ABSTRACT
The actinobacteria are the most abundant bacterial group in the soil, where they present different characteristics and antagonistic effects on other microorganisms. The objectives of this study were to characterize and evaluate the antagonistic effect of actinobacteria from the semiarid region on rhizobia from the same climatic region. Tests were performed to observe chromogenic and micro-morphological characteristics, tolerance to pH levels of melanin production, and use of carbon sources. This occurred due to the use of modified CDA culture media and basal medium supplemented with specific carbon sources, as well as the use of Ral color chat to observe the colonies. The in vitro antagonism on rhizobia was evaluated in yeast malt agar (YMA) culture medium for the observation of inhibition halo by actinobacteria. We observed different characteristics in relation to the color of aerial mycelium and reverse pigments; tolerance to media with acid and alkaline pH; and production of melanoid pigment occurred only in three strains. The morphological characteristics were described for genera Streptomyces and Saccharothrix, later confirmed by molecular sequencing. These genera were generalists in use of carbon sources, showing their physiological versatility. Regarding antagonism, nine strains of the genus Streptomyces were able to inhibit in vitro strains of Rhizobium tropici and Bradyrhizobium yuanmingense also coming from semiarid soils. The antagonism between actinobacteria and rhizobia can directly affect the symbiosis between nitrogen-fixing bacteria and leguminous plants. These results are pioneering in observing antagonism of these species of rhizobia under the climate condition of the semiarid region, and may contribute to agricultural biotechnology.
Key words: Streptomyces, soil, microbe interactions, Brazil, agricultural biotechnology, actinobacteria, antagonistic effect.
Several mechanisms occur in the soil that influence the structure and composition of the soil communities, as well as intra and inter-specific interactions, for example, competition. The competitor microbial species often secrete compounds that affect the fitness of other species. This fact is characterized by antibiosis (Morris
and Blackwood, 2007). The actinobacteria are an example of organisms that antagonize microorganisms by means of secondary metabolites (Karlovsky, 2008; Tarkka and Hampp, 2008) and/or secretion of enzymes that inhibit the growth of other organisms in the soil (Suneetha and Zaved, 2011).
These filamentous microorganisms are gram-positive bacteria, and have a high content of guanine and cytosine in their DNA (Monciardini et al., 2002; Flärdh and Buttner, 2009). They are abundant and dispersed in different ecosystems; soil is their main habitat, where they perform important functions in soil fertility (El-Tarabily and Sivasithamparam, 2006; Jayasinghe and Parkinson, 2008). The genera Streptomyces, Nocardia, Micromonospora, Actinoplanes and Streptosporangium stand out in the edaphic environment (Anandan et al., 2016). The actinobacteria have crucial role in the decomposition of organic matter in ecosystems and thereby influence the cycling of nutrients in the soil (Mabrouk and Saleh, 2014), making them an excellent indicator of biological activity of the soil (Arifuzzaman et al., 2010). On the other hand, the literature contains records of the antagonistic effect of actinobacteria, in vitro and/or in vivo on other microorganisms (Parmar and Dufresne, 2011), for example, when they interact with rhizobia (Gregor et al., 2003; Mingma et al., 2014).
Rhizobia are bacteria responsible for biological nitrogen fixation (BNF) through the established symbiosis with leguminous plants (Zilli et al., 2010). These bacteria are of great importance for maintenance and restoration of ecosystems, being useful for the recovery of areas degraded by different impacts (Pontes et al., 2012). Soils with high population density of actinobacteria can inhibit the growth of rhizobia, negatively affecting plant nodulation and consequently reducing nitrogen fixation and plant productivity (Pereira et al., 1999; Parmar and Dufresne, 2011).
There are few studies that address the relationship of actinobacteria versus rhizobia, especially in isolates from semiarid regions. Thus, the aim of this study was to characterize the actinobacteria obtained in the Brazilian semiarid region and test their antagonistic effect in vitro on rhizobia strains from semiarid soils.
Microorganisms
Fourteen strains of actinobacteria (Lima et al., 2014) obtained from samples of soil (AC 46, AC 49, AC 50 and AC 56) and leaf litter ((AC 3, AC 5, AC 10, AC 12, AC 13, AC 14, AC16, AC 38, AC 42 and AC43) at the Aiuaba Ecological Station (6°40' S & 40°10' W),located in the southwestern part of the municipality of Aiuaba, Ceará state, and five strains of rhizobia (L 4, L 7, L 9, L 16, L 18 and L 22) originating from the rhizosphere of leguminous plants (Mimosa hostilis, Mimosa caesalpiniaefolia, Poincianella bracteosa and Erythrina verna) (Pinheiro et al., 2014) in areas of Ceará (4°58' S and 39°1' W and 4°7' S and 38°14' W), and the state of Rio Grande do Norte (5°39' S to 35°58' W and 5°57' S to 36°39' W) were used in this study. All microorganism collection sites are located in the semiarid region of northeastern Brazil with prevailing vegetation of Caatinga, shallow soils rich in minerals, and an average annual rainfall of 400 to 800 mm. These microorganisms belong to the collection of the Environmental Microbiology Laboratory of Micro-organisms (LAMAB) of Federal University of Ceará, Department of Biology.
Morphological characterization
The actinobacteria strains were cultured in Casein Dextrose Agar (CDA) medium (0.5 g of K2HPO4, 0.2 g of MgSO4.7H2O, 0.01 g of FeSO4.7H2O, 2.0 g of dextrose, 0.2 g of casein, 15 g of agar, 1000 mL of distilled water, and pH 6.5 to 6.6) (Clark, 1965), for seven days at 28°C in a biochemical oxygen demand (BOD) chamber. The chromogenic characterization was performed by observing the aerial and reverse mycelium after growth of strains in Petri dishes, according to Wink (2012), with the use of a RAL color chart. The micro-morphological characterization was carried out by microcultivation according to Kern and Blevins (2003), with modifications. A blade and two pieces of cotton moistened with distilled water (sterile) were placed in sterilized Petri dishes. A cube of CDA culture medium measuring approximately 1 cm³ (Clark, 1965) was placed on the blade. The strain was inoculated on the cube sides, covered with a sterile coverslip, while the dish was closed and incubated in a BOD chamber for 7 to 14 days at 28°C. After this time, the coverslip was removed and placed on a clean slide containing a drop of cotton blue stain. The slides were observed under a Zeiss Axioplan optical microscope (Leica DM750 M, Heerbrugg, Switzerland) with a 100x magnification to visualize the actinobacteria’s characteristics (Miyadoh, 1997; Goodfellow et al., 2012).
Utilization of carbon sources
The use of carbon sources was analyzed according to Shirling and Gottlieb (1966), using mineral salts agar as basal medium. A solution was prepared by 10% of each carbon source (arabinose, sucrose, xylose, inositol, mannitol, fructose, rhamnose, raffinose and cellulose), with glucose (positive control) and water (negative control). After this, each dish with medium was sterilized and cooled to about 60°C, a concentration of 1% of each carbon source was added. These carbon sources were sterilized by a Millipore filter (0.45 µm). Finally, the strains were transferred to Petri dishes (triplicates) and incubated in a BOD chamber at 28°C for 10 days. The use of the sources was analyzed by the observation of increase (+) or no growth (-) in Petri dishes with medium.
Production of melanoid pigment
Strains were grown in tyrosine agar medium (Shinobu, 1958) with and without tyrosine. The Petri dishes were incubated at 28°C in a biochemical oxygen demand (BOD) chamber for 10 days. The formation of melanoid pigment was considered (formation of dark-colored pigment) after growth in a Petri dishes with tyrosine, this is because the pigment formation in both media does not reflect the production of melanin.
Tolerance to different pH values
The test for growth of actinobacteria at different pH was carried out according to Kishore et al. (2012), modified by using the CDA culture medium (Clark, 1965). The medium was adjusted to pH 4.0, 5.0, 6.0, 7.0, 8.0 or 9.0. The strains were transferred to Petri dishes (triplicates) with a solid medium and incubated at 28°C for 10 days. The growth of the strains will indicate tolerance of the tested pH. The soil from which the actinobacteria had been collected was analyzed to determine the pH according to Donagema et al. (2011).
Genomic DNA extraction and analysis of genetic variability and amplification of 16S rRNA
Total genomic DNA was obtained from actinobacteria and rhizobia belonging to the microbiological collection with the Wizard® genomic DNA purification kit (Promega, Madison, WI, USA), following the manufacturer's instructions. The Wizard® genomic DNA kit is based on a four-step process.
The first step is the purification procedure, lysing of the cell and the nucleus; an RNase digestion step was included at this time. The cellular proteins are then removed by a salt-precipitation step, which precipitates the proteins but leave the high molecular weight genomic DNA in solution. Finally, the genomic DNA was concentrated and desalted by isopropanol precipitation. The oligonucleotides (Tabe 1), enterobacterial repetitive intergenic consensus (ERIC) were used to evaluate the genetic diversity of strains (Tian-Xing, 2011).
The polymerase reaction (PCR) was performed with a final volume of 25 μL, using 60 ng of DNA; 2.5 μL of a reaction buffer 10x; 0.75 μL of MgCl2 (50 mM); 2 μL of dNTP (2.5 mM); 1 μL of forward oligonucleotide, and 1 μL of reverse oligonucleotide (both in concentration of 10 pmol); 0.25 μL of Taq polymerase enzyme (5 U/μL) and ultrapure q.s.p. water to 25 μL. Amplification was performed using the following steps: initial denaturation at 94°C for 2 min followed by 30 cycles at 94°C for 1 min, 50°C for 1.5 min and 68°C for 4 min, and final extension at 68°C for 10 min. The ERIC-PCR product was subjected to electrophoresis on 1.2% agarose gel in triton extraction buffer (TEB) buffer 0.5x stained with ethidium bromide and was visualized under ultraviolet light. For amplification of 16S rRNA, the primers 3 and 4 (Table 1) were for the rhizobia (Weisburg et al., 1991), and the oligos 5 and 6 (Table 1) for the actinobacteria (Rintala et al., 2001).

The final volume of the PC reaction was 25 µL and amplification conditions are presented in Table 2. The reaction was carried out in an AmpliTherm thermal cycler. The amplicons generated by the PC reactions were purified using ExoSAP (exonuclease I, shrimp alkaline phosphatase). Sequencing was performed by the Sanger method with the BigDye Kit ABI3100 and an Applied Biosystem sequencer. The genetic diversity of the isolates was assessed by ERIC-PCR markers. The sequences were compared with existing sequences in the GenBank database using the BLAST program available at the site National Center for Biotechnology Information (NCBI; www.ncbi.nlm.nih.gov).
Antagonistic test in vitro between actinobacteria and rhizobia
The antagonistic effect of actinobacteria on the rhizobia was evaluated according to the methods of Gregor et al. (2003). The strains of rhizobia were grown in glass tubes with liquid (without agar) YMA medium (10 g of mannitol, 0.5 g of K2HPO4, 0.2 g of MgSO4.7H2O, 0.1 g of NaCl, 0.5 g of yeast extract, 5 mL of bromothymol blue, 15 g of agar, 1000 mL of distilled water, and pH 6.8) (Vincent, 1970), and shaken (150 rpm) on a shaker orbital for about seven days. The actinobacteria were also grown in glass tubes, but with liquid CDA medium on a shaker orbital at the same speed and time period as foe rhizobia. The inhibition test was performed in Petri dishes (90 mm in diameter), divided into four quadrants with solid (with agar) YMA medium, plus 100 uL of YMA broth spread with rhizobia with a Drigalski handle. Then, 5 uL of CDA broth with actinobacteria was added. The Petri dishes (triplicate) were incubated for 5 days at 28°C in a BOD chamber. After this period, the plates were evaluated by observing the presence of inhibition zones, which were measured (cm) using a caliper rule.
Data analysis
The number of strains with and without growth at the different pH levels were subjected to the chi-square test with p ≤ 0.05, based at pH 7. The radii of the inhibition halos were measured and then the inhibition area (πr²) was calculated. These area values were log-transformed. The Shapiro-Wilk test was applied to test for normality, and then the data were subjected to analysis of variance (ANOVA) at p ≤ 0.05 with the agricolae package software R®.
Chromogenic and morphological characteristics
The strains from the semiarid region had different characteristics (Table 3), in relation to color from aerial mycelium and reverse pigment. The predominant colors in the air mass were brown (5), green (4), beige (2), and only 1 each of grey, white and cream. The pigmentations of the reverse mycelium were brown (5), green (3), yellow (3), cream (2) and beige (1). After the microcultivation, it was observed that the actinobacteria had the following morphological characteristics: spore chain hooked to looped, spore chain straight to flexuous, long chains of smooth spores, spiral spore chain and smooths pore surface.These morphological traits are similar and described for genera Streptomyces and Saccharothrix.
pH tolerance and melanin production
Table 4 shows the growth characteristics of the strains at different pH and the production of melanin. Regarding tolerance to extreme pH levels, the strains of the two genera had different behavior, except at pH 7.0 and 8.0, where all were able to grow in CDA medium with modified pH. The pH of the soil samples from which the actinobacteria were obtained had acid characteristics ranging from 4.5 to 6.3. Finally, melanoid pigment production occurred only in three strains (AC 3, AC 10 and AC 49) of actinobacteria.
Use of carbon sources
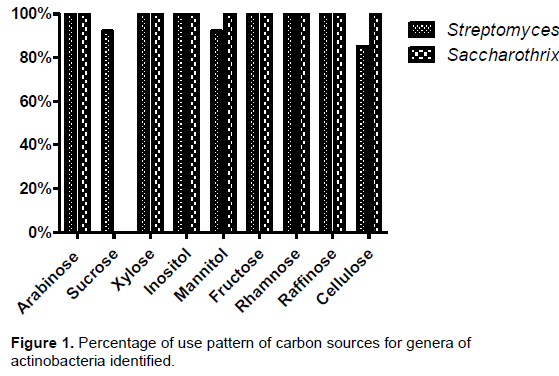
The strains of actinobacteria had wide use of carbon sources (Figure 1), with glucose being used by all strains tested. The strains of the genus Saccharothrix, used all the carbon sources except sucrose. However, the use of these sources by strains of the genus Streptomyces varied, with 85% for cellulose, 92% for sucrose and mannitol, and 100% for the other sources.
Molecular identification of microorganisms
The amplification profile ranged from 1 to 9 amplicons
with sizes of 500 to 1400 bp for actinobacteria and 4 to 12 amplicons with sizes from 300 to 1400 bp for strains of rhizobia. After the molecular sequencing of actinobacteria and rhizobia, we observed the presence of actinobacteria of the genera: Saccharothrix and Streptomyces (Table 5). Regarding rhizobia, we identified two genera: Bradyrhizobium and Rhizobium.

In vitro inhibition of rhizobia
Only the actinobacteria strains of the genus Streptomyces had an antagonistic effect on in vitro rhizobial strains tested (Table 6). Of the 14 tested actinobacteria, nine presented antibiosis, and of five strains of rhizobia used, only two were inhibited. There was no statistical difference between the inhibition of actinobacteria (F = 1.55 and P = 0.14) and between the rhizobia strains inhibited (F = 1.29 and P = 0.28). However, Rhizobium tropici was inhibited by six strains of Streptomyces, particularly Streptomyces graminisoli, with 9.4 cm² inhibition halo. Bradyrhizobium yuanmingense was antagonized by seven strains of Streptomyces, particularly Streptomyces sp. with 6.8 cm² inhibition area.
Observation of the colors of the aerial mycelium mass and reverse side is one of the first methods used to distinguish isolates.
In this study, brown was the predominant color, in contrast to those observed by Ramos et al. (2015) and Silva et al. (2015), who in characterizing actinobacteria strains coming from the semiarid region by cultures noted the predominance of gray, cream and white. This color variation is widely reported in the literature in actinobacteria strains originating from different regions, such as the Himalayas (Duraipandiyan et al., 2010), India (Kumar et al., 2010; Kumar et al., 2012; Das et al., 2014; Amsaveni et al., 2015), China (Yu et al., 2015), Egypt (Mabrouk e Saleh, 2014) and Iraq (Jaralla et al., 2014).
The color differences in the strains can be related to factors such as temperature, soil type, pH and carbon sources in the environment (Amal et al., 2011). However, since the pigmentation of the colonies is similar among the different genera of actinobacteria, this feature would not be decisive for the strains classification.
Through microcultivation it was possible to distinguish two genera. This result of the spore chain through microcultivation corroborates the molecular identification analysis. This number was lower than that reported by Brito et al. (2015), who observed a similar genus (Streptomyces) to those found for this study. The genus Streptomyces was previously reported by Silva et al. (2013) in soils from the Brazilian Cerrado which is the most common in several regions (Anandan et al., 2016), with reports in the literature describing its occurrence in Atacama desert soils in Chile (Okoro et al., 2009), India (Kumar et al., 2012; Das et al., 2014) and Egypt (Mabrouk and Saleh, 2014).
Most actinobacteria strains did not produce melanoid pigments, but two (one of each genus), were able to produce melanin. Ramos et al. (2015) also observed this lower number of melanin-producing strains in strains coming from the semiarid region. Strains of Streptomyces derived from soil in India (Dastager et al., 2006) and Egypt (Mabrouk and Saleh, 2014) were characterized as not producing this pigment.
In mangrove sediments, Janaki et al. (2014) found a similar percentage of isolates capable of producing melanin as observed for us. This pigment is synthesized by fermentative oxidation, and has properties that can protect microorganisms against gamma and ultraviolet radiation (Amal et al., 2011; Manivasagan et al., 2013; Ahmed et al., 2014); this mechanism is against environmental stresses (Zhu et al., 2007; Manivasagan et al., 2013). However in our work, this mechanism was only used by some isolated actinobacteria from litter leaf (AC 3 and AC10), perhaps due to direct exposure to sunlight.
Regarding tolerance to pH, there was greater growth range of Streptomyces strains in media with pH 4.0, 7.0 and 8.0, but some strains were able to grow with other levels. However, the Saccharothrix strain only grew at pH 7.0 and 8.0. In soils of Colorado (USA), Lauber et al. (2009) found similar behavior to that of actinobacteria strains from the semiarid region of Brazil, an increase in pH from < 4.0 to > 8.0, while in coastal sediments in India, Ramesh and Mathivanan (2009) observed strains at pH 7.0 to 8.5, with most isolates between pH 8.1 to 8.5. But the variation in pH observed in the literature was different from that found in the soil from which actinobacteria were isolated. Actinobacteria are distributed in soils of different pH levels and are generally sensitive to extreme pH.
However, some genera can grow in acidophilus habitats (Anandan et al., 2016), with the most abundant microbial communities being found at pH < 4.1 (Rousk et al., 2010), and in alkaline soils (Meena et al., 2013). This allows actinobacteria to adapt and colonize different types of environments (Shivlata and Satyanarayana, 2015).
The carbon sources were used by both genera, except for sucrose by Saccharothrix. This general use has been reported in actinobacteria from marine sediments (Augustine et al., 2013; Meena et al., 2013) and soil of various habitats (Das et al., 2014; Jaralla et al., 2014.). Species within the genus are diversified regarding the use of sources, based on what the environment offers (Amal et al., 2011; Goodfellow et al., 2012.). This may explain the variation in some strains of the genus Streptomyces. Thus, the usage profile of sugars and alcohol (inositol) suggests potential generalist ability of the strains, and these differences in use may be due to the availability of carbon sources and adjustment of the isolates to various niches (Augustine et al., 2013).
The actinobacteria of the genus Streptomyces inhibited in vitro rhizobia strains, forming inhibition zones ranging from 2.2 cm² to 9.4 cm², with antagonistic interaction with the species Rhizobium tropici and Bradyrhizobium yuanmingense. These two species of rhizobia, is known to nodulate some legume genera (Phaseolus, Leucaena and Lespedeza), vary in some traits, such as growth time (faster in R. tropici and slower in B. yuanmingense) and reaction in YMA culture medium (acid for R. tropici and alkaline for B. yuanmingense) (Martinez-Romero et al., 1991; Yau et al., 2002). The first record of antagonistic activity of actinobacteria against rhizobia was reported by Landerkin and Lochhead (1948), which test actinobacteria strains isolated from soil of the genus Rhizobium, and Van Schreven (1964) that compared actinobacteria of the genera Streptomyces, Actinomyces and Nocardia over Rhizobium strains. In Bradyrhizobium japonicum, Gregor et al. (2003) observed inhibition of these bacteria in in vitro growth by actinobacteria of the genera Streptomyces and Amycolatopsis. Mingma et al. (2014) isolated actinobacteria from roots and rhizosphere of leguminous plants which showed that, strains of the genus Streptomyces have negative effects on the growth of strains of Rhizobium sp. and Rhizobium japonicum.
There are reports in literature of such inhibition of rhizobia B. japonicum and B. elkanii, used as inoculants for soybean by actinobacteria of Brazilian Cerrado soils (Pereira et al., 1999). This work is the first reported observation of an antagonistic relationship, between actinobacteria and rhizobia coming from semiarid region soils from Brazil. The antagonistic inhibition occurs in the rhizosphere host with the release of secondary metabolites, enzymes and antibiotics (Karlovsky, 2008; Tarkka and Hampp, 2008; Suneetha and Zaved, 2011). The acquisition of nitrogen in some plants such as legumes often occurs by symbiosis with rhizobia, and the effect of inhibiting actinobacteria on rhizobia in the soil can be the cause of failure of nodulation process, and hence stunted plant growth (Tarkka and Hampp, 2008; Parmar and Dufresne, 2011).
Finally, results obtained expand knowledge of actinobacteria of the semiarid region of Brazil, and show the occurrence of an antagonistic effect exerted by these microorganisms on the rhizobia R. tropici and B. yuanmingense. This information can serve as a base for future studies, aiming to confirm this relationship and the in vivo effect which may contribute to agricultural biotechnology.
The authors have not declared any conflict of interests.
REFERENCES
|
Ahmed ES, Keyba, HM, Moussa, LA (2014). Gamma ray induced effects on new species of Streptomyces (AEFO2) (HM775973.1GI:302495616) isolated from Egyptian soil. Int. J. Bioassays. 3: 3586-3593.
|
|
|
|
Amal AM, Abeer KA, Samia HM, Nadia AEH, Ahmed KA, El-Hennawi HM (2011). Selection of pigment (melanin) production in Streptomyces and their application in printing and dyeing of wool fabrics. Res. J. Chem. Sci. 1(5): 22-28.
|
|
|
|
Amsaveni R, Sureshkumar M, Vivekanandhan G, Bhuvaneshwari V, Kalaiselvi M, Padmalochana K, Preethikaharshini J (2015). Screening and isolation of pigment producing Actinomycetes from soil samples. J. Biosci. Nanosci. 2(2):24-28.
|
|
|
|
Anandan R, Dharumadurai D, Manogaran GP (2016). An Introduction to Actinobacteria. In: Dhanasekaran D, Jiang Y (eds) Actinobacteria: Basics and Biotechnological Applications. Intech, Rijeka. Pp. 3-37.
|
|
|
|
Arifuzzaman M, Khatun MR, Rahman H (2010). Isolation and screening of actinomycetes from Sundarbans soil for antibacterial activity. Afr. J. Biotechnol. 9(29): 4615-4619.
|
|
|
|
Augustine D, Jacob J C, Ramya KD, Philip R (2013). Actinobacteria from sediment samples of Arabian Sea and Bay of Bengal: Biochemical and physiological characterization. Inte. J. Re. Mar. Sci. 2(2): 53-63.
|
|
|
|
Brito FAE, Ramos KA, Silva RM, Martins CM, Martins SCS (2015). Actinobacteria from rizospheric soil in the caatinga biome. Encicl. Biosf. 11:1992-2004.
|
|
|
|
Clark FE (1965). Actinomyces. In: Black CA (ed) Methods of soil analysis. American Society of Agronomy, Madison. pp. 1498-501.
|
|
|
|
Das P, Solanki R, Khanna M (2014). Isolation and screening of cellulolytic actinomycetes from diverse habitats. Int. J. Adv. Biotec. Res. 5(3):438-451.
|
|
|
|
Dastager SG, Li WJ, Dayanand A, Tang SK, Tian XP, Zhi XY, Xu LH, Jiang CL (2006). Seperation, identification and analysis of pigment (melanin) production in Streptomyces. Afr. J. Biotechnol. 5(8): 1131-1134.
|
|
|
|
Donagema GK, Campos DVB, Calderano SB, Teixeira WG, Viana JHM (2011). Manual de métodos de análise de solos. Embrapa Solos. Rio de Janeiro. 212p.
|
|
|
|
Duraipandiyan V, Sasi AH, Islam VIH, Valanarasu M, Ignacimuthu S (2010). Antimicrobial properties of actinomycetes from the soil of Himalaya. J. Mycol. Med. 20:15-20.
Crossref
|
|
|
|
El-Tarabily KA, Sivasithamparam K (2006). Non-streptomycete actinomycetes as biocontrol agents of soil-borne fungal plant pathogens and as plant growth promoters. Soil. Biol. Biochem. 34: 1-16.
Crossref
|
|
|
|
Flärdh K, Buttner MJ (2009). Streptomyce morphogenetics: Dissecting differentiation in a filamentous bacterium. Nat. Rev. Microbiol. 7:36-49.
Crossref
|
|
|
|
Goodfellow M, Kämpfer P, Busse H, Trujillo ME, Suzuki K, Ludwig W, Whitman WB (2012). Bergey's Manual of Systematic Bacteriology, Volume 5: The Actinobacteria. Springer. Heidelberg. 2083p.
|
|
|
|
Gregor AK, Klubek B, Varsa EC (2003). Identification and use of actinomycetes for enhanced nodulation of soybean co-inoculated with Bradyrhizobium japonicum. Can. J. Microbiol. 49(8): 483-491.
Crossref
|
|
|
|
Janaki T, Nayak BK, Ganesan T (2014). Different pre-treatment methods in selective isolation of Actinomycetes from mangrove sediments of Ariyankuppam back water estuary, Puducherry. Int. J. Adv. Res. Biol. Sci. 1(6):154-163.
|
|
|
|
Jaralla EM, Al-Dabbagh NN, Hameed N, Abdul-Hussain N (2014). Screening for Enzymatic Production Ability and Antimicrobial Activity of Actinomycetes Isolated from Soil in Hillah/Iraq. Int. J. Pharm. Biol. Sci. 9:42-47.
|
|
|
|
Jayasinghe BATD, Parkinson D (2008). Actinomycetes as antagonists of litter decomposer fungi. Appl. Soil. Ecol. 38:109-118.
Crossref
|
|
|
|
Karlovsky P (2008). Secondary Metabolites in Soil Ecology. In: Karlovsky P (ed) Secondary Metabolites in Soil Ecology. Springer. Heidelberg. pp. 1-19.
Crossref
|
|
|
|
Kern ME, Blevins KS (2003). Micologia médica – Texto e Atlas. Editoral Premier. São Paulo. 256p.
|
|
|
|
Kishore P (2012). Isolation, characterization and identification of Actinobacteria of Mangrove ecosystem, Bhitarkanika, Odisha. Dissertation, National Institute of Technology Rourkela.
|
|
|
|
Kumar N, Singh RK, Mishra SK, Singh AK, Pachouri UC (2010). Isolation and screening of soil Actinomycetes as source of antibiotics active against bacteria. Int. J. Microbiol. Res. 2(2):12-16.
Crossref
|
|
|
|
Kumar PS, Raj JPP, Duraipandiyan V, Ignacimuthu S (2012). Antibacterial activity of some actinomycetes from Tamil Nadu, India. Asian. Pac. J. Trop. Biomed. 2(12):936-943.
Crossref
|
|
|
|
Landerkin GB, Lochhead AG (1948). A comparative study of the activity of fifty antibiotic actinomycetes against a variety of soil bacteria. Can. J. Microbiol. 26: 501-506.
Crossref
|
|
|
|
Lauber CL, Hamady M, Knight R, Fierer N (2009). Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. Appl. Environ. Microbiol. 75(15):5111-5120.
Crossref
|
|
|
|
Lima JVL, Pinheiro MS, Fiúza LMCG, Martins SCS, Martins CM (2014). Microbial populations cultivable litter soil and of a storage unit in brazilian semi-arid. Encicl. Biosf. 10(18):2300-2316.
|
|
|
|
Mabrouk MI, Salen NM (2014). Molecular identification and characterization of antimicrobial active actinomycetes strains from some Egyptian soils. Am. Eurasian. J. Agric. Environ. Sci. 14:954-963.
|
|
|
|
Manivasagan P, Venkatesan J, Sivakumar K, Kim SK (2013). Marine actinobacterial metabolites: current status and future perspectives. Microbiol. Res. 168(6): 311-332.
Crossref
|
|
|
|
Martinez-Romero A, Segovia L, Mercante FM, Franco AA, Graham P, Pardo MA (1991). Rhizobium tropici, a novel species nodulating Phaseolus vulgaris L. Beans and Leucaena sp. trees. Int. J. Syst. Bacteriol. 41: 417-426.
Crossref
|
|
|
|
Meena B, Rajan LA, Vinithkumar NV, Kirubagaran R (2013). Novel marine actinobacteria from emerald Andaman & Nicobar Islands: a prospective source for industrial and pharmaceutical by products. BMC Mol. Biol. 13(1):1-17.
Crossref
|
|
|
|
Mingma R, Pathom AW, Trakulnaleamsai S, Thamchaipenet A, Duangmal K (2014). Isolation of rhizospheric and roots endophytic actinomycetes from Leguminosae plant and their activities to inhibit soybean pathogen, Xanthomonas campestris pv. Glycine. World. J. Biol. Psychiatr. 30(1):271-280.
Crossref
|
|
|
|
Miyadoh S (1997). Atlas of Actinomycetes. Asakura Publishing. Tóquio. 223p.
|
|
|
|
Monciardini P, Sosio M, Cavaletti L, Chiocchini C, Donadio S (2002). New PCR primers for the selective amplification of 16S rDNA from different groups of actinomycetes. FEMS Microbiol. Ecol. 42:419-429.
Crossref
|
|
|
|
Morris SJ, Blackwood CB (2007). The ecology of soil organisms. In: Paul EA (ed) Soil Microbiology, Ecology, and Biochemistry. Elsevier. Oxford. Pp. 195-229.
Crossref
|
|
|
|
Okoro CK, Brown R, Jones AL, Andrews BA, Asenjo JA, Goodfellow M, Bull AT (2009). Diversity of culturable actinomycetes in hyper-arid soils of the Atacama Desert, Chile. A. Van. Leeuw. 95(2):121-133.
Crossref
|
|
|
|
Parmar N, Dufresne J (2011). Beneficial Interactions of Plant Growth Promoting Rhizosphere Microorganisms. In: Singh A, Parmar N, Kuhad RC (eds) Bioaugmentation, Biostimulation and Biocontrol. Springer. Heidelberg. Pp. 27-42.
Crossref
|
|
|
|
Pereira JC, Neves MCP, Drozdowicz A (1999). Influence of antibiosis produced by actinomycetes on strains of Bradyrhizobium spp. on soybean nodulation. Pesq. Agropec. Bras. 34(1):99-108.
|
|
|
|
Pinheiro MS, Sousa JB, Bertini CHCM, Martins SCS, Martins CM (2014). Isolation and screening of rhizobial strains native from semi-arid tolerant to environmental stress. Encicl. Biosf. 10:2071-2082.
|
|
|
|
Pontes MMCM, Pontes Neto TP, Chaves LFC, Albuquerque SF, Oliveira JP, Figueiredo MVB (2012). Dual inoculation of β-Rhizobium and mycorrhiza at sabiazeiro seedlings for rehabilitation of degraded areas. Pesq. Agropec. Bras. 17:37-45.
|
|
|
|
Ramesh S, Mathivanan N (2009). Screening of marine actinomycetes isolated from the Bay of Bengal, India for antimicrobial activity and industrial enzymes. World. J. Microbiol. Biotechnol. 25(12):2103-2111.
Crossref
|
|
|
|
Ramos KA, Brito FAE, Nunes KJF, Martins CM, Martins SCS (2015). Characterization and crhomogenic diversity of actinobacteria from undisturbed microbial niche in the caatinga biome. Encicl. Biosf. 11:2115-2125.
|
|
|
|
Rao KR, Kiran CR, Rao DB, Madhavi Y, Rao PK, Rao TR (2012). Antagonistic activities of actinobacteria from mangrove sediment. Int. J. Pharm. Pharm. Sci. 4:364-367.
|
|
|
|
Rintala H, Nevalainen A, Rönkä E, Suutari M (2001). PCR primers targeting the 16S rRNA gene for the specific detection of streptomycetes. Mol. Cell. Probes. 15(6):337-347.
Crossref
|
|
|
|
Rousk J, Bååth E, Brookes PC, Lauber CL, Lozupone C, Caporaso JG, Knight R, Fierer N (2010). Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME. J. 4(10):1340-1351.
Crossref
|
|
|
|
Shinobu R (1958). Physiological and cultural study for the identification of soil Actinomycetes species. Mem. Osaka. Univ. B. Nat. Sci. 7:1-76.
|
|
|
|
Shirling EB, Gottlieb D (1966). Methods for characterization of Streptomyces species. Int. J. Syst. Bacteriol. 16:313-340.
Crossref
|
|
|
|
Shivlata L, Satyanarayana T (2015). Thermophilic and alkaliphilic Actinobacteria: biology and potential applications. Front. Microbiol. 6:1-29.
Crossref
|
|
|
|
Silva M S, Sales AN, Magalhães-Guedes KT, Dias DR, Schwan RFS (2013). Brazilian Cerrado Soil Actinobacteria Ecology. Biomed. Res. Int. 2013:1-10.
Crossref
|
|
|
|
Silva VMA, Lima JVL, Gondim PM, Martins CM, Martins SCS (2015). Effect of irrigation and type of cultivation on richness and diversity of chromogenic actinobacteria of soil from ceará semiarid region. Encicl. Biosf. 11:2965-2979.
|
|
|
|
Suneetha V, Zaved AK (2011). Actinomycetes: Sources for Soil Enzymes. In: Shukla G, Varma A (eds) Soil Enzymology. Springer. Heidelberg. Pp. 259-269.
|
|
|
|
Tarkka M, Hampp R (2008). Secondary Metabolites of Soil Streptomycetes in Biotic Interactions. In: Karlovsky P (ed) Secondary Metabolites in Soil Ecology. Springer. Heidelberg. Pp. 107-126.
Crossref
|
|
|
|
Tian-Xing L, Chao L, MingFu G (2011). Genetic diversity of antagonistic endophytic bacteria isolated from Sophora alopecuroide. J. Plant. Dis. Prot. 2:6-8.
|
|
|
|
Van Schreven DA (1964). The effect of some actomycetes on rhizobia and Agrobacterium radiobacter. Plant. Soil. 21:283-302.
Crossref
|
|
|
|
Vincent JM (1970). Manual for the practical study of root nodule bacteria. Blackwell Scientific Publications. Oxford. 176p.
|
|
|
|
Weisburg WG, Barns SM, Pelletier DA, Lane DJ (1991). 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 173:697-703
Crossref
|
|
|
|
Wink JM (2012). Compendium of actinobacteria. Braunschweig University of Technology. Braunschweig. Pp. 1-37.
|
|
|
|
Yao ZY, Kan FL, Wang ET, Wei GH, Chen WX (2002). Characterization of rhizobia that nodulate legume species of the genus Lespedeza and description of Bradyrhizobium yuanmingense sp. nov. Int. J. Syst. Evol. Microbiol. 52(6):2219-2230.
Crossref
|
|
|
|
Yu J, Zhang L, Liu Q, Qi X, Ji Y, Kim BS (2015). Isolation and characterization of actinobacteria from Yalujiang coastal wetland, North China. Asian. Pac. J. Trop. Biomed. 5(7):555-560.
Crossref
|
|
|
|
Zhu HH, Guo J, Yao Q, Yang SZ, Deng MR, Hanh VT, Ryan MJ (2007). Streptomyces vietnamensis sp. nov., a streptomycete with violet–blue diffusible pigment isolated from soil in Vietnam. Int. J. Syst. Evol. Microbiol. 57(8):1770-1774.
Crossref
|
|
|
|
Zilli JE, Campo RJ, Hungria M (2010). Effectiveness of Bradyrhizobium inoculation at pre-sowing of soybean. Pesq. Agropec. Bras. 45:335-338.
|