Full Length Research Paper

ABSTRACT
INTRODUCTION
MATERIALS AND METHODS
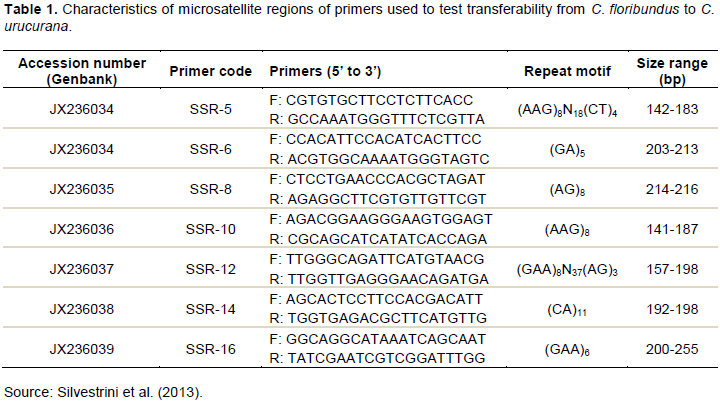
RESULTS AND DISCUSSION
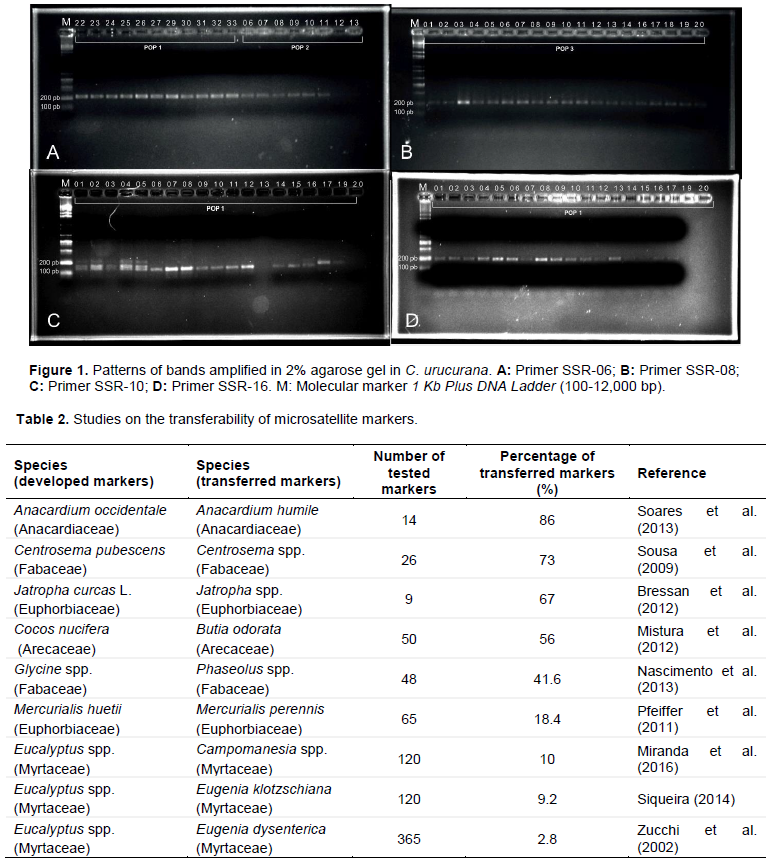
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
REFERENCES
|
Almeida-Pereira CS, Silva AVC, Alves RP, Feitosa-Alcantara RB, Arrigoni-Blank MF, Alvares-Carvalho SV, Costa TS, White LAS, Pinto VS, Sampaio TS, Blank AF (2017). Genetic diversity of native populations of Croton tetradenius Baill. using ISSR markers. Genetics and Molecular Research16(2):gmr16029602. |
|
|
Alves M, Araújo MFL, Gusmão CLS, Lira-Neto AC, Carvalho R, Benko-Iseppon AM (2012). Diversity and Uses of the Genus Croton (Euphorbiaceae) in Northeastern Brazil. In: Rai MK, Cordell GA, Martinez JL, Marinoff M, Rastrelli L (eds) Medicinal Plants: Biodiversity and Drugs, 1st edn. CRC Press, New York, pp. 106-141. |
|
|
Angelo PCS, Chaves FCM, Xavier JJBN, Cruz JC, Lira MPS (2005). Avaliação inicial do RAPD como forma de acessar a variabilidade genética na espécie rica em linalol Croton cajuçara Benth. (sacaca). Manaus: Embrapa Amazônia Ocidental 28:1-4. |
|
|
Barbara T, Palma-Silva C, Paggi GM, Bered F, Fay MF, Lexer C (2007). Cross- species transfer of nuclear microsatellite markers: potential and limitations. Molecular Ecology 16:3759-3767. |
|
|
Bressan EDA, Scotton DC, Ferreira RR (2012). Development of microsatellite primers for Jatropha curcas (Euphorbiaceae) and transferability to congeners. American Journal of Botany 99:237-239. |
|
|
Burger WC, Huft M (1995). Flora Costaricensis: Family #113 Euphorbiaceae. Fieldiana Botany 36:1-169. |
|
|
Caruzo MBR, Cordeiro I (2007). Synopsis of the tribe Crotoneae Dumort. (Euphorbiaceae s.s.) in the State of São Paulo, Brazil. Hoehnea 34(4):571-585. |
|
|
Coley PD (1993). Gap size and plant defenses. Trends in Ecology & Evolution 8(1):1-2. |
|
|
Deshmukh VP, Thakare PV, Chaudhari US, Gawande PA (2007). A simple method for isolation of genomic DNA from fresh and dry leaves of Terminalia arjuna (Roxb.) Wight and Arnot. Electronic Journal of Biotechnology 10:468-472. |
|
|
Doyle JJ, Doyle JL (1990). Isolation of plant DNA from fresh tissue. Focus 12:13-15. |
|
|
Ferreira ME, Grattapaglia D (1998). Introducción al uso de marcadores moleculares en el análisis genético. Brasília: Embrapa-Cenargen. |
|
|
Guichoux E, Lagache L, Wagner S, Chaumeil P, Léger P, Lepais O, Lepoittevin C, Malausa T, Revardel E, Salin F, Petit RJ (2011). Current trends in microsatellite genotyping. Molecular Ecology Resources 11:591-611. |
|
|
Hodel RGJ, Segovia-Salcedo MC, Landis JB, Crowl AA, Sun M, Liu X, Gitzendanner MA, Douglas NA, Germain-Aubrey CC, Chen S, Soltis DE, Soltis PS (2016). The report of my death was an exaggeration: A review for researchers using microsatellites in the 21st century. Applications in Plant Sciences 4(6):apps.1600025. |
|
|
Kalia RJ, Rai MK, Kalia S, Singh R, Dhawan AK (2011). Microsatellite markers: an overview of the recent progress in plants. Euphytica 177(3):309-334. |
|
|
Lorenzi H (2002). Árvores brasileiras: manual de identificação e cultivo de plantas arbóreas nativas do Brasil. 2nd edn. Nova Odessa: Instituto Plantarum. |
|
|
Maciel MAM, Pinto AC, Veiga Jr. VF (2002). Plantas Medicinais: a necessidade de estudos multidisciplinares. Quimica Nova 25(3):429-438. |
|
|
Miranda EAGC, Boaventura-Novaes CRD, Braga RS, Reis EF, Pinto JFN, Telles MPC (2016). Validation of EST-derived microsatellite markers for two Cerrado-endemic Campomanesia (Myrtaceae) species. Genetics and Molecular Research 15(1):gmr.15017658. |
|
|
Mishra RK, Gangadhar BH, Yu JW, Kim DH, Park SW (2011). Development and characterization of EST based SSR markers in Madagascar periwinkle (Catharanthus roseus) and their transferability in other medicinal plants. Plant Omics 4(3):154-161. |
|
|
Mistura CC, Barbieri RL, Castro CM, Priori D, Villela JCB (2012). Transferibilidade de marcadores microssatélites de coco (Cocos nucifera) para butiá (Butia odorata). Magistra 24(4):360-369. |
|
|
Mnejja M, Garcia-Mas J, Audergon JM, Arus P (2010). Prunus microsatellite marker transferability across rosaceous crops. Tree Genetics & Genomes 6(5):689-700. |
|
|
Nascimento ATB, Silva CB, Costa MMR, Silva RLO, Kido EA (2013). Transferibilidade de marcadores moleculares de soja para aplicação no melhoramento de feijão caupi. In: XII Jornada de Ensino, Pesquisa e Extensão - JEPEX, 2013. Anais. Recife/PE: UFPE. |
|
|
Oliveira TG, Pereira AMS, Coppede JS, França SC, Ming LC, Bertoni BW (2016). Genetic diversity analysis of Croton antisyphiliticus Mart. using AFLP molecular markers. Genetics and Molecular Research 15(1):gmr.15017461. |
|
|
Peakall R, Gilmore S, Keys W, Morgante M, Rafalski A (1998). Cross-species amplification of soybean (Glycine max) simple sequence repeats (SSRs) within the genus and other legume genera: implications for the transferability of SSRs in plants. Molecular Biology and Evolution 15(10):1275-1287. |
|
|
Pfeiffer T, Roschanski AM, Pannell JR, Korbecka GY, Schnittler M (2011). Characterization of microsatellite loci and reliable genotyping in a polyploid plant, Mercurialis perennis (Euphorbiaceae). Journal of Heredity 102(4):479-488. |
|
|
Randau KP, Florêncio DC, Fereira CP, Xavier HS (2004). Pharmacognostic study of Croton rhamnifolius H.B.K. and Croton rhamnifolioides Pax & Hoffm. (Euphorbiaceae). Revista Brasileira de Farmacognosia 14(2):89-96. |
|
|
Rath P, Rajaseger G, Goh CJ, Kumar PP (1998). Phylogenetic analysis of Dipterocarps using random amplified polymorphic DNA markers. Annals of Botany 82:61-65. |
|
|
Resende MDV (1999). Melhoramento de essências florestais. In: Borém A. Melhoramento de espécies cultivadas. UFV: Viçosa, MG, Brazil, pp. 589-647. |
|
|
Riina R, Berry PE, Van Ee BW (2009). Molecular phylogenetics of the dragon's blood Croton section Cyclostigma (Euphorbiaceae): a polyphyletic assemblage unraveled. Systematic Botany 34(2):360-374. |
|
|
Rinthong PO, Zhu S, Komatsu K, Chanama S, De-Eknamkul W (2011). Genetic variation of Croton stellatopilosus Ohba based on non-coding DNA sequences of ITS, trnK and trnL-F regions. Journal of Natural Medicines 65(3-4):641-645. |
|
|
Romano E, Brasileiro ACM (1999). Extração de DNA de plantas: Soluções para problemas comumente encontrados. Biotecnologia Ciência e Desenvolvimento: Brasília 2(9):40-43. |
|
|
Salatino A, Salatino MLF, Negri G (2007). Traditional uses, chemistry and pharmacology of Croton species (Euphorbiaceae). Journal of the Brazilian Chemical Society 18(1):11-33. |
|
|
Scaldaferri MM, Freitas JS, Santos ESL, Vieira JGP, Gonçalves ZS, Cerqueira-Silva CBM (2013). Comparison of protocols for genomic DNA extraction from velame pimenta (Croton linearifolius), a native species to the Caatinga, Brazil. African Journal of Biotechnology 12(30):4761-4766. |
|
|
Scaldaferri MM, Freitas JS, Vieira JGP, Gonçalves ZS, Souza AM, Cerqueira-Silva CBM (2014). Comparison of methods for estimates of molecular genetic diversity in genus Croton: influence of coefficients, clustering strategies and data projection. Genetics and Molecular Research 13(3):5566-5573. |
|
|
Silva TSS, Freitas JS, Santos ESL, Cardoso TS, Cerqueira-Silva CBM (2018). Caracterização e seleção de marcadores moleculares em Croton linearifolius Mull. Arg. como subsídio para estudos genéticos. Multi-Science Journal 1(10):4-8. |
|
|
Silvestrini M, Pinto-Maglio CAF, Zucchi MI, Santos FAM (2013). Cytogenetics and characterization of microsatellite loci for a South American pioneer tree species, Croton floribundus. Genome 56(12):743-751. |
|
|
Silvestrini M, McCauley DE, Zucchi MI, Santos FAM (2015). How do gap dynamics and colonization of a human disturbed area affect genetic diversity and structure of a pioneer tropical tree species? Forest Ecology and Management 344:38-52. |
|
|
Siqueira MN (2014). Transferibilidade e variabilidade genética de marcadores microssatélites gênicos em Eugenia klotzschiana Berg. (Myrtaceae). MSc Thesis, Federal University of Goiás, UFG. |
|
|
Soares TN, Sant'Ana LL, Oliveira LK, Telles MPC, Collevatti RG (2013). Transferability and characterization of microsatellite loci in Anacardium humile A. St. Hil. (Anacardiaceae). Genetics and Molecular Research 12(3):3146-3149. |
|
|
Sousa ACB, Carvalho MA, Boaventura LR (2009). Microsatellite markers in tropical legume (Centrosema pubescens Benth): development, characterization, and cross-species amplification in Centrosema sp. Conservation Genetics Resources 1:347-352. h |
|
|
Van Ee BW, Riina R, Berry PE (2011). A revised infrageneric classification and molecular phylogeny of New World Croton (Euphorbiaceae). Taxon 60(3):791-823. |
|
|
Varshney RK, Graner A, Sorrells ME (2005). Genic microsatellite markers in plants: features and applications. Trends in Biotechnology 23(1):48-55. |
|
|
Wang ML, Barkley NA, Jenkins TM (2009). Microsatellite markers in plants and insects. Part I: Applications of biotechnology. Genes, Genomes and Genomics 3(1):54-67. |
|
|
Zucchi MI, Brondani RPV, Pinheiro JB, Brondani C, Vencovsky R (2002). Transferability of microsatellite markers from Eucalyptus spp. to Eugenia dysenterica (Myrtaceae family). Molecular Ecology Notes 2:512-513. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


