Full Length Research Paper

ABSTRACT
One of the strategies of Integrated Pest Management (IPM)-soybean is the use of genetically modified plants. However, there are concerns about the unknown effects associated with this technology on non-target organisms. The objective of this study was to determine the population dynamics of species of floral-visiting insects in Bacillus thuringiensis (Bt) and non-Bt soybean. In the experiment, two soybean treatments were assessed, one of soy DM 6563 Intacta and the other BMX Potência RR. Floral-visiting insects were collected in the flowering period every 15 min/h for 12 h/day, every 3 to 4 days. 549 floral-visiting insects (Class Insecta) were collected, comprising 8 orders, 30 families, and 92 species. Of these, 279 were present in the cultivar DM 6563 Intacta and 270 in the cultivar BMX Potência. The most abundant species were Apis mellifera (35.15%), Musca species 1 (10.01%), and Lagria villosa (5.28%). Species composition was similar for the two cultivars. The highest number of species visited the flowers of the cultivars between 8:00 and 11:00 am, while the lowest intensity of visitation occurred between 12:00 and 15:00 pm. After diversity was determined, it was observed that cultivar BMX Potência (non-Bt) had a higher diversity than cultivar DM 6563 Intacta (Bt).
Key words: Pollinators, species diversity, transgenic.
INTRODUCTION
Soybean culture has been affected by many agricultural pests, including arthropods, resulting in decreased crop yields and reduced seed and grain quality (Roggia, 2010). Frequent pest infestations have resulted in increased use of synthetic chemical insecticides in all producing regions in Brazil. Currently, the harmful effects of insecticides on pollination by entomophilous agents, an important production factor of this agricultural crop, are of concern (Freitas and Pinheiro, 2012; Malaspina and Silva-Zacarin, 2006; Spadotto et al., 2004). Besides increasing the number of fruits or pods, effective pollination can also contribute to increased oil in the seeds, number of seeds per pod, seed weight, shortened crop cycles, uniform seed ripening, and reducing crop losses (Free, 1993; Freitas, 1997; Nogueira-Couto, 1998). To reduce risks to human health, environmental contamination, and possible harmful effects on pollinating insects, including bees, the restricted use of insecticides in soybean culture has been encouraged (Anvisa, 2015). To minimize consequences from insect pests in soybean culture and regulate the use of pesticides, the Integrated Pest Management of soy (IPM-Soy) was implemented, integrating the use of various strategies and control tactics (Corrêa-Ferreira et al., 2010), one of which is the use of genetically modified plants (transgenic) resistant to pests. Currently, available transgenic plants resistant to caterpillars contain genes of the bacterium Bacillus thuringiensis (Bt) that encode lethal toxins for certain insect groups. Bt plants have the potential to minimize losses caused by insect pests, especially the order Lepidoptera, as well as reduce insecticide use (Yu et al., 2011). Bt insecticidal proteins kill insects by a process comprising the following steps: crystal solubilization processing, conversion to an active protoxin form, high-affinity binding to the midgut receptors, irreversible insertion of the toxin into the membrane, and the formation of infection pores (Bravo et al., 2005; de Maagd et al., 2001).
Despite numerous advantages of using genetically modified insect-resistant plants, there are still concerns about the unknown effects associated with this technology (Kouser and Qaim, 2011; Nunes, 2010). Several studies show that the effect this technology may have on non-target organisms is still unclear (Faria et al., 2007; Liu et al., 2005; Naranjo, 2005; Silva, 2013; Whitehouse et al., 2007). Thus, the large-scale use of genetically modified crops for resistance to pests can pose a risk to biodiversity as a result of possible effects on non-target organisms (Dutra et al., 2012), which underscores the importance of conducting studies that assess such interactions.
Non-target organisms are defined as species that may be exposed to Bt proteins for a long time, but which are not direct targets of transgenic technology (Andow and Hilbeck, 2004). Among these organisms, one of the most important groups are the pollinating insects that may be both directly exposed to the toxins of these plants, through consumption of the plant, nectar, or pollen (Nunes, 2010), as indirectly through the food chain when arthropods feed on herbivores or honeydew from insects that have fed off transgenic plants (Groot and Dicke, 2002; Faria et al., 2006).
There is evidence that the expression of Bt toxins in transgenic plants can affect (directly or indirectly) the populations of non-target species (Birch, 1997; Monnerat and Bravo, 2000) by reducing the populations of pollinating insects and floral visitors, including beetles, butterflies, and a number of beneficial arthropods, which can lead to reduced biological pest control (Hong et al., 2008). In Bt maize that expresses the Cry1Ab toxin, as does the soybean used in the present study, a negative effect on the non-target species Spodoptera littoralis (Boisduval) (Lepidoptera: Noctuidae) and its larval parasitoid Cotesia marginiventris (Cresson) (Hymenoptera: Braconidae) was determined (Vojtech et al., 2005). Similar negative effects were also observed on the natural enemy Pirata subpiraticus (Araneae: Lycosidae) in cultures of transgenic rice expressing the same protein Cry1Ab (Chen et al., 2009). Other studies also show that the survival of bees feeding on pollen of Bt cotton expressing this toxin was negatively affected (Liu et al., 2000). Therefore, the objective of the current study was to determine the characteristics of the population dynamics of species of floral-visiting insects in Bt and non-Bt soybean Glycine max (L.) Merrill cultivars.
MATERIALS AND METHODS
Experimental area
The experiment was conducted at the experimental farm of the Federal University of Grande Dourados, Dourados-MS, Brazil (22°14′ 20.51″south, 54° 59′M 58.4″ west; altitude 394 m). The studied areas comprised two fields of 1.0 ha each. One field was sown with the cultivar DM 6563 Intact (MON 87701 × MON 89788) and the other with the cultivar BMX Potência RR (M 8360 RR). Both cultivars, which have a similar phenological cycle, were sowed on 5 November, 2014.
Survey of floral visitors
The collection of floral visitors took place throughout the flowering period (26 December, 2014 to 11 January, 2015) of the cultivars. The sampling method was adapted from that of Pires et al. (2006a). The collection of floral visitors was done to determine the abundance and diversity of these insects in the Bt and non-Bt soy, as well as visitation characteristics and species composition for each period in both cultivars.
Four evaluations were carried out every three or four days and in each assessment. Active insect collection was carried out every 15 min for 12 h in each cultivar. Insects were collected using an insect net or directly in a deadly vial. All insects found resting on or taking off from flowers were captured. After the 15 min of collection, the dead insects were placed in an envelope, stating date/time, collector, and treatment and then stored in a styrofoam box to be taken to the Entomology Laboratory of the Federal University of Grande Dourados (UFGD) for later sorting.
Identification of floral visitors
The insects were identified to the lowest taxonomic level possible using specific keys for the groups and confirmed with experts in the field.
Statistical analysis
To analyze the diversity of the two communities studied, the Shannon-Wiener function was calculated (Shannon and Weaver, 1949), as well as Simpson’ index (Pinto-Coelho, 2002). To obtain representative gradients of the floral visitors community structure, based on the species composition at each time and day of collection, the normality of the data of all individuals of each species in each day and time of collection, Kolmogorov-Smirnov, and Shapiro-Wilk tests were used. Then, comparative analysis was performed between the different schedules and treatments using the Kruskal-Wallis test.
RESULTS
Both cultivars flowered for approximately 15 days. During flowering, 549 species of floral-visiting insects (Class Insecta) were collected, comprising 8 orders, 30 families, and 92 species. Of this total, 279 individuals (50.81%) were present in the Bt cultivar, whereas 270 (49.18%) were collected from flowers of non-Bt cultivar (Table 1). No significant differences were noted in the abundance of insect species in relation to the cultivars, as similar numbers of individuals were observed in both cultivars. The Hymenoptera, to which the largest number of collected specimens belonged, showed the highest abundance among all of the orders collected, both in the Bt cultivar where 122 individuals were sampled (43.72% of the total found in this cultivar), as well as in the non-Bt cultivar that presented 121 individuals of this order (44.81%). Among the specimens from this order collected, representatives of the Apidae family were the most abundant, representing 40.86 and 38.14%, respectively, of all insects found in the Bt and non-Bt cultivars, totaling 9 species in both cultivars, and Apis mellifera was the most abundant.
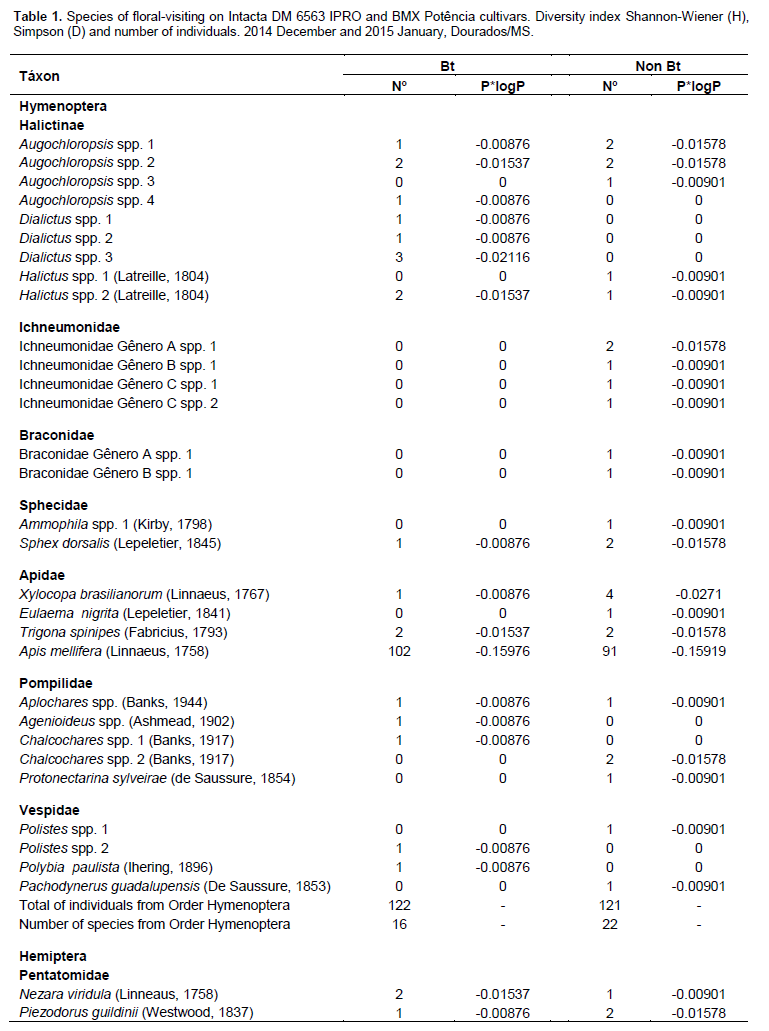
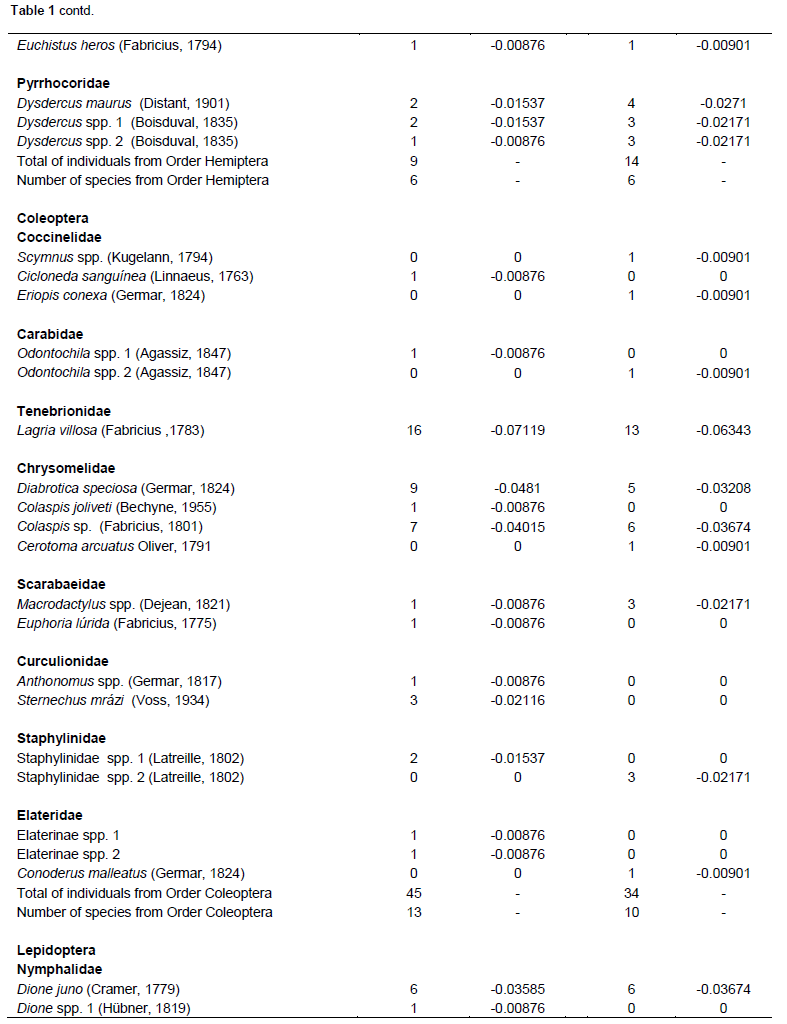
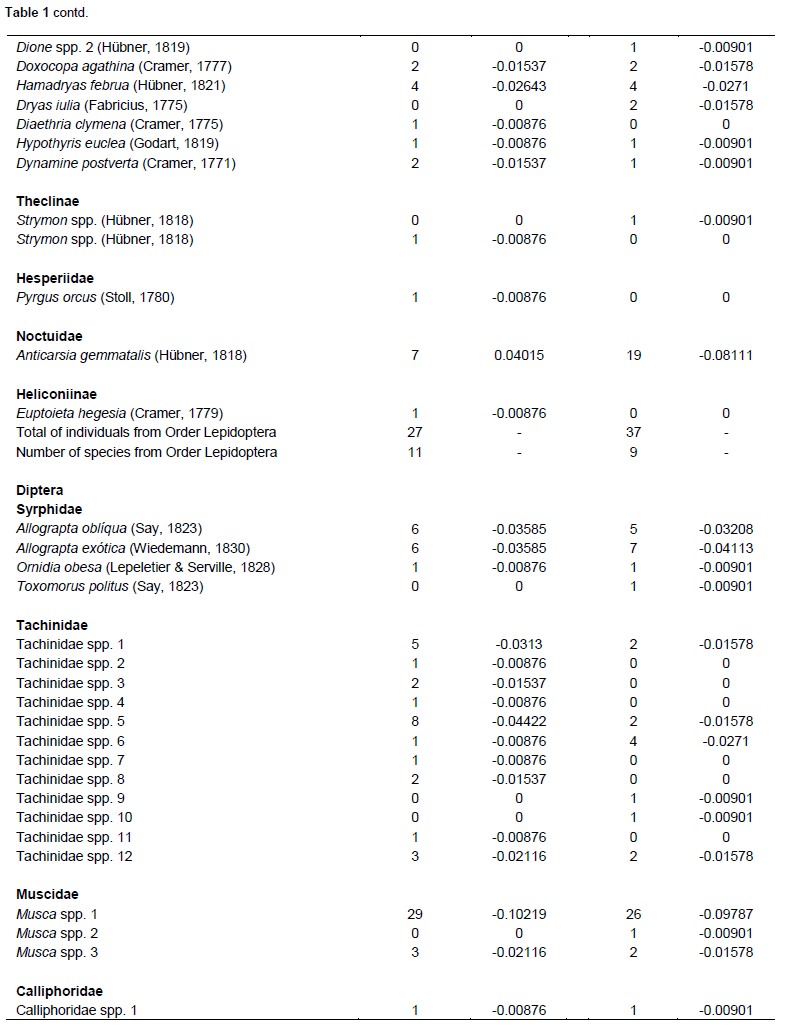
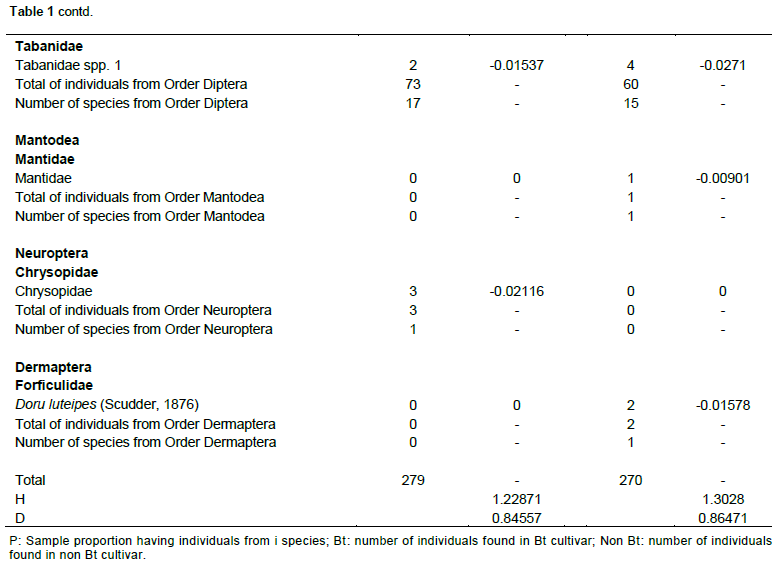
Diptera had the highest diversity, with 15 species in the non-Bt cultivar and 17 species observed in the Bt cultivar, with Hymenoptera having 22 species in the Bt cultivar and 16 in the non-Bt cultivar. Coleoptera had 10 species in the Bt cultivar and 13 species in the non-Bt cultivar and Lepidoptera had 9 in the Bt cultivar and 11 species in the non-Bt cultivar. Hemiptera had six species in both cultivars, while Neuroptera, Mantodea, and Dermaptera had only one species each. Although, the distribution of species was very similar between the two evaluated cultivars, the results of the Shannon-Wiener and Simpson tests demonstrated that the Bt cultivar had greater diversity (H = 1.30280; D = 0.86471) than the non-Bt cultivar (H = 1.22871; D = 0.84557).
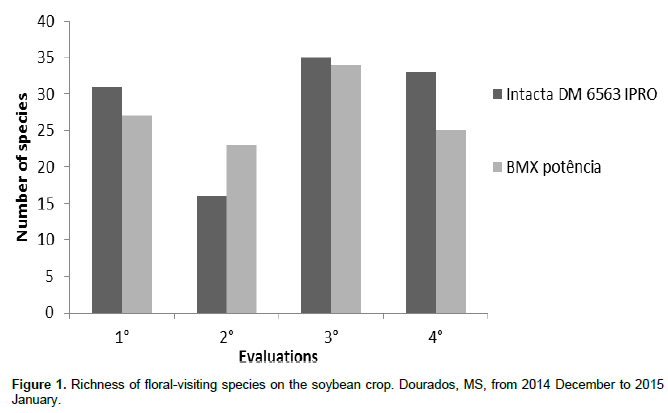
The greatest number of species was observed in the third evaluation on January 6 with a total of 34 species in the BMX Potência and 35 in the DM 6563 IPRO (Figure 1). The first (December 30) and the fourth (9 of January) evaluations had nearly equal number of species, while at the second evaluation (02 January) was noticed the lowest value of the species found during the period of sampling.
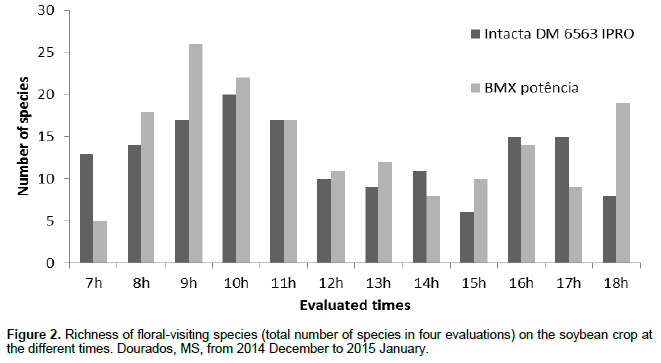
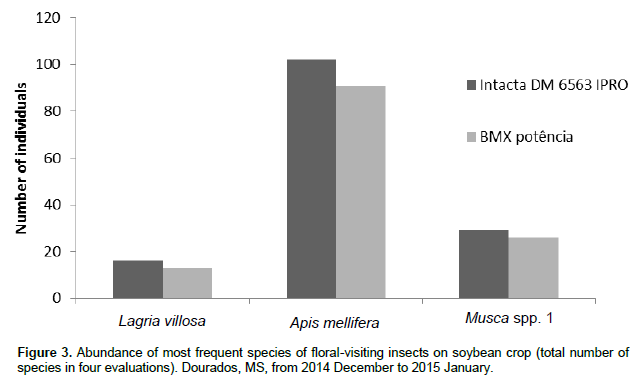
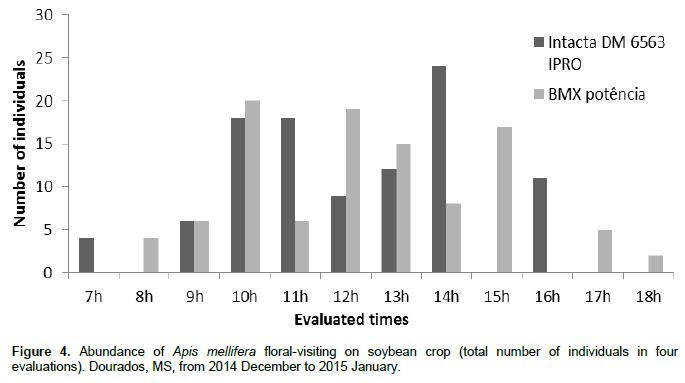
The greatest number of floral visitor species in the soybean cultivars occurred between 8:00 and 11:15 am. The least species visitation to the soybean flowers occurred between 12:00 and 15:15 pm (Figure 2). Among the 103 species collected in the two studied soybean cultivars, A. mellifera L. had the highest number of individuals (102 individuals in the cultivar Intacta and 91 in the cultivar BMX Potência), representing 36.55 and 33.70%, respectively, of the total insects found in both crop cultivars (Figure 3). The total number of individuals of A. mellifera observed during the period of the evaluated day (from 07:00 am to 18:15 pm) (Figure 4) did not indicate normality by the Kolmogorov-Smirnov and Shapiro-Wilk tests, that evaluated block (evaluation days) and treatment (evaluated times). Therefore, the nonparametric Kruskal-Wallis test (α = 5%) was performed. The variation in the abundance of A. mellifera in the different classes of times was determined and no significant differences were noted between the classes of times in both cultivars.
DISCUSSION
Because there was no significant difference in the abundance and richness of floral-visiting insects between the two treatments of G. max, this group of insects is therefore not directly affected by the Bt toxin expressed in the evaluated cultivar (Intacta). Higgins (2009) examined non-target arthropods at the community level in corn in the United States for three years, and did not observe a significant difference in the abundance of the community, when fields of Bt and non-Bt corn were compared.
In the present study, the Shannon-Wiener diversity (H') index showed similar values in both evaluated cultivars. These close results between Bt and non-Bt cultivars represent equality in richness and abundance in relation to species among the evaluated cultivars. However, lower species richness was initially expected for the Bt cultivar, owing to a higher degree of environmental disturbance caused by the Cry1Ac toxin, since less altered environments tend to have greater species richness (Odum, 1988; Freitas and Pinheiro, 2012). Nonetheless, in non-Bt cultivars or tolerant insects, the generally required use of insecticides can also provide a disturbance of the environment.
Considering the Hymenoptera collected, representatives of the Apidae family were the most abundant. A. mellifera was the most abundant. Similar results were also observed in Mato Grosso do Sul, Brazil, regarding the diversity of floral-visiting insects in cotton crops (Dutra et al., 2012), as well as in other regions of Brazil, such as Bahia, Goiás, Mato Grosso, São Paulo, and in Distrito Federal (Pires et al., 2006b).
The high abundance of A. mellifera can be attributed to this species visiting flowers in exchange for offered floral resources (Machado, 2006; Dutra et al., 2012); the peak of visitors during the period between 08:00 and 11:15 am is possibly due to higher production and resources offered to insects by the soybean plants to attract pollinators to flowers. This species is an important pollinator, very common, and has the highest number of interactions with plants (Mouga et al., 2012). The presence of these bees is fundamental for soybean production since it positively influences the production of grains and seed quality due to its pollination efficiency of 97.43% when compared with that of other insects (Chiari et al., 2008).
An important disturbance called Colony Collapse Disorder (CCD) has been the focus of many studies globally due to the disappearance of individuals that form bee colonies, as seen by the absence of dead bees in apiaries, the rapid decrease in the number of adult bees, and the lack of cleptoparasites (Cox-Foster and van Engelsdorp, 2009). One of the most important probable causes of this disorder is the effect of chemical insecticides on or near the colonies. Therefore, in efforts to protect bee species, mainly A. mellifera, from CCD, the data obtained in this study regarding the time of least visitation to the flowers, can determine the most sustainable insecticide applications, when necessary.
Based on the results of the current study, it can be concluded that the Cry1Ac toxin from Bt did not affect the population dynamics of the species of flower-visiting insects in soybean crops. Therefore, it is possible to add that this technology led to no significant differences in diversity, species composition, frequency, richness, or time of occurrence among the species of floral visitors to the culture.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENT
The authors would like to thank FUNDECT/CAPES, chamada nº 44/2014-PAPOS-MS/Manoel A. Uchoa-Fernandes.
REFERENCES
|
Andow DA, Hilbeck A (2004). Science-based risk assessment for non-target effects of transgenic crops. BioScience. 54:637-649. |
|
|
Agencia Nacional de Vigilância Sanitária (2015). ANVISA. |
|
|
Birch RG (1997). Plant transformation: Problems and Strategies for Practical Application. Ann. Rev. Plant Physiol. Plant Mol. Biol. 48:297-326. |
|
|
Bravo A, Gill SS, Soberon M (2005). Bacillus thuringiensis mechanisms and use. In: Gilbert LI, Iatrou K, Gill SS (eds.). Compr. Mol. Insect Sci. Elsevier. 6:175-206. |
|
|
Chen M, Liu ZC, Frag Q, Hu C, Peng YF, Shelton AM (2009). Analysis of Cry1Ab toxin bioaccumulation in a food chain of Bt rice, an herbivore and a predator. Ecotoxicology 18:230-238. |
|
|
Chiari WC, Toledo VAA, Hoffmann-Campo CB, Rúvolo-Takasusuki MCC, Toledo TCSOA, Lopes TS (2008). Polinização por Apis mellifera em soja transgênica [Glycine max (L.)] Roundup Ready ™ cv. BRS 245 RR e convencional cv. BRS 133. Acta Sci. Agron. 30:267-271. |
|
|
Corrêa-Ferreira BS, Alexandre TM, Pellizzaro EC, Moscardi F, Bueno AF (2010). Práticas de manejo de pragas utilizadas na soja e seu impacto sobre a cultura. Londrina:Embrapa Soja. Circ. Técnica.78:15. |
|
|
Cox-Foster D, vanEngelsdorp D (2009). Solving the mystery of the disappearing bees. Scientific American, New York. |
|
|
de Maagd RA, Bravo A, Crickmore N (2001). How Bacillus thuringiensis has envolved specific toxins to colonize the insect world. Trends in Genetics. 17:193-199. |
|
|
Dutra CC, Meotti C, Fernandes MG, Raizer J (2012). Riqueza e composição de insetos visitantes florais em algodoeiro Bt e não Bt. Arq. Inst. Biol. 79:353-361. |
|
|
Faria CA, Wackers FL, Pritchard J, Barrett DA, Turlings TCJ (2007). High susceptibility of Bt maize to aphids enhances the performance of parasitoids of lepidopteran pests. PLoS One. 7:1-11. |
|
|
Faria MF, Lundgren JG, Fontes EMG, Fernandes OA, Schimidt F, Nguyen VT, Andow DA (2006). Assessing the effects of Bt Cotton on Generalist Arthropod Predator. In: Hilbeck, A., Andow, D.A., Fontes, E.M.G. (ed). Environmental risk assessment of genetically modified organisms methodologies for assessing Bt cotton in Brazil. CABI Publishing pp. 175-199. |
|
|
Free JB (1993). Insect pollination of crops. London: Academic Press. |
|
|
Freitas BM (1997). Changes with time in the germinability of cashew (Anacardium occidentale) pollen grains found on different body areas of its pollinator bees. Rev. Bras. Biol. São Carlos 57:289-294. |
|
|
Freitas BM, Pinheiro JN (2012). Polinizadores e pesticidas: princípios e manejo para os agroecossistemas brasileiros. Brasília: MMA. P 112 |
|
|
Groot AT, Dicke M (2002). Insect-resistant transgenic plants in amultitrophic context. Plant J. 31:387-406. |
|
|
Higgins LS, Babcock J, Neese P, Layton RJ, Moellenbeck D, Storer N (2009). Three-year field monitoring of Cry1F, Event DASØ15Ø7–1, maize hybrids for non-target arthropod effects. Environ. Entomol. 38:281-292. |
|
|
Hong LTT, Tâm DQ, Tuân HA (2008). Potencial effects of transgenic Cotton on flower visitors in Vietnam. In: Andow, DA, Hillbeck A, Van Tuat AN (Ed.). Environmental Risk Assessment of Genetically Modified Organisms: challenges and opportunities with Bt cotton in Vietnam. Wallingford: Centre Agric. Biosci. Int. 9:432. |
|
|
Kouser S, Qaim M. (2011). Impact of Bt cotton on pesticide poisoning in smallholder agriculture: A panel data analysis. Ecol. Econ. 70:2105-2113. |
|
|
Liu XX, Sun CG, Zhang QW (2005). Effects of transgenic Cry1A+CpTI cotton and Cry1Ac toxin on the parasitoid, Campoletis chlorideae (Hymenoptera: Ichneumonidae). Insect. Sci. 12:101-107. |
|
|
Liu W, Wang F, Zhang F, Meng Z, Wang F (2000). Evaluation on role of predators in Helicoverpa armigera control. Chinese Journal of Biological Control. 16:97-101. |
|
|
Machado CS (2006). Aspectos de interesse da polinização entomófila de Helianthus annus L. no Recôncavo Baiano. Dissertação de mestrado. Universidade Federal da Bahia. Cruz das Almas, Bahia. |
|
|
Malaspina O, Silva-Zacarin ECM (2006). Cell markers for ecotoxicological studies in target organs of bees. Braz. J. Morphol. Sci. 23:303-309. |
|
|
Monnerat RG, Bravo A (2000). Proteínas bioinseticidas produzidas pela bactéria Bacillus thuringiensis: modo de ação e resistência. In: Melo IS, Azevedo JL (Ed.). Controle biológico 3. Jaguariúna: Embrapa Meio Ambien¬te.163-200. |
|
|
Mouga DMDS, Noble CF, Bussmann DBG, Krug C (2012). Bees and plants in a transition area between atlantic rain forest and araucaria forest in southern Brazil. Revue d'Ecologie. 67:313-327. |
|
|
Naranjo SE (2005). Long-Term Assessment of the Effects of Transgenic Bt Cotton on the Abundance of Nontarget Arthropod Natural Enemies. Environ. Entomol. 34:1193-1210. |
|
|
Nogueira-Couto RH (1998). Manejo das colméias de abelhas africanizadas para polinização. In: XII Congresso Brasileiro de Apicultura. Salvador: Confederação Brasileira de Apicultura. 2:129-134. |
|
|
Nunes DH (2010). Efeitos do algodoeiro geneticamente modificado Bollgard® em organismos não-alvo. Tese Doutorado em Ciências. Universidade de São Paulo, Piracicaba. 211f. |
|
|
Odum EP (1988). Ecologia. Ed. Guanabara, Rio de Janeiro. |
|
|
Pinto-Coelho RM (2002). Fundamentos em Ecologia. Porto Alegre, Ed:Artmed. |
|
|
Pires C, Silveira, FA, Pereira FO, Paes JS, Sujii ER, Fontes E (2006a). Protocolo de amostragem de visitantes florais em algodoeiro (Gossypium spp.). Brasília: Embrapa Recursos Genéticos e Biotecnologia, Boletim de Pesquisa e desenvolvimento P 131. |
|
|
Pires C, Silveira F, Cardoso CF, Oliveira GM, Pereita FFO, Souza VV, Nakasu EYT, Paes JSO, Teles É, Silvie P, Rodrigues S, Miranda J, Scomparini J, Bastos C, Oliveira GS, Oliveira JE, Santos JB, Barroso PAV, Sujii E, Fontes E (2006b). Visitantes florais em espécies cultivadas e não cultivadas de algodoeiro (Gossypium spp.), em diferentes regiões do Brasil. Brasília: Embrapa Recursos Genéticos e Biotecnologia. Boletim de pes-quisa e desenvolvimento. 148:40. |
|
|
Roggia S (2010). Caracterização de fatores determinantes dos aumentos populacionais de ácaros tetraniquídeos em soja. Tese, Doutorado em Entomologia, Universidade de São Paulo, Piracicaba. |
|
|
Shannon CE, Weaver W (1949). The mathematical theory of communication. Urbana: University of Illinois Press. |
|
|
Silva GV (2013). Efeito das plantas Bt de soja e milho sobre pragas não-alvo e seus inimigos naturais. Dissertação, Mestrado em Ciências Biológicas, Universidade Federal do Paraná, Curitiba. |
|
|
Spadotto CA, Gomes MAF, Luchini LC, Andrea MM (2004). Monitoramento de riscoambiental de agrotóxicos: princípios e recomendações. Jaguariúna, SP: Embrapa Meio Ambiente. 29 p. |
|
|
Vojtech E, Meissle M, Poppy GM (2005). Effects of Bt maize on the herbivore Spodoptera littoralis (Lepidoptera: Noctuidae) and the parasitoid Cotesia marginiventris (Hymenoptera: Bracondiae). Transgenic Res. 14:133-144. |
|
|
Yu HL, Yun HL, Kong MW (2011). Risk assessment and ecological effects of transgenic Bacillus thuringiensis crops on non-target organisms. J. Integr. Plant Biol. 53:520-538. |
|
|
Whitehouse MEA, Wilson L J, Constable GA (2007). Target and non-target effects on the invertebrate community of Vip cotton, a new insecticidal transgenic. Aust. J. Agric. Res. 58:383-385. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


