ABSTRACT
Jatropha curcas L. is an undomesticated plant species that has recently received great attention for its utilization in biofuel production in many countries including Botswana. The experiment investigated 30 J. curcas accessions, and the results revealed high significant differences (P<0.01) among accessions for peduncle length, number of seed per fruit, length of fruit stalk, fruit length, fruit width, seed length, seed width, seed weight, leaf length, leaf width, petiole length, total flower per cluster, female and male flower per cluster, and number of fruit per cluster. The qualitative characters revealed substantial amount of variability among the accessions except in fruit shape, stem color and flower color. Multivariate cluster analysis based on morphological characters and molecular characterization grouped the accessions into four clusters at dissimilarity coefficient of 1.37 and 0.85, respectively. The clustering based on morphological characters slightly matched that of groupings derived through molecular analysis with group A and B having some similar accessions. One Ghana accession used as reference was clearly separated from other accessions by both morphological and molecular markers, and also by seed weight and inflorescence compactness. Eight morphological characters and six molecular markers correlated positively with correlation coefficient values ranging from 0.56 to 0.66.
Key words: Characterization, genetic diversity, Jatropha curcas, molecular markers, morphological markers
Jatropha (Jatropha curcas L.) originated from Central America now found in both tropical and subtropical countries all over the world (Singh et al., 2010, Henning, 2009). More than 85% of J. curcas plantations are in Asia, chiefly Myanmar, India, China and Indonesia (Brittaine and Lutaladio, 2010). Africa accounts for only 12% or approximately 120,000 ha, mostly in Madagascar, Zambia, Tanzania and Mozambique (Renner et al., 2008). Latin America has approximately 20,000 ha of J. curcas plantation, mostly in Brazil (Brittaine and Lutaladio, 2010). Jatropha has been grown in the past mainly for producing oil for biofuels preferably in marginal environments in sub-Saharan Africa (Martin et al., 2019). The main problem of elevating J. curcas from a wild species to a profitable biodiesel crop is the low genetic and phenotypic variation of the accessions found in different regions of the world, hampering efficient plant breeding for productive traits (Montes et al., 2014).
Jatropha is an important and multipurpose plant with many attributes of considerable potential in bioenergy systems (Openshaw, 2000). The seed contain viscous, non-edible oil, which besides being a source of bio-diesel can also be used for manufacturing other useful products such as candle, high quality soap, cosmetics and can be used for healing several skin disorders (Openshaw, 2000). The seed cake can be used as organic fertilizer and feedstock for biogas production (Gavilanes et al., 2017; Staubmann et al., 1997).
J. curcas is known for its ability to survive in very poor dry soils in conditions considered marginal for agriculture, and can even root into rock crevices, but survival ability does not mean that high productivity can be obtained from growing the plant under marginal agricultural environments (Brittaine and Lutaladio, 2010). The studies conducted previously had indicated that there was morphological variation in J. curcas germplasm growing in different parts of the world. Evaluation of genetic diversity through morphological traits is direct, in-expensive and easy. However, morphological estimations are more dependent on the environment and are more subjective than other measurements (Ye et al., 2009). Morphological variability depends on a limited number of genes, and may not access much of the potential variability for the agronomic traits present in a crop (Mayes et al., 2009). Gohil and Pandya (2008)studied fourteen characters in Indian J. curcas accessions finding moderate genetic diversity and none of the morphological variables had heritability of over 75%. In another study of Indian accessions, Saikia et al. (2015)compared 34 sources, finding moderate variation in plant height, stem girth, branches per plant and seed weight.
The molecular markers are now important tools in plant genetic diversity analysis due to their sensitivity and specificity. The use of DNA-based markers has precedence in genetic characterization of J. curcas. In India and China, a wide variety of molecular marker systems have been used to assess intra-specific genetic diversity within species of J. curcas involving accessions from different agro-climatic zones (Singh et al., 2010). In this study, morphological and molecular markers were used to characterize J. curcas genotypes growing in Botswana that may serve as parental lines for breeding programs.
Experimental site and plant materials
The molecular work was done at Botswana University of Agriculture and Natural Resources (BUAN) Tissue Culture Laboratory, and the morphological work was field based at Sebele Agricultural Research Station Nursery in 2015 and 2016. The nursery was established in 2011 with 86 J. curcas accessions that were collected by National Plant Genetic Resources Center (NPGRC) from Southern and Northern parts of Botswana and 1 accession from Ghana. Thirty J. curcas accessions from the nursery were selected for the study.
Morphological data collection and analysis
Three trees per accession were randomly tagged from the 30 selected accessions, and morphological data was collected only from the tagged trees. The accessions were selected based on their phenotype and location. A total of 24 of both qualitative and quantitative traits were recorded following the National Bureau of Plant Genetic Resources (NBPGR) minimal descriptor list for J. curcas as developed by Sunil et al. (2010). The data collected was subjected to analysis of variance (ANOVA) using the SAS version 9.3 General Linear Model (GLM) package to determine the statistical differences on the traits for the given accessions.
Cluster analysis and principal coordinate analysis
The data for each morphological character was first transformed using the STAND procedure in Numerical Taxonomy System-pc (NTSYSpc version 2.1; Rohlf, 2002)in order to eliminate the effects of different scales of measurement. Then the unweighted pair group method with arithmetic averages (UPGMA) cluster analysis and principal coordinates analysis were performed with NTSYSpc version 2.1 (Rohlf, 2002). For cluster analysis, average linkage based on Euclidean distance was used to produce a dendrogram to show the similarities and differences between J. curcas accessions. For the principal coordinate analysis, the data matrix was standardized by row, a matrix of distance was computed and distance matrix was double–centered, the double centered matrix was then factored and a plot was made showing the accessions in a 3-dimensional space.
Molecular data collection
The young leaves were collected from the tagged trees in the field and placed in liquid nitrogen, transported to the laboratory and stored in a freezer. The DNA was extracted following the CTAB protocol. A total of twenty-two (22) SSR primers pairs developed by Wang et al. (2011) were tested with 30 J. curcas DNA sample for reproducible amplification. The PCR conditions for the microsatellite analysis were the touchdown PCR program: Initial denaturation 94°C for 2 min, Denaturation at 92°C for 30 s, annealing at 50°C for 30 s and extension at 72°C for 30 s, and the procedure was repeated for 16 cycles; followed by 18 cycles of denaturalization at 92°C for 30 s, annealing at 45°C for 30 s and extension at 72°C for 5 min; then one final extension step at 72°C for 5 min. The PCR products were separated on 3% agarose gel and stained with 2.5 μL of ethidium bromide. The gel documentation and visualization was done using the Bio-Rad Gel Doc systems XR+ with image Lab Software.
Molecular data analysis
The amplified bands for simple sequence repeats (SSR) markers were scored as present (1) or absent (0) for each accession by manual inspection. The data were tabulated in a matrix using Microsoft Excel v2010. Data analysis was performed using Numerical Taxonomic and Multi-variant Analysis System (NTSYSpc version 2.1). The genetic similarity coefficient of pair-wise comparisons among the J. curcas accessions analyzed were calculated based on Dice coefficient within the similarity for Qualitative data module of NTSYSpc version 2.1 (Rohlf, 2002). Clustering was performed using the generated coefficient according to the unweighted pair group mean algorithm (UPGMA) within the Sequential Agglomerative Hierarchical Nested analysis module. The dendrogram was drawn in the tree format using the tree plot module of NTSYSpc version 2.1 (Rohlf, 2002). The cophenetic correlation was estimated to measure goodness of fit among matrices generated using the matrix comparison module. The principal coordinated analysis was determined using the same procedure as for morphological markers above.
Characterization of Jatropha curcas accessions using morphological characters
There were significant differences (P<0.01) among the J. curcas genotypes in the characters that were analyzed, except for number of leaf lobes (Table 1). This indicated substantial amount of genetic variability among the traits analyzed for this set of accessions. The coefficient of variation (CV) revealed higher variations in the number of female flowers per cluster (42.90%), number of male flowers per cluster (40.52%), peduncle length (39.55), total flowers per cluster (38.75%), length of fruit stalk (35.40%) and number of fruit per cluster (33.00%). Lower coefficients of variation were shown in number of leaf lobes (9.13%), seed length (11.91%) and number of seed per fruit (13.00%) which indicates the small variation for these traits.
The J. curcas accessions differed in some qualitative characters (Table 2). All the accessions had a grey stem, cream yellow flower and an oval fruit shape. The pigmentation of emerging leaves was distributed as 71% dark greyed purple, 22.6% greyed purple and 6.4% red among the accessions. About 90.3% displayed brown pigmentation at the base of the petiole while only 9.7% displayed green petiole base. Inflorescence compactness was distributed as 77.4% compacted and 22.6% loose compact while the flowering occurrence was distributed as 51.6% flowered twice, 38.7% flowered once and only 9.7% had continuous flowering. The proportion of female flowers to male flower was distributed as 38.7% (2.1:11-20), 35.5% (3.1:>20) and 25.8% (1<1:10).
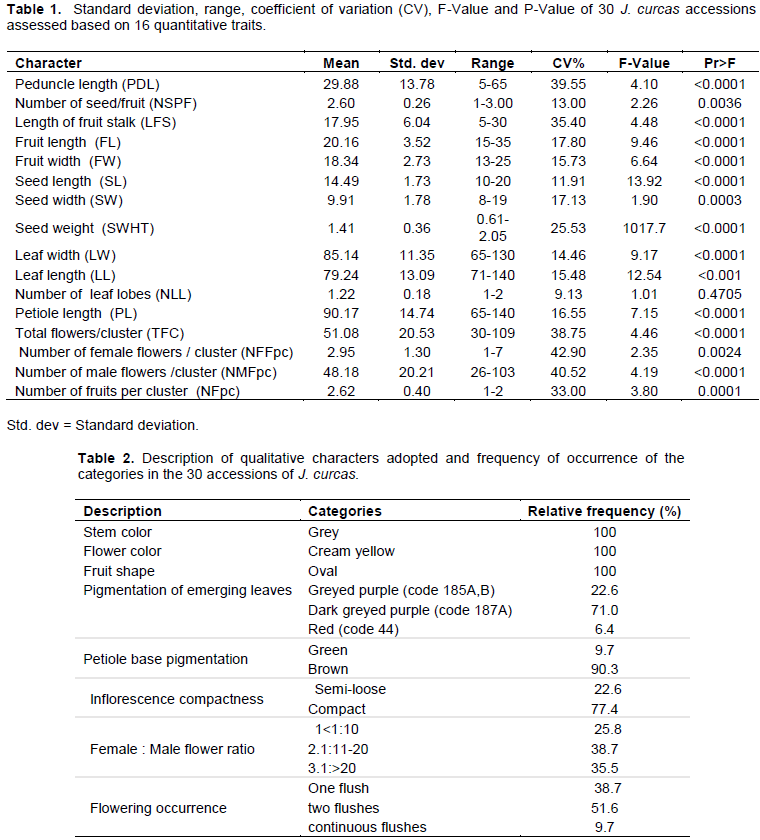
Morphological cluster analysis showed four clusters (A, B, C and D) at dissimilarity coefficient of approximately 1.37 (Figure 1). The first cluster (A) comprised of one accession which comes from Ghana (GJC1). The accession was separated from the rest of the accessions by seed weight and inflorescence compactness (Table 3).
Cluster B was the biggest with 23 accessions as follows: BJC5, BJC7, BJC13, BJC18, BJC14, BJC20, BJC51, BJC43, BJC65, BJC67, BJC57, BJC26, BJC33, BJC34, BJC17, BJC22, BJC10, BJC24, BJC25, BJC62, BJC19, BJC30 and BJC45. The major characters that led to the grouping were fruit stalk length, pigmentation on the emerging leaves, fruit length, fruit width, female to male flower ratio, pigmentation of peduncle base and number of seed per fruit. Within these cluster accession BJC14 and BJC20 were related with the distance coefficient of about 0.51.
Cluster C was the second largest with five accessions (BJC66, BJC68, BJC71, BJC75 and BJC79). The major characters that led to the grouping were female to male flower ratio, number of seeds per fruit, number of male flower per cluster and total flower per cluster. Cluster D comprises of one accession (BJC27) from the southern region and it was separated from the rest by leaf length, leaf width and petiole length.
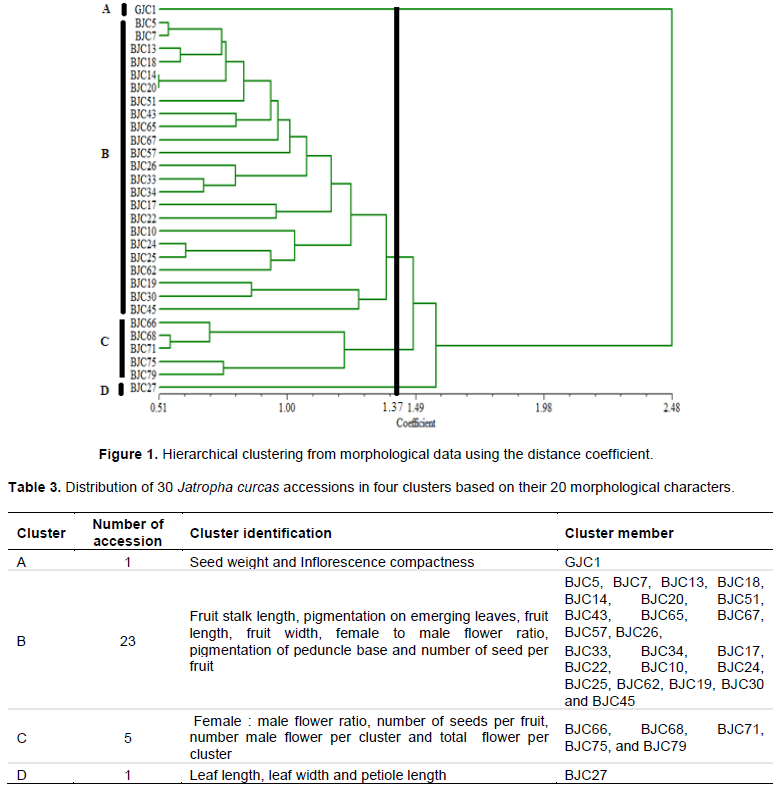
The Principal Coordinates Analysis (PCoA) revealed exactly the same four groupings obtained by cluster Analysis (Figure 2). The clusters were also designated by letter A, B, C and D. Cluster B being the biggest with 23 members followed by C with five accessions and lastly A and D comprising of only one accession each. The PCoA also clearly separated GJC1 (Ghana accession) from the rest of the accessions.
Characterization of Jatropha curcas accessions using Simple Sequence Repeat (SSR) markers
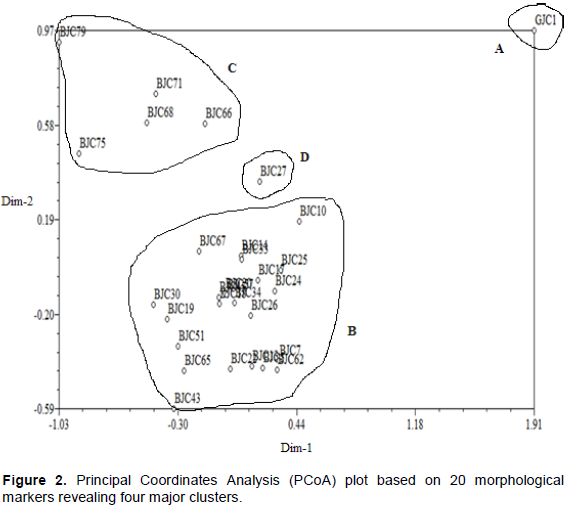
The genetic relationships among the 30 J. curcas accessions are shown in Figure 3. The amplitude in the similarity coefficient ranged from 0.71 to 1.00. The dendrogram obtained revealed four distinct clusters at 0.85 similarity coefficient, and grouping is from right to the left. The first cluster (A) comprised of one accession (GJC1 - Ghana). The second cluster (B) comprised of twenty-seven accessions (BJC5, BJC7,BJC68, BJC17, BJC79, BJC19, BJC20, BJC25, BJC27, BJC34, BJC45, BJC51, BJC57, BJC62, BJC24, BJC65, BJC33, BJC26, BJC71, BJC43, BJC67, BJC18, BJC30, BJC75, BJC66, BJC10 and BJC13) which are all Botswana accessions but collected from different villages in the north and south. Eleven accessions which are BJC17, BJC79, BJC19, BJC20, BJC25, BJC27, BJC34, BJC45, BJC51, BJC57 and BJC62 had the highest similarity coefficient of 1.00, which the used markers failed to distinguish them from one another and treated them as duplicates.
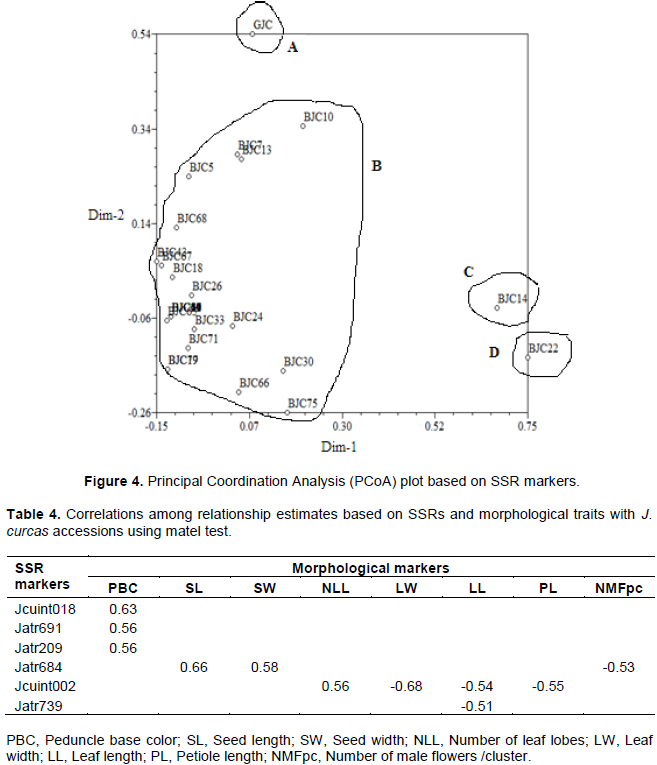
The third cluster (C) comprised of one accession (BJC14) which came from the northern region and the forth cluster (D) comprised also of one accession (BJC22) which came from the Southern region.
The Principal Coordinates Analysis (PCoA) revealed exactly the same four groupings obtained by cluster analysis (Figure 4). The clusters were also designated by letter A, B, C and D. Cluster B is the biggest with 27 members followed while cluster A, C and D are the smallest with one accession each.
Correlations between morphological and molecular markers
The correlation analysis showed correlation and no correlations between morphological and molecular markers. Eight morphological characters and six molecular markers showed moderate correlation (Table 4). The markers that showed positive correlation were peduncle base color with molecular marker Jcuint018 (0.63), Jatr691 (0.56) and Jatr209 (0.56). The seed size (length and width) correlated positively with molecular marker Jatr684 (0.66) and (0.58). The number of leaf lobes as well correlated positively with molecular marker Jcuint002 (0.56).
The negative correlations were observed between molecular marker Jcuint002 with leaf width (-0.68), leaf length (-0.54) and petiole length (-0.55). The leaf length showed a negative correlation with molecular marker Jatr739 (-0.51). The number of male flowers per cluster also correlated negatively with molecular marker Jatr684 (-0.53).
J. curcas is a polymorphic species with variations reported in tree architecture, plant height, number of fruits, seed dimensions (length, width, lateral diameter) and seed weight (Guan et al., 2013). In this study, the highest coefficient of variation was observed in the number of female flower per cluster, number of male flower per cluster, peduncle length, total of flowers, length of fruit stalk, number of fruits per cluster and seed weight. This indicates a wider variation existing among the accessions for the characters. Such wide variation in the species was not surprising because the characters are predominantly influenced by environment (Ovando-Medina et al., 2011). The trend has been reported previously by several authors for different characters in different population of J. curcas (Kaushik et al., 2007; Guan et al., 2013; Saadaoui et al., 2015). In addition, low coefficients of variation were revealed by number of leaf lobes, seed length, and number of seed per fruit and this indicated that a narrow variation existed among accessions for the characters. These traits with low coefficients of variations are more prone to highly hereditary traits, because of their uniformity.
The analysis of variance for quantitative characters also revealed high significant differences (P<0.01) in the thirty accessions. This indicated that substantial amount of genetic variability existed in the accessions. Saadaoui et al. (2015)investigated five characters of J. curcas, foliar surface area, leaf length, leaf width, petiole length and number of nodes, and all of them revealed high morphological variability (P<0.05) among the eight accessions in Tunisia. Similarly, Guan et al. (2013)found that five seed traits (seed weight, seed length, seed width, lateral diameter, seed length and width ratio) were significantly different among eight populations of J. curcas in China. The qualitative characters also revealed substantial amount of variability in the accessions. The pigmentation on emerging leaves was in line with what was observed by Pazeto et al. (2015) where the young leaves were distributed as red and purple. In a study conducted on 24 accessions of J. curcas collected from different zones of India, Kaushik et al. (2007)reported the variability in seed traits and oil content. There were significant differences (P<0.05) in seed size, 100-seed weight as well as in oil content between accessions. However, the coefficient of variation was higher for phenotype than genotype, indicating a predominant role of the environment. Similar results were found by Sunil et al. (2008a)who observed correlation between morphological characteristics (plant height, collar height and thickness, number of primary branches, petiole length, number of fruits per cluster, pedicel length and seed yield) with the oil content of the seed. Saikia et al. (2015)found moderate variation in plant height, stem girth, branches per plant and seed weight.
The clustering based on morphological characters slightly match that of groupings derived through molecular analysis, in that four groupings were produced by both methods, with group A and B having some similar accessions. The four groupings were confirmed by the Principal Coordinated analysis. For both morphological and molecular clustering, the highest group (B) comprised of accessions from the northern and southern parts of the country, suggesting that there is no relation between the place of origin and the clustering for the accessions. This also may be due to similar agro-climatic conditions or seed movement. The morphological markers revealed slightly higher genetic distance estimates ranging from 0.51 to 2.48 while the molecular marker showed relatively smaller genetic distance estimates ranging from 0.71 to 1.00. Cai et al. (2010)investigated the genetic diversity of 219 J. curcas accessions from China using Simple Sequence Repeat (SSR) markers and revealed a low genetic diversity in the Chinese germplasm. Pamidimarri et al. (2009a)isolated SSR markers, investigated the genetic diversity of J. curcas accessions from India, and found a narrow genetic diversity in accessions. Also, low polymorphism was found in 64 genotypes from five geographic locations (Brazil, Cape Verde, Cuba, Mozambique and Senegal) using 32 SSR markers (Ricci and Chekhovskiy, 2012). Ambrosi et al. (2010)analyzed 26 accessions from different geographical regions (including Mexico, South America, Asia and Africa), using 10 RAPD, 6 ISSR and 10 SSR markers, and low genetic variability was documented not only among accession groups but also among accessions of different geographical origin, with the exception of Mexican landraces. Tanya et al. (2011)characterized 26 Mexican, 3 Chinese, 3 Thai and 4 Vietnamese accessions using SSR markers, and five of these loci clearly displayed distinct banding patterns between 26 Mexican accessions (non- toxic) and the 10 Asian accessions (toxic). The slight difference between clustering based on molecular markers and morphological markers may be that most of the characters are controlled by polygenes and are highly influenced by environment (Behera et al., 2012). However, the SSR markers and the morphological markers cluster analysis clearly separated the Ghana accession (GJC) and the Botswana accessions. The characters that separated the Ghana accession from others were seed weight and inflorescence compactness, which suggests that the Ghana accessions could be valuable in the improvement of the genetic materials found in Botswana, as it can be used to improve the seed yield of the local accessions.
The authors have not declared any conflict of interests.
REFERENCES
|
Ambrosi DG, Galla G, Purelli M, Barbi T, Fabbri A, Lucretti S, Sharbel TF, Barcaccia G (2010). DNA markers and FCSS analyses shed light on the genetic diversity and reproductive strategy of Jatropha curcas L. Diversity 2:810-836.
Crossref
|
|
|
|
Behera TK, Gaikwad AB, Swati S, Bharadwaj C, Munshi AD (2012). Morphological and Molecular analyses define the genetic diversity of Asian bitter gourd (Momordica charantia L.). Australian Journal of Crop Science 6(2):261-267.
|
|
|
|
|
Brittaine R, Lutaladio N (2010). Jatropha: a smallholder bioenergy crop: the potential for pro-poor development. Integrated Crop Management 8:114.
|
|
|
|
|
Cai Y, Sun D, Wu G, Peng J (2010). ISSR-based genetic diversity of Jatropha curcas germplasm in China. Biomass and Bioenergy 34:1739-1750.
Crossref
|
|
|
|
|
Gavilanes FZ, Guedes CLB, Silva HR, Nomura RG (2017). Physic Nut Seed Cake Methanation and Chemical Characterization of Anaerobic Bio-digested Substrate. Waste and Biomass Valorization. 10.1007/s12649-017-0148-y.
Crossref
|
|
|
|
|
Gohil RH, Pandya JB (2008). Genetic diversity assessment in physic nut (Jatropha curcas L.). International Journal of Plant Production 2(4):321-326.
|
|
|
|
|
Guan J, Yu H, Zhang J, Yang R, Fan Y (2013). Study on Seed Morphology and Genetic Diversity of Jatropha curcas L. from Different Provenances. Advance Journal of Food Science and Technology 5:169-173.
Crossref
|
|
|
|
|
Henning RK (2009). The Jatropha System An integrated approach of rural development. Evolution and Human Behaviour 18:375-386.
|
|
|
|
|
Kaushik N, Kumar K, Kumar S, Kaushik N, Roy S (2007). Genetic variability and divergence studies in seed traits and oil content of Jatropha (Jatropha curcas L.) accessions. Biomass and Bioenergy 31:497-502.
Crossref
|
|
|
|
|
Martin M, Bohlinger B, Senger E, Dongmeza E, Andrianirina ZT, Montes JM (2019). Genetic Improvement of Edible and Non-edible Jatropha for Marginal Environments in Sub-Saharan Africa. In: Mulpuri S, Carels N, Bahadur B (eds) Jatropha, Challenges for a New Energy Crop. Springer, Singapore, pp. 3-27.
Crossref
|
|
|
|
|
Mayes S, Basu S, Murchie E, Roberts JA, Aam-Ali SN, Stadler F, Mohler V, Wenzel G, Massawe F, Killian A, Bonin A, Beena A, Sheshshayee MS (2009). BAMLINK- A Cross disciplinary programme to enhance the role of bambara groundnut (Vigna subterranea L. Verdc.) for food security in Africa and India. Acta Hortculturae 806:137-150.
Crossref
|
|
|
|
|
Montes Osorio LR, Torres Salvador AF, Jongschaap RE, Azurdia Perez CA, Berduo Sandoval JE, Trindade LM, Visser RG, van Loo EN (2014). High level of molecular and phenotypic biodiversity in Jatropha curcas from Central America compared to Africa, Asia and South America. BMC Plant Biology 25:14-77.
Crossref
|
|
|
|
|
Openshaw K (2000). A review of Jatropha curcas: an oil plant of unfulfilled promise. Biomass and Bioenergy 19:1-15.
Crossref
|
|
|
|
|
Ovando-Medina I, Sánchez-Gutiérrez A, Adriano-Anaya L, Espinosa-García F, Núñez-Farfán J, Salvador-Figueroa M (2011). Genetic Diversity in Jatropha curcas Populations in the State of Chiapas, Mexico. Diversity 3(4):641-659.
Crossref
|
|
|
|
|
Pamidimarri DVNS, Sinha R, Kothari P, Reddy MP (2009a). Isolation of novel microsatellites from Jatropha curcas L. and their cross-species amplification. Molecular Ecology Resources 9:431-433.
Crossref
|
|
|
|
|
Pazeto MSR, Unêda-trevisoli SH, Corrêa AAP, Vianna VF, Leite DC, Di Mauro AO (2015). Genetic diversity in Jatropha species from different regions of Brazil based on morphological characters and inters-simple sequence repeat (ISSR) molecular markers. African Journal of Biotechnology 14:2066-2079.
Crossref
|
|
|
|
|
Renner A, Zelt T, Gerteiser S (2008). Global Market Study on Jatropha Final Report. Berlin, Germany.
|
|
|
|
|
Ricci A, Chekhovskiy K (2012). Molecular Characterization of Jatropha curcas Resources and Identification of Population-Specific Markers. Bioenergy Research 5:215-224.
Crossref
|
|
|
|
|
Rohlf FJ (2002). NTSYS-pc, Numeral taxonomy and multivariate analysis system. Version 2.1. Department of Ecology and Evolution. State University of New York, Stony Brook, NY.
|
|
|
|
|
Saadaoui E, Martin JJ, Bouazizi R, Romdhane CB, d Grira M, Abdelkabir S, Khouja ML, Cervantes E (2015). Phenotypic variability and seed yield of Jatropha curcas L. introduced to Tunisia. Acta Botanica Mexicana 110:119-134.
Crossref
|
|
|
|
|
Saikia SP, Mapelli S, Breviario D, Galasso I, Giani S, Braglia L, Pecchia P, Gogoi A, Mudoi KD (2015). Comparative studies for selection of Jatropha curcas L. capable of high yield and oil quality in Assam environment. Current Science 109(3):552-566.
|
|
|
|
|
Singh P, Singh S, Mishra SP, Bhatia SK (2010). Molecular Characterization of Genetic Diversity in Jatropha curcas L . Genes, Genomes and Genomics 4:1-8.
|
|
|
|
|
Staubmann R, Foidl G, Foidl N, Gubitz GM, Lafferty RM, Arbizu VM, Steiner W (1997). Biogas production from Jatropha curcas press-cake. Applied Biochemistry and Biotechnology 63-65:457-467.
Crossref
|
|
|
|
|
Sunil N, Kumar V, Varaprasad KS, Rao GR, Sujatha M, Sharma SK (2010). Minimal descriptors of Jatropha curcas L. National Bureau of Plant Genetic Resources, Pusan Campus, New Delhi viii: 32p.
|
|
|
|
|
Sunil N, Varaprasad KS, Sivaraj N, Suresh-Kumar T, Abraham B, Prasad RBN (2008a). Assessing Jatropha curcas L. germplasm in-situ-A case study. Biomass Bioenergy 32:198-202.
Crossref
|
|
|
|
|
Tanya P, Dachapak S, Tar MM, Srinives P (2011). New microsatellite markers classifying nontoxic and toxic Jatropha curcas. Genetics 90:76-78.
Crossref
|
|
|
|
|
Wang CM, Liu P, Yi C, Gu K, Sun F, Li L, Lo LC, Liu X, Feng F, Lin G, Cao S, Hong Y, Yin Z, Yue GH (2011). A first generation microsatellite- and SNP-based linkage map of Jatropha. PLoS One 6(8):e23632.
Crossref
|
|
|
|
|
Ye M, Li C, Francis G, Makkar HPS (2009). Current situation and prospects of Jatropha curcas as a multipurpose tree in China. Agroforestry Systems 76:487-497.
Crossref
|
|