Full Length Research Paper

ABSTRACT
The Indonesian fat-tailed sheep (IFTS) is a local sheep that has been long time raised and well adapted to the extreme environments of Lombok Island. The present study was conducted to determine the polymorphism of bone morphogenetic protein receptor 1B (BMPR-1B) gene and its association with litter size in the IFTS breed by employing forced polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) methods in the IFTS. Polymorphism of BMPR-1B genes on IFTS population were identified by the forced PCR-RFLP methods. The result of this study showed that mutation of BMPR-1B gene resulted in two allele of wild type (+) and mutan allele (B) with frequencies of 0.807 and 0.193, respectively and three genotype of BB (110 bp/110 bp), B+ (110 bp/140 bp), and ++ (140 bp/140 bp) with almost unequal frequencies distribution of 0.060, 0.268, and 0.672. Genetic diversity of BMPR-1B genes caused different litter sizes on individual IFTS. A highly significant difference (P<0.01) was observed in the average of litter size in different genotypes. The highest average of litter size was found in the genotype of BB, that is, 1.685 lambs.
Key words: Bone morphogenetic protein receptor 1B (BMPR-1B), gene, mutation, polymorphism, litter size.
Abbreviation: IFTS, Indonesian fat-tailed sheep; PCR-RFLP, polymerase chain reaction-restriction fragment length polymorphism; FecB, Fecundity Booroola; BMPR-1B, bone morphogenetic protein receptor type-1B; BMP-15, bone morphogenetic protein 15; GDF9, growth differentiation factor; TGF-β, transforming growth factor β; FSH, follicle stimulating hormorne.INTRODUCTION
Prolificacy in sheep is regulated by three major genes: bone morphogenetic protein receptor 1B (BMPR-1B), bone morphogenetic protein 15 (BMP-15), and growth differentiation factor 9 (GDF9). BMPs are members of the transforming growth factor β (TGF-β) superfamily. BMP is multifunctional protein which controls growth, differen-tiation and apoptosis in many types of cells and plays indispensable roles during embryogenesis and the fertility in mammals (Davis, 2005).
BMPR-1B (fecundity booroola/FecB) gene is an autosomal gene located on sheep chromosome 6 which is syntenic with human chromosome 4 and is the first major gene for prolificacy identified in sheep (Wilson et al., 2001). The BMP receptor is expressed by oocytes and granulosa cells from the primary stage to the late antral stage of follicle development and binds to BMP15. Mutation in the BMPR-1B gene is a single nucleotide non-conservative substitution that has an additive effect on ovulation rate (Davis, 2006; Pramod et al., 2013). A single copy of the gene in heterozygous ewes produced about 1.5 extra eggs and gave birth to about 1.0 extra lamb per ewe lambing. Homozygous ewes carrying two copy of the gene produced about 3.0 extra eggs resulting in about 1.5 extra lambs per ewe lambing (Davis, 2004).
The FecB mutation in sheep is due to a single nucleotide substitution (Arginine to Glutamine) at 746th position of open reading frame (ORF) in exon 6 that induces a nonsynonymous substitution of glutamine with an arginine corresponding to 249th position (Q249R) of the mature peptide (Mulsant et al., 2001). This mutation causes loss of reaction capabilities to BMP-4 that plays a central role in determining the formation of the primordial germ cell (PGC) in the ovary (Mulsant et al., 2001; Wilson et al., 2001). The demage on the BMP system during follicle development led to increase average ovulation (Fabre et al., 2006) and was reported in Australian Merino sheep, Indian Garole, Kendrapara, and Bonpala sheep (Kumar et al., 2008; Roy et al., 2011).
Genetic mechanism caused by mutations in BMPR-1B and BMP-15 genes, which has a relationship with the average number of ovulation in sheep, is still not widely known. The tendency for sheep producing twins (twining) or three (triplets) lambs are the same, although there are differences in the level of gene regulation. The Indonesian fat-tailed sheep (IFTS) is a local prolific and high economic value sheep in Indonesia. It is essential to do research on the genetics and reproduction in IFTS to identify genes with major effect on prolificacy which will be useful for increasing and accelerating the rate of genetic improvement on litter size. The aim of this study was to identify polymorphism in BMPR-1B gene and its possible association with litter size in IFTS.
MATERIALS AND METHODS
The University of Mataram, Faculty of Medicine, Ethical Committee for Medical Research, Mataram, Indonesia approved all animal procedures for this experiment (Register No.35/UN18.8/ETIK/2015).
Phenotyping (measurement of the litter size)
Two hundred and fifty IFTS (aged 1.5 to 3 years) reared under extensive communal conditions were used in this study. All animals were ear tagged. The number of lambs born for every birth of each ewe was recorded. The average litter size for each ewe of the first three parities was recorded and used for further analysis.
Sample collection and DNA isolation
Blood samples for DNA analysis were collected from the jugular vein of each animal. Blood was collected on K2EDTA 0.5 M and stored at -25°C for few weeks or at -75°C up to several months. Genomic DNA was extracted from whole blood by the phenol-chloroform method, then dissolved in TE buffer (10 mM Tris-HCl and 1 mM EDTA, pH 8.0), and kept at -20°C (Sambrook et al., 1989).
Detection of the FecB mutations
A primer pair was designed to detect single nucleotide poly-morphisms in exon 6 of the BMPR-1B gene in prolific IFTS by PCR-RFLP as described by Wilson et al. (2001). Primers amplified a 140-bp band. After digestion with Ava II (G|GWCC), the BB animals had a 110-bp band, the B+ animals had 140- and 110-bp bands, and the ++ animals had a 140-bp band. The primer sequences were as follows: Forward, 5′-GTCGCTATGGGGAAGTTTGGATG-3′ and Reverse, 5′-CAAGATGTTTTCATGCCTCATCAACACGGTC-3′.
Amplification was carried out in a volume of 25 µl. The PCR reaction contained 100 ng DNA, 0.5 µM of each primer (10 pmol/µl), 1x PCR buffer (10 mM Tris-HCl pH 9.0), 1.5 mM MgCl2 and 50 mM KCl, 5% deionized formamide, 200 µM dNTPs, and 0.025 U Taq DNA polymerase (Pharmacia). Amplification was performed for 35 cycles using DNA thermal Cycler (Perkin Elmer Cetus Corp.). The first cycle was at 95°C for 5 min followed by 35 subsequent cycles of 94°C × 45 s, 60°C × 45 s, then 72°C × 60 s, and the last cycle at 72°C for 5 min. The PCR products of 5 µl were digested separately with 10 U of Ava II (Fermentas) at 37°C over night in a 15 µl reaction mixture. The DNA fragments of BMPR-1B were separated by electrophoresis on 2.5% agarose gels. The gels were visualized with ethidium bromide staining and analyzed using an AlphaImager EP Documentation and Analysis Systems (Alpha Innotech Corporation, USA).
Statistical analysis
Genotype and allele frequency within and among genetic groups were determined by the method of Goodman adapted by Maskur et al. (2014). Association analysis was performed using general linear model (GLM) and the least square means of the genotypes were compared by t-test (as implemented in the SAS programme). The linear model used was as follows:
Y=A+LS+Gi+eij
where Y is phenotypic value of litter size, A is population mean, LS is lambing season effect, Gi is fixed effect of the BMPR-1B genotype, and eij is random error.
RESULTS AND DISCUSSION
Detection of polymorphism in BMPR-1B (FecB) gene in IFTS
The PCR product is about 140 bp, located in exon 6 of the BMPR-1B gene. Mutations at the gene can be identified using restriction enzyme Ava II (G|GACC). The digestion using Ava II (G|GACC) produces two alleles, namely the wild-type allele (+) is 140 bp (uncut) and the mutant allele (B) is 110 bp/30 bp (cut). The wild-type allele (+) was not sensitive to Ava II, while the mutant allele (B) was cut by Ava II producing two DNA fragments 110 and 30 bp. The analysis of the FecB locus implies that mutation occured to make it sensitive to Ava II enzyme which recognized the sequence (G|GACC) as the site of cutting.
Mutations at the BMPR-1B gene locus are transition mutations that change adenine bases into guanine (A/G transition) at base 746 of the coding region of the BMPR-1B gene. This point mutation results in a change in the amino acid sequence of BMPR-1B in which a glutamine (CAG) in the wild-type to an arginine (CGG) in the mutant (CAGàCGG, Q249R). The alteration of amino acid sequence causes functional change in the intracellular kinase domain of mature protein and was reported to affect ovulation rate in Australian Merino sheep, Indian Garole, Kendrapara and Bonpala sheep (Kumar et al., 2008; Roy et al., 2011), British Milk sheep, Chinese Small Tail Han and Hu sheep (Chu et al., 2007).
The FecB mutation of the BMPR-1B gene has been studied in various sheep breeds. The results indicated that IFTS carried the same FecB mutation as found in Small Tailed Han ewes (Chu et al., 2007), Greek Chios and Florina sheep (Michailidis et al., 2008) and Booroola Merino ewes (Mulsant et al., 2001; Souza et al., 2001; Wilson et al., 2001). The single nucleotide polymorphisms (SNPs) identified in exon 8 (GenBank accession number GQ863578) of Mehraban sheep was found to be associated with reproduction traits (Abdoli et al., 2013).
Allelic and genotypic frequency of BMPR1B gene in IFTS
Allelic and genotypic identification of exon 6 BMPR-1B gene using the forced PCR-RFLP technique produces two alleles, namely, the wild-type allele (+) and the mutant allele (B) with three genotypes BB, B+ and ++ (Figure 1). There are contrasting allelic frequencies distributions between B and + allele in BMPR-1B gene of IFTS. The distribution of allelic frequency of the + allele was higher than B allele, respectively of 0.807 and 0.193, while the frequency distribution of the genotypes BB, B+ and ++ were respectively 0.060, 0.268 ,and 0.672 (Table 1).
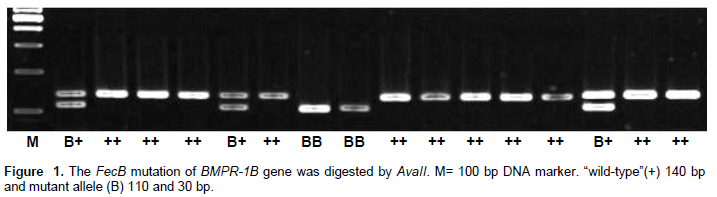

Chi-square (c2) test showed that the genotype distributions of exon 6 BMPR-1B gene was not at Hardy-Weinberg equilibrium (H-WE) in IFTS. The genotype frequencies at polymorphic loci of exon 6 BMPR-1B gene showed a highly significant difference (P<0.01). This contrasts with the same FecB mutation in the Small Tailed Han ewes in China, as reported by Chu et al. (2007) where the allelic frequencies distributions between the B and + allele differed, the + allele (0.27) being lower than that of the B allele (0.73), while the frequencies of genotypes BB, B+, and ++ were 0.52, 0.42, and 0.06, respectively.
Genetic index has an important meaning to get the description about the genetic variability (Marson et al., 2005). Data in Table 2 show the results of genetic index measurements in IFTS population. These data indicate a genotypic imbalance in the population where genotype heterozygote frequencies are higher than the Hardy- Weinberg expectation agreement. This could be due to intensive selection, resulting in a tendency towards the accumulation of certain genotypes (Tambasco et al., 2003) and the possibility of inbreeding (Machado et al., 2003). Polymorphism information content (PIC) value is commonly used in genetics as a measure of polymorphism for a marker locus used in linkage analysis. Based on the PIC value of 0.264, it can be stated that the genetic diversity of BMPR-1B gene within IFTS population is at the medium level. This statement is based on PIC levels of polymorphism as determined by Botstein et al. (1980) in which levels of ≤0.25 are classified as low, 0.25≤PIC≤0.5 are classified as medium and PIC≥0.5 are classified as high polymorphism.

Association of BMPR-1B gene polymorphism with litter size in IFTS
The association between genotype of the BMPR-1B gene and average litter size in IFTS is shown in Table 3. The effect of genotype of BMPR-1B gene was found to be significant for litter size on IFTS. BMPR-1B is a receptors for majority of the TGF-β superfamily members. One explanation could be that the action of BMPR-1B gene on the target gene through formation of the receptor complexes, causes the phosphorylation of intracellular signaling molecules called Smads, which then translocate to the nucleus and regulate transcription of target genes (Moore et al., 2002).

The effect of FecB mutation on litter size in the IFTS is lower than other breeds that had been reported by any researcher. Expression of BMPR-1B gene can varied between different breed of sheep and different environ-mental conditions or the interaction between breed of sheep and environment will give different expression (Fogerty, 2008). The difference could be due to the background genotype, environmental factors (feeding and management) such as the relatively low nutritional value of the tropical forages available to these ewes or combination of these factors. IFTS were reared under the extensive farming system in the region with dry weather along 8 to 9 months per year. This condition causes low nutritional value of forage that is available to support the production and reproduction of IFTS. Several studies showed the different expression of BMPR-1B gene in different environmental conditions. Guan et al. (2007) reported that the expression of BMPR-1B gene on Garole and Hu sheep varied between different location with different environmental conditions. The litter size of Garole sheep is 2.27 in humid rice paddies of the Sundarbans region, but lower mean litter size is 1.74 in the semiarid environment of the Deccan plateau of Maharashtra and 1.68 in Rajhastan. The avarage litter size of Hu sheep in conservation area in China is 2.12, higher than in Shanghai and Suzhou is 1.78 and 1.90 respectively. The Small Tailed Han ewes with genotypes FecBB/FecBB (BB); FecBB/FecB+ (B+) and FecB+/FecB+ (++) has litter size 2.65±0.10, 2.36±0.12, and 1.25±0.17, respectively (Chu et al., 2007). In the Awassi sheep, litter size of ++, B+, and BB ewes was 1.28, 1.90, and 1.92, respectively (Gootwine et al., 2008). Litter sizes of the Javanese sheep with genotypes FecBB/FecBB (BB), FecBB/FecB+ (B+), and FecB+/FecB+ (++) for FecB have been measured at 2.59, 1.95, and 1.24, respectively (Inounu, 1996).
The data in Table 3 indicates a significant increase in litter size of IFTS that carry FecB mutations in both heterozygous and homozygous condition. This implies that the sheep with FecB mutations has granulosa cells and are more sensitive to follicle stimulating hormorne (FSH); thus, the cells divides more actively leading to ovarian follicles becoming adults and mature (Davis 2008; Fabre et al., 2006). The prolific nature occurs based on the concept that increased ovulation rate lead to an increasing number of ovulated oocytes and if fertilization occurs and embryo viability can be maintained by the parent, it will be followed by the birth of a child more than one (Wilson et al., 2001).
CONCLUSION
Polymorphism of FecB gene was identified in IFTS by forced PCR-RFLP method. Restriction enzyme digestion with AvaII for FecB gene resulted in three genotypes of BB, B+ and ++ with a frequency of 0.060, 0.268 and 0.672, respectively and two alleles of wild type (+) and mutan (B) with almost unequal frequency distribution of 0.807 and 0.193. Genotypic polymorphism of BMPR-1B has a significant influence on litter size in IFTS. Higher litter size was observed in BB genotype as compared to B+ and ++ gentopes of FecB gene.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ABBREVIATIONS
IFTS, Indonesian fat-tailed sheep; PCR-RFLP, polymerase chain reaction-restriction fragment length polymorphism; FecB, Fecundity Booroola; BMPR-1B, bone morphogenetic protein receptor type-1B; BMP-15, bone morphogenetic protein 15; GDF9, growth differentiation factor; TGF-β, transforming growth factor β; FSH, follicle stimulating hormorne.
REFERENCES
|
Abdoli R, Zamani P, Deljou A, Rezvan H (2013). Association of BMPR1B and GDF9 genes polymorphisms and secondary protein structure changes with reproduction traits in Mehraban ewes. Gene 524(2):296-303. |
|
|
Botstein D, White RL, Skolnick M, Davis RW (1980). Construction of a genetic linkage map in human using restriction fragmenth length polymorphisms. Am. J. Hum. Genet. 32:314-331. |
|
|
Chu MX, Liu ZH, Jiao CL, He YQ, Fang L, Ye SC, Chen GH, Wang JY (2007). Mutations in BMPR-IB and BMP-15 genes are associated with litter size in Small Tailed Han sheep (Ovis aries). J. Anim. Sci. 85:598-603 |
|
|
Davis GH (2004). Fecundity genes in sheep. Anim. Reprod. Sci. 82-83:247-253. |
|
|
Davis GH (2005). Major genes affecting ovulation rate in sheep. Genet. Sel. Evol. 37(Suppl. 1):S11-S2. |
|
|
Davis GH (2008). The Booroola gene: origin, distribution, use and management of the FecB mutation. Use of the FecB (Booroola) gene in sheep-breeding programs. Proc. of the Helen Newton Turner Memorial International Workshop held in Pune. Maharashtra. India. 1:22-29. |
|
|
Davis GH, Balakrishnan L, Ross IK, Wilson T, Galloway SM, Lumsden BM, Hanrahan JP, Mullen M, Mao XZ, Wang GL, Zhao ZS, Zeng YQ, Robinson JJ, Mavrogenis AP, Papachristoforou C, Peter C, Baumung R, Cardyn P, Boujenane I, Cockett NE, Ey-thorsdottir E, Arranz JJ, Notter D (2006). Investigation of the Booroola (FecB) and Inverdale (FecXI) mutations in 21 prolific breeds and strains of sheep sampled in 13 countries. Anim. Reprod. Sci. 92:87-96. |
|
|
Fabre S, Pierre A, Mulsant P, Bodin L, Di Pasquale E, Persani L, Monget P, Monniaux D (2006). Regulation of ovulation rate in mammals: contribution of sheep genetic models. Reprod. Biol. Endocrinol. 4:20. |
|
|
Fogerty (2008). Environmental modulation of FecB expression. Use of the FecB (Booroola) gene in sheep-breeding programs. Proc. of the Helen Newton Turner Memorial International Workshop held in Pune. Maharashtra. India. 1:66-75. |
|
|
Gootwine E, Reicher S, Rozov A (2008). Prolificacy and lamb survival at birth in Awassi and Assaf sheep carrying the FecB (Booroola) mutation. Anim. Reprod. Sci. 108(3-4):402-411. |
|
|
Guan F, Liu SR, Shi GQ, Yang LG (2007). Polymorphism of FecB gene in nine sheep breeds or strains and its effects on litter size, lamb growth and development. Anim. Reprod. Sci. 99:44-52. |
|
|
Inounu I (1996). Keragaan produksi ternak domba prolifik [disertasi]. Bogor: Program Pascasarjana. Institut Pertanian Bogor. |
|
|
Kumar S, Mishra AK, Kolte AP, Dash SK, Karim S (2008). Screening for Booroola (FecB) and Galway (FecXG) mutations in Indian sheep. Small Rumin. Res. 80(1-3):57-61. |
|
|
Machado MA, Schuster I, Martinez ML, Campos AL (2003). Genetic diversity of four cattle breeds using microsatellite markers. Rev. Bras. Zootec. 32:92-98. |
|
|
Marson EP, Ferraz JB, Meirelles FV, Balieiro JC, Eler JP, Figueiredo LG, Mourao GB (2005). Genetic characterization of European-Zebu composite bovine using RFLP markers. Genet. Mol. Res. 4(3):496-505. |
|
|
Maskur, Rodiah, Chairussyuhur Arman (2014). Association of a novel single nucleotide polymorphism in growth hormone receptor gene with production traits in Bali cattle. Ital. J. Anim Sci. 13:3461 |
|
|
Michailidis G, Melpomeni A, Pappa V (2008). Reproductive performance and investigation of BMPR-IB and BMP-15 gene mutations in Greek Chios and Florina sheep breeds. Arch. Zootech. 11(1):24-31. |
|
|
Moore RK, Otsuka F, Shimasaki S (2003). Molecular basis of bone morphogenetic protein-15 signaling in granulosa cells. J. Biol. Chem. 278:304-310. |
|
|
Mulsant P, Lecerf F, Fabre S, Schibler L, Monget P, Lanneluc I, Pisselet C, Riquet J, Monniaux D, Callebaut I, Cribiu E, Thimonier J, Teyssier J, Bodin L, Cognie Y, Chitour N, Elsen JM (2001). Mutation in the bone morphogenetic protein receptor-IB is associated with increased ovulation rate in Booroola ewes. Proc. Natl. Acad. Sci. U.S.A. 98: 5104-5109. |
|
|
Pramod KR, Sharma SK, Kumar R, Rajan A (2013). Genetics of ovulation rate in farm animals. Vet. World 6(11):833-838. |
|
|
Roy J, Polley S, De S, Mukherjee A, Batabyal S, Pan S, Brahma B, Datta TK, Goswami SL (2011). Polymorphism of fecundity genes (FecB, FecX, and FecG) in the Indian Bonpala sheep. Anim. Biotechnol. 22(3):151-162. |
|
|
Sambrook J, Fritsch EF, Maniatis T (1989). Molecular Cloning : A laboratory Manual. 2nd edition. Cold Spring Harbor Laboratory Press, USA. |
|
|
Souza CJH, Mac Dougal C, Campbell BK, McNeilly AS, Baird DT (2001). The Booroola (FecB) phenotype is associated with a mutation in the bone morphogenetic receptor type 1B (BMPR-1B) gene. J. Endocrinol. 169:1-6. |
|
|
Tambasco DD, Paz CCP, Tambasco-Studart, Pereira AP, Alencar MM, Freitas AR, Coutinho LL, Packer IU, Regitano LCA (2003). Candidate genes for growth traits in beef cattle crosses Bos taurus X Bos indicus. J. Anim. Breed. Genet. 120:51-56. |
|
|
Wilson T, Wu XY, Juengel JL, Ross IK, Lumsden, JM, Lord EA, Dodds KG, Walling GA, McEvan JC, O'Connel AR, McNatty, KP, Montgomery GW (2001). Highly prolific Booroola sheep have a mutation in the intracellular kinase domain of bone morphogenetic protein IB receptor (ALK-6) that is expressed in both oocytes and granulosa cells. Biol. Reprod. 64:1225-1235. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


