Full Length Research Paper

ABSTRACT
INTRODUCTION
MATERIALS AND METHODS
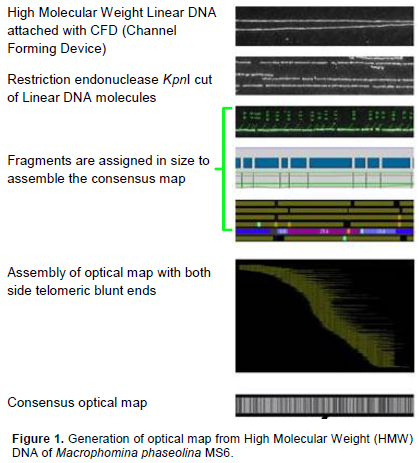
RESULTS
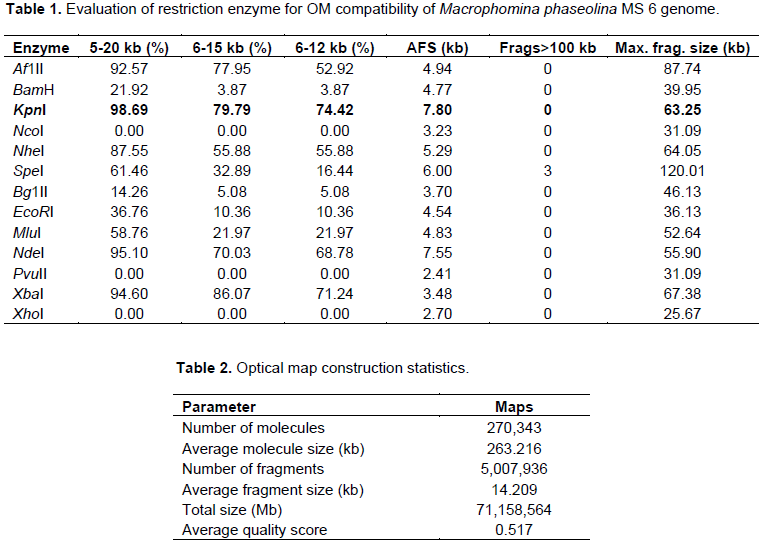
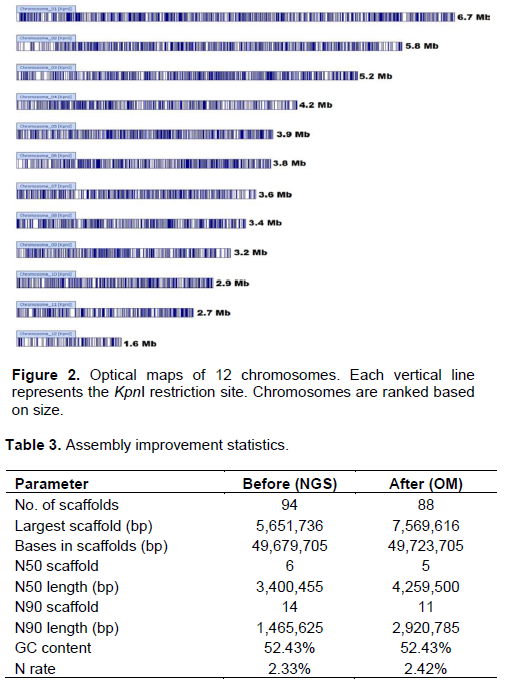
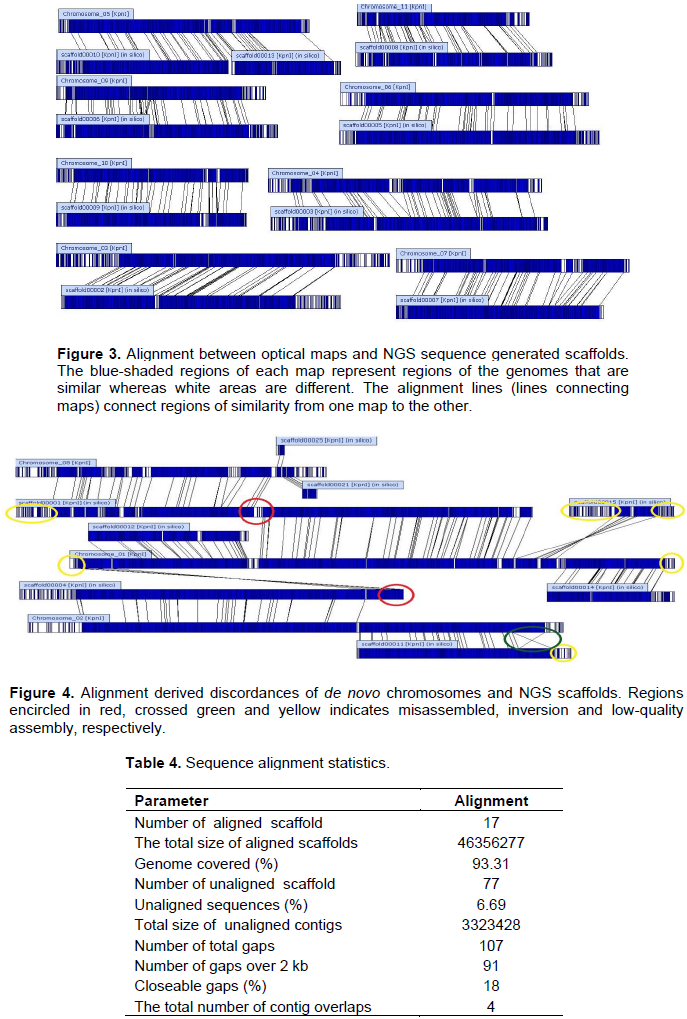
DISCUSSION
CONCLUSION
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
REFERENCES
|
Abbas MF, Farah N, Gulshan I (2013). Important fungal diseases of potato and their management - a brief review. Mycopathology 11(1):45-50. |
|
|
Aly AA, Abdel SMA, Omar MR, Abd-Elsalam KA (2007). Differential antagonism of Trichoderma sp. against Macrophomina phaseolina. Journal of Plant Protection Research 47(2):91-102. |
|
|
Aston C, Mishra B, Schwartz DC (1999). Optical mapping and its potential for large-scale sequencing projects. Trends in Biotechnology 17(7):297-302. |
|
|
Bangladesh Bureau of Statistics (2011). Government of the People's Republic of Bangladesh, Bangladesh Bureau of Statistics, Agriculture Wing, Parishankhyan Bhaban E-27/A, Agargaon, Dhaka-1207. |
|
|
Biemond C, Oluwole O, Tjeerd JS, Kumar L, Aad T, Struik PC (2013). Health of farmer-saved maize seed in north-east Nigeria. European Journal of Plant Pathology 137(3):563-572. |
|
|
Chamala S, Chanderbali AS, Der JP, Lan T, Walts B, Albert VA, dePamphilis CW, Leebens-Mack J, Rounsley S, Schuster SC, Wing RA, Xiao N, Moore R, Soltis PS, Soltis DE, Barbazuk WB (2013). Assembly and validation of the genome of the non-model basal angiosperm Amborella. Science 342(6165):1516-1517. |
|
|
Chen S, Xu J, Liu C, Zhu Y, Nelson DR, Zhou S, Li C, Wang L, Guo XZ, Sun Y, Luo H, Li YR, Song J, Henrissat B, Levasseur A, Qian J, Li J, Luo X, Shi L, He L, Li X, Xu X, Niu Y, Li Q, Han MV, Yan H, Zhang J, Chen H, Lv A, Wang Z, Liu M, Schwartz DC, Sun C (2012). Genome sequence of the model medicinal mushroom Ganoderma lucidum. Nature Communications 3(1):913. |
|
|
Da Silva WL, Clark CA (2013). Infection of Sweet potato by Fusarium solani and Macrophomina phaseolina Prior to Harvest. Plant Disease 97(12):1636-1644. |
|
|
Dinakaran D, Mohammed N (2001). Identification of resistant sources to root rot of sesame caused by M. phaseolina (Tassi.) Goid. Sesame and Safflower Newsletter 16:68-71. |
|
|
De BK, Chattopadhya SB, Arjunan G (1992). Effect of potash on stem rot diseases of jute caused by Macrophomina phaseolina. Journal of Mycopathological Research 30(1):51-55. |
|
|
Dong Y, Xie M, Jiang Y, Xiao N, Du X, Zhang W, Gwenola TK, Jinhuan W, Shuang Y, Jie L, Wenbin C, Jing C, Peng Z, Yong H, Chao B, Shengkai P, Yuxiang L, Xin L, Wenliang W, Bertrand S, Brian S, Bin Z, Deacon S, Rich M, Wenhui N, Yongyi S, Ruoping Z, Guojie Z, Jinquan L, Thomas F, James W, Yaping Z, James K, Noelle C, Xun X, Shuhong Z, Jun W, Wen W, Wang W (2013). Sequencing and automated whole-genome optical mapping of the genome of a domestic goat (Capra hircus). Nature Biotechnology 31(2):135-141. |
|
|
Ganapathy G, Howard JT, Ward JM, Li J, Li B, Li Y, Xiong Y, Zhang Y, Zhou S, Schwartz DC, Schatz M, Aboukhalil R, Fedrigo O, Bukovnik L, Wang T, Wray G, Rasolonjatovo I, Winer R, Knight JR, Koren S, Warren WC, Zhang G, Phillippy Jarvis AM (2014). Highâ€coverage sequencing and annotated assemblies of the budgerigar genome. GigaScience 3:11 doi: 10.1186/2047-217X-3-11. |
|
|
Ghurye J, Pop M (2019). Modern technologies and algorithms for scaffolding assembled genomes. PLOS Computational Biology 15(6):e1006994. |
|
|
Hyder S, Gondal AS, Ahmed R, Sahi ST, Rehman A, Hannan A (2018). First Report of Charcoal Rot in Tomato Caused by Macrophomina phaseolina (Tassi) Goid From Pakistan. Plant Disease 102(7):1459. |
|
|
Hai KMA, Ali AAE, El-Metwally MA (2017). Down-regulation of Damping-off and Root Rot Diseases in Lentil Using Kinetin and Trichoderma. International Journal of Agricultural Research 12(1):41-51. |
|
|
Islam MS, Haque S, Islam MM, Emdad EM, Halim A, Hossen QM, Hossain MZ, Ahmed B, Rahim S, Rahman MS, Alam, MM, Hou S, Wan X, Saito JA, Alam M (2012). Tools to kill: Genome of one of the most destructive plant pathogenic fungi Macrophomina phaseolina. BMC Genomics 13(1):493. |
|
|
Kaur S, Dhillon GS, Brar SK, Vallad GE, Chand R, Chauhan VB (2012). Emerging phytopathogen Macrophomina phaseolina: biology, economic importance, and current diagnostic trends. Critical Reviews in Microbiology 38(2):136-151. |
|
|
Khan AN, Faluk S, Kamran M, Zafar H, Ayub KM, Yusuf HF, Nadeem HM (2017). Molecular Identification and Genetic Characterization of Macrophomina phaseolina Strains Causing Pathogenicity on Sunflower and Chickpea. Frontiers in Microbiology 8:1309 |
|
|
Khan SN (2007). M. phaseolina as causal agent for charcoal rot of sunflower. Mycopathologia 5(2):111-118. |
|
|
Koboldt DC, Steinberg KM, Larson DE, Wilson RK, Mardis, ER (2013). The next-generation sequencing revolution and its impact on genomics. Cell 155(1):27-38. |
|
|
Kremer FS, McBride A, Pinto LS (2017). Approaches for in silico finishing of microbial genome sequences. Genetics and Molecular Biology 40(3):553-576. |
|
|
Lai Z, Jing J, Aston C, Clarke V, Apodaca J, Dimalanta ET, Carucci DJ, Gardner MJ, Mishra B, Anantharaman TS, Paxia S, Hoffman SL, Craig Venter J, Huff EJ, Schwartz DC (1999). A Shotgun Optical Map of the Entire Plasmodium Falciparum Genome. Nature Genetics 23(3):309-313. |
|
|
Lim A, Dimalanta ET, Potamousis KD, Yen G, Apodoca J, Tao C, Schwartz DC (2001). Shotgun optical maps of the whole Escherichia coli O157:H7 genome. Genome Research 11(9):1584-1593. |
|
|
Lin J, Lin J, Qi R, Aston C, Jing J, Anantharaman TS, Mishra B, White O, Daly MJ, Minton KW, Venter JC, Schwartz DC (1999). Whole-genome shotgun optical mapping of Deinococcus radiodurans. Science 285(5433):1558-62. |
|
|
Lodha S, Mawar R (2019). Population dynamics of Macrophomina phaseolina in relation to disease management: A review. Journal of Phytopathology 00:1-17. |
|
|
Long E, Evans C, Chaston J, Udall JA (2018). Genomic Structural Variations within Five Continental Populations of Drosophila melanogaster. G3-Genes Genomes Genetics 8(10):3247-3253. |
|
|
Lu C, Song B, Zhang H, Wang Y, Zheng X (2015). Rapid Diagnosis of Soybean Seedling Blight Caused by Rhizoctonia solani and Soybean Charcoal Rot Caused by Macrophomina phaseolina Using LAMP Assays. Phytopathology105 (12):1612-7. |
|
|
Mak AC, Lai YY, Lam ET, Kwok TP, Leung AK, Poon A, Kwok PY(2016). Genome-Wide Structural Variation Detection by Genome Mapping on Nanochannel Arrays. Genetics 202(1):351-362. |
|
|
Majumder S, Datta K, Sarkar C, Saha SC, Datta SK (2018). The Development of Macrophomina phaseolina (Fungus) Resistant and Glufosinate (Herbicide) Tolerant Transgenic Jute. Frontiers in Plant Science 9:920. |
|
|
Mahadevakumar S, Janardhana GR (2016). First report of leaf blight caused by Macrophomina phaseolina on Jasminium multiflorum in india. Journal of Phytopathology 98(1):120-125. |
|
|
Mamun MA, Shamsi S, Bashar MA (2016). Estimation of interrelationships among some quality factors of jute seeds. Dhaka University Journal of Biological Science 25(10):9-17. |
|
|
Mayek-Pérez N, López-Castañeda C, López-Salinas E, Cumpián-Gutiérrez J, Acosta-Gallegos JA (2001): Macrophomina phaseolina resistance in common bean under field conditions in Mexico. Agrociencia 35:649-661. |
|
|
McCain AH, Scharpf RF (1989). Effect of inoculum density of Macrophomina phaseolina on seedling susceptibility of six conifer species. European Journal of Forest Pathology 19(2):119-123. |
|
|
Meena PN, Rajib D, Roy A, Bhimashankar G, Sabyasachi M (2015). Evaluation of stem rot disease in jute (Corchorus olitorius) germplasm caused by Macrophomina phaseolina (Tassi) Goid. Journal of Applied and Natural Science 7(2):857-859. |
|
|
Mukherjee K, Washimkar D, Muggli MD, Salmela L, Boucher C (2018). Error correcting optical mapping data. Gigascience 7(3):61. |
|
|
Mengistu A, Smith JR, Ray JD, Bellaloui N (2011). Seasonal progress of charcoal rot and its impact on soybean productivity. Plant Disease 95(9):1159-1166. |
|
|
OpGen Inc. U.S. 708 Quince Orchard Road Gaithersburg, Maryland 20878 USA. |
|
|
Pendleton M, Sebra R, Pang AW, Ummat A, Franzen O, Rausch T, Stütz AM (2015). Assembly and diploid architecture of an individual human genome via singleâ€molecule technologies. Nature Methods12(8):780-786. |
|
|
Piperkova N, Zarkova M, Ahmed B (2016). Characterization of Macrophomina Phaseolina and Fusarium Spp. isolates from sunflower. Agricultural Sciences VIII (19):95-100. |
|
|
Reslewic S, Zhou S, Place M, Zhang Y, Briska A, Goldstein S, Churas C, Runnheim R, Forrest D, Lim A, Lapidus A, Han CF, Roberts GP, Schwartz DC (2005). Whole-Genome Shotgun Optical Mapping of Rhodospirillum rubrum. Applied and Environmental Microbiology 71(9):5511-5522. |
|
|
Ruperao P, Chan CK, Azam S, Karafiátová M, Hayashi S, Cížková J, Saxena RK, Simková H, Song C, Vrána J, Chitikineni A, Visendi P, Gaur PM, Millán T, Singh KB, Taran B, Wang J, Batley J, Doležel J, Varshney RK, Edwards D (2014). A chromosomal genomics approach to assess and validate the desi and Kabuli draft chickpea genome assemblies. Plant Biotechnology Journal 12(6):778-786. |
|
|
Samad A, Huff EF, Cai W, Schwartz DC (1995). Optical mapping: a novel, single-molecule approach to genomic analysis. Genome Research 5(1):1-4. |
|
|
Schwartz DC, Li X, Hernandez I, Ramnarian SP, Huff EJ, Wang YK (1993). Ordered restriction maps of Saccharomyces cerevisiae chromosomes constructed by optical mapping. Science 262(5130):110-114. |
|
|
Singh SK, Nene YL, Reddy MV (1990). Influence of cropping systems on Macrophomina phaseolina populations in soil. Plant Disease 74(10):812-814. |
|
|
Shukla SK, Kislow J, Briska A, Henkhaus J, Dykes C (2009). Optical Mapping Reveals a Large Genetic Inversion between Two Methicillin-Resistant Staphylococcus aureus Strains. Journal of Bacteriology 191(18):5717-5723. |
|
|
Su G, Suh SO, Schneider RW, Russin JS (2001). Host specialization in the charcoal rot fungus, Macrophomina phaseolina. Phytopathology 91(2):120-126. |
|
|
Teague B, Waterman MS, Goldstein S, Potamousis K, Zhou S, Reslewic S, Sarkar D, Valouev A, Churas C, Kidd JM, Kohn S, Runnhei R, Lamers C, Forrest D, Newton MA, Eichler EE, Kent-First M, Surti U, Livny M, Schwartz DC (2010). High-resolution human genome structure by single-molecule analysis. Proceedings of the National Academy of Sciences 107(24):10848-10853. |
|
|
Udall JA, Dawe RK (2018). Is It Ordered Correctly? Validating Genome Assemblies by Optical Mapping. The Plant Cell 30(1):7-14. |
|
|
Wyllie TD (1998). Soybean Diseases of the North Central Region. In: Wyllie TD, Scott DH, editors. Charcoal rot of soybean-current status: American Phytopathological Society, St. Paul, pp. 106-113. |
|
|
Wyllie TD (1993). Charcoal rot. Compendium of Soybean Diseases. Edited by: Sinclair JB, Backman PA., American Phytopathological Society, St. Paul, pp. 30-33. |
|
|
Zhou S, Bechner M, Place, M, Churas C, Pape L, Leong SA, Runnheim R, Forrest DK, Goldstein S, Livny M, Schwartz DC (2007). Validation of rice genome sequence by optical mapping. BMC Genomics 15(8):278. |
|
|
Zhou S, F. Wei J, Nguyen M, Bechner K, Potamousis S, Goldstein L, Pape MR, Mehan C, Churas S, Pasternak DK, Forrest R, Wise D, Ware RA, Wing MS, Waterman ML, Schwartz DC (2009). A single molecule scaffold for the maize genome. PloS Genetics 5(11):e1000711. |
|
|
Živanov D, TanÄić Živanov S, Nagl N, Savić A, Katanski S, Milić D (2019). First Report of Macrophomina phaseolina on chickpea (Cicer arietinum L.) in Serbia. Plant Disease 103(10):2685. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


