ABSTRACT
Indolocarbazole (ICZ) alkaloid constitutes a group of natural products which manifested powerful biological activity, especially anticancer. However, it was laborious to find ICZs-yield organism when anticancer activity was adopted as selection target. The specificity of 6 primer pairs was detected and the results revealed that primer rebBN1/rebBC1 possessed preferable specificity because indolocarbazole-producers DNA was exclusively amplified and no target fragment was amplified from any of indolocarbazole nonproducers. Therefore, primer rebBN1/rebBC1 could specifically identify indolocarbazole-producers from microorganism strains.
Key words: Indolocarbazole, rebBN1/rebBC1, ICZs-producing microorganism screening.
The indolocarbazole (ICZ) family is an important class of natural products isolated from bacteria, actinomycetes, cyanobacteria, fungi, slime moulds and marine inver-tebrates (Sanchez et al., 2006). Since the discovery of staurosporine as the first ICZs in 1977, a variety of ICZs (more than 120) have been found and attracted great attention of chemists, biologists, physicians and pharmaceutical for their biological activities and the prospective number of derivatives from the basic back-bone alone. Indolocarbazole (ICZs) compounds are structurally characterized by possessing an indolo[2,3-a]pyrrolo[3,4-c ]carbazole core derived from two units of tryptophan, with sugars attached derived from glucose and methionine. According to their structural features and mechanisms of actions, ICZs are divided into two major classes (Gribble and Berthel, 2001). The first class was characterized by a sugar moiety linked to both indole nitrogen atoms of the indolocarbozole core to form a“closed” indol-carbazole and showed inhibition to protein kinase C, such as K-252a and staurosporine (STA). The second class contains a sugar moiety attached only to one of the indole nitrogen atoms of the aglycone by a β-glycosidic linage to form an “open” bisindolymaleimide and exhibit inhibition to DNA topoisomerase I, such as rebeccamycin (REB) and AT2433. Despite the differences, it is assumed that the presence of the sugars in both classes appears to be essential for the biological activity of ICZ compounds (Bailly et al., 1999).
ICZs display a wide range of biological activities including antibacterial, antifungal, antiviral, hypotensive, and neuro-protective activities. However, the most attracting point is their therapeutic potential as anti-cancer drugs. Recently, their strong effects to various harmful insects were discovered by our team (related article under review). Although several ICZs derivatives have already entered clinical trials for the treatment of cancer or other diseases, no analog has yet been launched onto the market and great efforts have been made to find or generate ICZs derivatives with improved properties for drugs. Despite the abundant diversity of ICZs-origin organisms, microbes were the main origin of most known ICZs. During the search for novel ICZs-producing organisms and the isolation of ICZs, blindness and repetitiveness have been the formidable obstacles with great loss of labor and time. In order to solve the problem, the feasi-bility of using molecular probes combined with HPLC to detect ICZs-producing microorganisms was investigated in this article.
Strains in this research
Totally, 56 strains with potential pharmaceutical activity, including bacteria 10, actinomycetes 36, and fungi 10 deposited in China Pharmaceutical Marine Microbiological Databank (which was established by our team) were selected as test microbes. Among them, Streptomyces nitrosporus CQT14-24 (T14-24) was the staurosporine-producing strain which was under intensive investi-gation in our lab. This strain was isolated from sediment of Chukchi Sea (the North Pole) and were deposited both in China Pharmaceutical Marine Microbiological Databank (No.HTTMS-F04008) and China General Microbiological Culture Collection Center (No.CGMCC 4607), and recorded at GenBank with the accession number FJ821473. S. nitrosporus CQT5-L25-1 (5-L25-1) was a mutant of T14-24 with increased yield of staurosporine and increased kinds of ICZs homolog. T14-24 and 5-L25-1 were used as positive control.
Culture conditions and DNA isolation
For ICZs extraction, bacteria, actinomycete and fungi strains were shaking cultured by using MYPD medium, Gauze No.1 medium, and MPDA medium at 25°C for 7 days, 12 days and 10 days respectively. For DNA extraction, strains were cultured by using the above mentioned medium for 2 days. Bacteria and actinomycete DNA were isolated by standard cetyl trimethyl ammonium bromide (CTAB) method while fungi DNA was isolated through glass beads method (Alessandro et al., 2001).
Molecular probes adopted in this article
Different conservative sequences of ICZs gene cluster were sought among known literatures and selected primers were listed as Table 1. Primers rebBN1 / rebBC1 were initially designed to amplify rebD of rebeccamycin gene cluster from Streptomyces sp. TP-A0274 (Onaka et al., 2002). Primers CS035 / CS036 were designed to carry out in situ colony hybridization of a constructed cosmid library of Streptomyces longisporoflavus DSM10189 which was highly similar to rebD in Lechevalieria aerocolonigenes (Salas et al, 2005). Primers StaCN / StaCH and StaPN /StaPH were designed to amplify staC and staP in L. aerocolonigenes (Howard-Jones et al., 2006). Primers RebD1 / RebD2 were designed to amplify rebO in L. aerocolonigenes ATCC 39243 (Howard-Jones et al., 2005). Primers NGT1 / NGT2 were designed to amplify ngt (now named rebG) in L. aerocolonigenes ATCC 39243 (Onaka et al., 2003).
Polymerase chain reaction (PCR) amplification
PCR amplifications were performed in 50 μl containing 1 μl template, 1 μl of each primer (10 μmol/L), 1 μl deoxynucleotide triphosphates (2.5 mmol/L), 5 μl 10× polymerase chain reaction (PCR) buffer, 1.5 μL MgCl2 (1.5 mmol/L), 1.5 μL dimethyl sulfoxide (DMSO), 1.5μL bovine serum albumin (BSA) (0.1%), 0.25μL TaqDNA polymerase (2U). The cycling parameters were: initial denaturation for 5 min at 95°C followed by 35 cycles of denaturation at 94°C for 1 min, annealing at 55°C for 1 min, and extension at 72°C for 2 min, and a terminal extension step of 72°C for 15 min.
Isolation of ICZs
After fermentation, the culture broth of different strains was filtered through cheesecloth to separate it into supernatant and mycelia; the former was extracted with equal volume ethyl acetate (EtOAc) for two times while the later was extracted with EtoAc (200ml) by supersonic method; both of the EtOAc solutions were concentrated under reduced pressure to afford an MeOH solutions (5 ml). The concentration of indolecarbazole alkaloids in MeOH solutions was determined by high performance liquid chromatography (HPLC) method. The analytical HPLC system was composed of Waters Acquity UPLC and BEH c18 column (1.7 μm, 2.1 × 5 0mm); chromatographic condition: (0 to 9 min, 5%-100% MeOH, 9-11 min, 100% min, 0.5 ml/min).
ICZs producing microorganisms
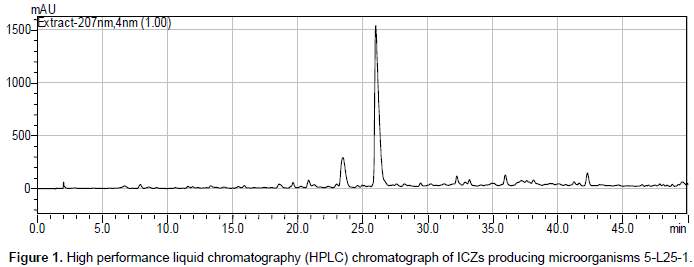
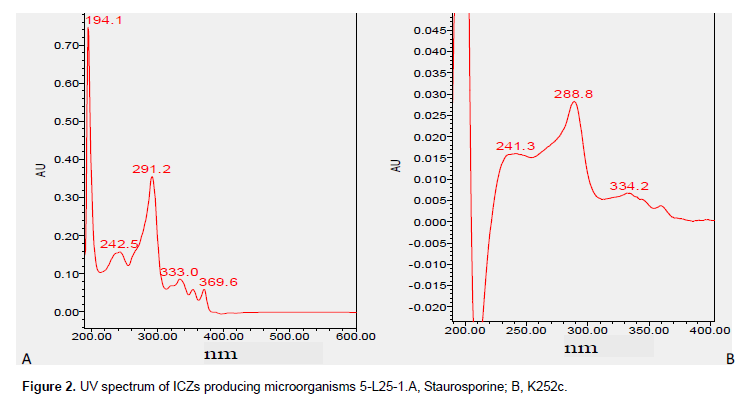
Among 54 test strains, four strains (all actinomycetes) including Streptomyces sp. T9-33, S. nitrosporeus T13-12, Streptomyces sp. YH3-2, and Streptomyces sp. YT1-28 were detected to yield ICZs compounds while other strain showed no production of ICZs. Therefore, there were totally 6 strains (T14-24 and 5-L25-1 as positive control) which produced ICZ compounds. The HPLC spectrum of the six strains was similar with difference ofpeak area. Among them, strain 5-L25-1 yielded the maximum amount of ICZ (peak area 17089081) while strain YH 3-2 yielded the minimum of ICZ (peak area 1311480) (Figure 1). The ultraviolet absorption spectrum of the 5 strain except 5-L25-1 was similar with the main component of staurosporine. As for 5-L25-1, the ultraviolet absorption spectrum showed two main components staurosporine and K252c (Figure 2).
The specificity of different primers
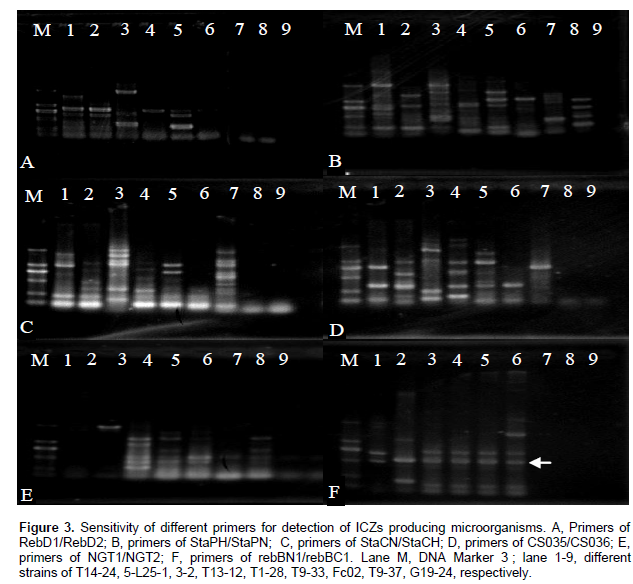
Different open reading frames (ORFs) fragments were amplified using specific primers and the results showed that for almost primers except StaPH/StaPN visible fragments were amplified only when chromosomal DNAs from indolocarbazole producers were used as templates. However, there was no regularity and specific indicator band(s) among the fragments bands except primers rebBN1/rebBC1 (Figure 3, A-E lane 1-6).
The specificity of primers rebBN1/rebBC1
The primers, rebBN1/rebBC1, were synthesized using the highly conserved regions corresponding to amino acids from 611 to 618 and from 780 to 786 in RebD amino acid sequence and were supposed to produce fragment of 0.5 to 0.7 kb. As shown in Figure 2F, a 0.5 kb and/or 0.75 kb fragments were amplified only when chromosomal DNAs from indolocarbazole producers were used as templates (Figure 3F; lane 1-6). On the other hand, any DNA fragment with the size of neither 0.5 kb nor 0.75 kb was amplified with chromosomal DNAs from indolocarbazole nonproducers. The results demonstrate that this primers pairs possessed preferable specificity because indolocarbazole-producers DNA was exclusively amplified. Since no target fragment was amplified from any of indolocarbazole nonproducers, this primer pair could specifically identify indolocarbazole-producers from strains.
ICZs have attracted the attention of many researchers from different disciplines for their biological activities especially anticancer property. This property may be due to different mechanisms of action, including inhibition of protein kinases, DNA intercalation, and inhibition of DNA topoisomerases. Protein kinases and DNA topoiso-merases are important in cell proliferation so specific inhibitors are promising candidates for antitumor or antifungi drugs and pesticide to kill harmful insects. Despite the high inhibition activities, certain known ICZs also exhibited high toxicity with low specificity which limited their direct clinical application. Therefore, great efforts have been made to search or generate novel ICZs derivatives with higher inhibitory specificity. Although organic chemistry methods are routinely used to synthesize and modify natural products, harvesting the product (or a modifiable precursor) from the natural source is often the only cost-effective way of production. Despite the abundant diversity of ICZs-origin organisms, most ICZs are isolated from actinomycetes: rebeccamycin is from L. aerocolonigenes ATCC 39243 (Nettleton et al., 1985), staurosporine is from L. aerocolonigenes subsp. staurosporeus and Streptomyces sp. strain TP-A0274 (Omura et al., 1977), K252a is from Nocardiopsis sp. strain K252 (Kase et al., 1986). Herein, actinomycetes were considered as the main test strains in this research along with certain bacteria and fungi strains and our results also verified this phenomenon. Among 54 test strains, 4 ICZs-yielding strains were all actinomycetes while no bacteria and fungus strains was detected to produce ICZs. Although the searching scope has been narrowed mainly to actinomycetes during the search for natural novel ICZs, it was still laborious to find ICZs-yield organism when anticancer activity was adopted as selection target.
Therefore, in order to reduce the blindness, specific molecular probes might be feasible in effectively narrowing screening strains scope considering the fact that ICZs were biosynthesized through similar pathways and encoded by corresponding biosynthesis gene cluster. Up to now, although the basic biosynthetic pathway for the indolocarbazole aglycone was revealed in the analysis of rebeccamycin and staurosporine biosynthesis, modified pathways for the individual indolocarbazole compounds remain unidentified. Studies on the biosyn-thesis of ICZs have shown that the indolocarbazole core is formed by decarboxylative fusion of two tryptophan-derived units, whereas the sugar moiety is derived from glucose (Pearce et al., 1988; Meksuriyen and Cordell, 1988). Biosynthetic studies through isotope-labeled pre-cursors indicate that the rebeccamycin and staurosporine indolocarbazole cores are derived from L-tryptophan (L-Trp) via a series of oxidative transformations. The initial pathways to the two aglycones follow very similar routes, differing only by the oxidation state at the C-7 position of aglycone and the pattern of connectivity between the deoxysugar and aglycone. Subsequent N-glycosylation and tailoring modifications follow divergent pathways toward rebeccamycin and staurosporine (Howard-Jones et al., 2006). Biosynthesis of ICZs can be divided into five stages catalyzed by different sets of enzymes, which are (i) tryptophan modification (halogenation) by RebH; (ii) dimerization by RebO/RebD or StaO/StaD; (iii) decarboxylative ring closure by RebC/RebP or StaC/StaP; (iv) glycosylation by RebG or StaG; and (v) sugar modification by RebM or several enzymes for staurosporine before and/or after glycosylation (refs. 10–13 and this article). Stages ii and iii constitute the central reactions in the pathway, whereas the other stages can be considered as accessories for ‘‘tailoring’’ the alkaloid skeleton (Sanchez et al., 2005). In this sense, RebO/RebD (or StaO/StaD) and RebC/RebP (or StaC/StaP) play crucial role in the formation of the ICZs molecule backbone. On the other hand, some researchers consider glycosyl in ICZs is essential for its biological activities (Sanchez et al., 2005). Therefore, we focus our intention also on genes encoded these two enzymes.
In recent years, an increasing amount of information has become available on the molecular genetics of indolocarbazole biosynthesis. Until now, four kinds of ICZs biosynthetic gene clusters including rebeccamycin, staurosporine, AT2433, and K252a were cloned. The genetic organization of these four indolocarbazole gene clusters is quite similar and all of them contain genes required for the biosynthesis of the aglycone, biosynthesis and transfer of the deoxysugar and regulation. The rebeccamycin biosynthesis gene cluster was taken as an example. On the basis of sequence analysis and database searches about rebeccamycin gene cluster from L. aerovolonigenes ATCC 39243, Sanchez et al. (2002) proposed that there are 11 ORFs including four for ICZ-core biosynthesis (rebO, rebD, rebC and rebP), two for halogenation (rebH and rebF), glycosylation (rebG, the name was changed from ngt), and sugar methylation (rebM), as well as a regulatory gene (rebR) and two resistance and secretion genes (rebU and rebT). There were approximately counterparts in other three gene clusters. Considering the conservation in related gene clusters, gene fragment of different ORFs were adopted as molecular probe to detect ICZs producing microorganisms among bacteria and fungi strains which was deposited in our lab. To be specific, primers rebBN1/rebBC1 and CS035/CS036 were designated to amplify rebD/StaD gene with about0.3 kb/0.5 kb fragment. In the biosynthesis of indolo-carbazoles, chromopyrrolic acid or 11,11'-dichloro-chromopyrrolic acid is presumed to be an intermediate which might be directly biosynthesized by rebD which encodes a putative chromopyrrolic acid synthase (Onaka et al., 2002; Salas et al., 2005). Primers RebD1/RebD2 were designed to amplify rebO fragment which was predicted to be a flavoprotein encode gene (Howard-Jones et al., 2005). Primers StaPN/StaPH and StaCN/StaCH were designed to amplify staP and staC, respectively (Howard-Jones et al., 2006). RebP and RebC are responsible for the oxidative decarboxylation and ring fusion reactions that create the six-ring indolo-pyrrolocarbazole rebeccamycin aglycone (Sanchez et al., 2005). The RebG (or StaG, once named Ngt) protein converted an indolocarbazole to its N-glucoside indicating that RebG (or StaG, once named ngt ) is responsible for N-glycosylation in rebeccamycin biosynthesis (Onaka et al., 2003). An internal fragment of ngt was usually adopted to clone rebeccamycin biosynthetic genes as a probe. Based on these information, 6 primer pairs were selected to amplify different ORF fragment for detection of ICZs-yield microorganisms. The results reveal that there were varied conservation degree among different ORFs, to be specific, staD (or rebD) possesses the maximum of conservation which may be owe to chromopyrrolic acid synthase which is the key enzyme in the indolocarbazole aglycone synthesis and thus its existence is expected to be a useful indicator of indolo-carbazole-producing strains. However, the conservation degree of different fragments in this ORF varied according to our results. Primer pair CS035/CS036 and rebBN1/rebBC1 were both designed to amplify DNA fragment in StaD (or rebD), with designated region of 1 to 323 and 611 to 1110, respectively. The results reveal that fragment amplified by primer pair rebBN1/rebBC1 showed preferable conservation among ICZs-yielding microorganisms. In addition, since chromopyrrolic acid (or 11,11'-dichlorochromopyrrolic acid) might be the common intermediate of different kinds of ICZs, the primer pair could theoretically detect microorganism producing each kind ICZs.
On the other hand, although glycosylation is very important because biological activity of ICZs requires the presence of the sugar, the primer pair to amplify gene encode for glycosyltransferase RebG (or StaG, once named ngt) showed low specificity to detect ICZs-yielding microorganisms. This might be because of the major structural difference between the two kinds of ICZs: either through a single nitrogen (in the case of rebeccamycin) or through two nitrogen and this property might lead to the flexibility of glycosyltransferase which makes RebG (or StaG) improper probe region.
The author(s) have not declared any conflict of interests.
This study was supported by the National High Technology Research and Development Program (863 Program) of China (2011AA10A202-2), National Key Technologies R&D Program (2011BAE06B04) and the National Natural Science Foundation of China (40976104).
REFERENCES
Bailly C, Qu X, Graves DE, Prudhomme M, Chaires JB (1999). Calories from carbohydrates: energetic contribution of the carbohydrate moiety of rebeccamycin to DNA binding and the effect of its orientation on topoisomerase I inhibition. Chem Biol. 6:277-286.
Crossref |
|
|
|
Gribble GW, Berthel SJ (2001). A survey of indolo[2,3-a]carbazoles and related natural products. In: Rahman AU Studies in natural products chemistry. 12:365-409. |
|
|
Howard-Jones AR, Walsh CT (2005). Enzymatic generation of the chromopyrrolic acid scaffold of rebeccamycin by the tandem action of RebO and RebD. Biochemistry. 44(48):15652-15663.
Crossref |
|
|
Howard-Jones AR, Walsh CT (2006). Staurosporine and rebeccamycin aglycones are assembled by the oxidative action of StaP, StaC, and RebC on chromopyrrolic acid. J Am Chem Soc. 128(37):12289-98.
Crossref |
|
|
Kase H, Iwahashi K, Ymatsuda T (1986). K-252a, a potent inhibitor of protein kinase C from microbial origin. J. Antibiotics. 39:1059-1065.
Crossref |
|
|
|
Meksuriyen D, Cordell GA. Biosynthesis of staurosporine (1988). 2. Incorporation of tryptophan. J. Nat. Prod. 51:893-899. |
|
|
Nettleton DE, Doyle TW, Krishnan B, Matsumoto GK, Clardy J (1985). Isolation and structure of rebeccamycin-a new antitumor antibiotic from Nocardia aerocoligenes. Tetrahedron lett. 26:4011-4014.
Crossref |
|
|
Omura S, Iwai Y, Hirano A, Nakagawa A, Awaya J, Tsuchiya H, Takahashi Y, Masuma R (1977). A new alkaloid AM-2282 of Streptomyces origin taxonomy, fermentation, isolation and preliminary characterization. J. Antibiot. 30(4):275-282. http://en.wikipedia.org/wiki/PubMed_Identifier
Crossref |
|
|
Onaka H, Tanifguchiy SI, Igarashi A, Furumai T (2002). Cloning of the staurosporine biosynthetic gene cluster from streptomyces sp. TP-A0274 and its heterologous expression in Streptomyces lividans. J. Antibiotics. 55(12):1063-1071.
Crossref |
|
|
Onaka H, Taniguchi S, Igarashi Y, Furumai T (2003). Characterization of the biosynthetic gene cluster of rebeccamycin from Lechevalieria aerocolonigenes ATCC 39243. Biosci. Biotechnol. Biochem. 67(1):127-138.
Crossref |
|
|
Pearce CJ, Doyle TW, Forenza S, Lam KS, Schroeder DR (1988). The biosynthetic origins of rebeccamycin. J. Nat. Prod. 51: 937-940.
Crossref |
|
|
Salas AP, Zhu L, Sánchez C, Bra-a AF, Rohr J, Méndez C, Salas JA (2005). Deciphering the late steps in the biosynthesis of the anti-tumour indolocarbazole staurosporine: sugar donor substrate flexibility of the StaG glycosyltransferase. Mol Microbiol. 58(1):17-27.
Crossref |
|
|
Sanchez C, Butovich AI, Brana FA, Rohr J, Mendez C, Salas AJ (2002). The biosynthetic gene cluster for the antitumor rebeccamycin: characterization and generation of indolocarbazole devivatives. Chem Biol. 9:519-531.
Crossref |
|
|
Sanchez C, Mendez C, Salas JA (2006). Indolocarbazole natural products: occurrence, biosynthesis, and biological activity. Nat Prod Rep. 23:1007-1045.
Crossref |
|
|
Sanchez C, Zhu L, Brana F A, Salas AP, Rohr J, Mendez C, Salas JA (2005). Combinatorial biosynthesis of antitumor indolocarbazole compounds. Proc Natl Acad Sci USA. 102:461-466.
Crossref |