmental conditions (temperature, luminosity, rainfall distribution), prevailing during the crop growth period.
The significant interaction between genotypes and harvests indicates that, the genotypes’ response is not the same over successive harvest that is, there are differences between the genotype means, or in the classification of their responses, along the five harvests.