Full Length Research Paper

ABSTRACT
Chickpea (Cicer arietinum L.) is an important leguminous crop grown in different parts of Ethiopia. It is a nutritionally valued and N2-fixing legume which forms a symbiotic association with Mesorhizobium. This study was conducted to characterize and evaluate symbiotic efficacy of chickpea rhizobia isolated from soil samples collected from the study area. Seventeen chickpea rhizobia were isolated by soil host plant trap method and characterized for edaphic stress tolerance. Five of the isolates were grown at high temperature of 45°C and salt concentration of 6%, whereas seven were grown at acidic pH of 4, and four were grown at basic pH of 9. All isolates were not resistant to kanamycin, while fair resistance to erythromycin and streptomycin and modest resistance to ampicillin and azithromycin were observed. Furthermore, most of the isolates showed a variation in nodulation with higher (22 NN/p) and lowest (9 NN/p) scores. Shoot dry weight (SDW) of the plant ranged from 1.18 to 1.84 g/p and isolates showed effective (67%) to highly effective (100%) N2-fixing performance. From these, four isolates showed multiple edaphic stress resistance and are recognized as promising candidate for chickpea production in stressed soil; however, further study in the filed is required.
Key words: Chickpea, Rhizobium, edaphic stress, symbiotic effectiveness
INTRODUCTION
Chickpea (Cicer arietinum L.) is a cool season leguminous crop commonly grown in tropical, subtropical, temperate and semi-arid regions of the world (Miller et al., 2002; Singh et al., 2014). Ethiopia is considered as the center of secondary diversity for chickpea (Van der Maesen, 1987). Chickpea production ranks third among pulse crops grown in the country next to Faba bean (Vicia faba) and Field pea (Pisum sativum). Spatially, Amhara regional state takes the first share and is considered as a potential chickpea producer with 62% of annual production (IFPRI, 2010). Among the existing chickpea varieties, Desi type that was preferably grown in semi-arid tropics is the most dominant in Ethiopia (Naser et al., 2008). Chickpea cultivation in the country covers more than 208,388.6 ha of the land (CSA, 2011).
Chickpea is being valued for its high dietary nutrition and serves as an invaluable source of protein and other nutrients for consumers (Shiferaw and Teklewolde, 2007; Mohammed et al., 2011). Chickpea contains 29% protein, 59% carbohydrate, 3% fiber, 5% oil and 4% ash, and it is a good source of absorbable ions like Ca, P, Mg, Fe and K (Christodoulou et al., 2005). Therefore, it is commonly incorporated as part of the different Ethiopian dishes and used for balanced diet.
On the other hand, chickpea serves as cash generating crop in the country with 312,000 tons of annual production and has appreciated export markets (IFPRI, 2010). For instance, in Ethiopia from the 48% of the pulse exported volume, chickpea accounts for about 27% of the total quantity production, while the remaining is used for domestic market and household consumption (Shiferaw and Teklewolde, 2007).
Besides its nutritional quality and source of income, chickpea plays tremendous role in soil fertility by improvement of symbiotic N2-fixation in association with Mesorhizobia bacteria (Werner, 2005; Funga et al., 2016). Improved soil fertility boosts crop production and maximizes chickpea yield (Jida and Assefa, 2012). The remaining plant biomass in the soil also increases nitrogen pool and serve as a nitrogen source for succeeding crops production by crop rotation cultivation process (Keneni et al., 2011; Beyene et al., 2013). Therefore, symbiotic N2-fixation of chickpea is economically cost effective and environmentally friendly alternative to benefit farmers and help in sustainable crop production by shift cultivation of crops with limited use of synthetic fertilizer (Tena et al., 2017).
The N2-fixation efficiency of chickpea infected by Mesorhizobium strains was determined by soil edaphic factors (Imran et al., 2015). Thus, the chickpea Mesorhizobium which were isolated from the local agro-ecology were expected to infect respective host plants and fix atmospheric nitrogen in a better way (Simon et al., 2014). This is because, indigenous Mesorhizobia are expected to have better adaptation mechanism of the localized soil ecological factors of a given farmland (Beyene et al., 2013). Hence, identifying efficient and superior N2-fixing Mesohizobium strain from the local agro-ecology has paramount importance to enhance chickpea production and improve soil fertility.
Therefore, this study aimed to obtain efficient N2-fixing Mesorhizobial isolates from chickpea rhizosphere soils and identify potential isolates which could be substitutes of synthetic fertilizer for chickpea cultivation.
MATERIALS AND METHODS
Study site and sample collection
This study was conducted in two purposely selected districts located in East Gojjam Zone, Amahara Regional state, Ethiopia because of their dominant production of chickpea (Dejen and Aneded). From those districts, the most potential chickpea grower kebeles (smaller administrative next to district) were identified during the field survey. In each selected kebele, one chickpea farm was taken as soil sample source. From these chickpea farms, triplicated soil samples were pooled by digging at 20 to 30 cm depth. Composite soil samples were collected using ethanol sterilized (70%) plastic bags in November 2015. Collected composite samples were then taken to Microbiology Laboratory, Department of Biology, Debre Markos University (DMU) for further work.
Nodule collection
Nodules were collected from chickpea by using soil-plant trap method in pots under greenhouse condition (Vincent, 1970). The chickpea plants grown in the collected soil samples for 45 days were uprooted and large sized, yellow colored nodules were picked and surface-sterilized (Somesagaran and Hoben, 1994).
Rhizobium isolation
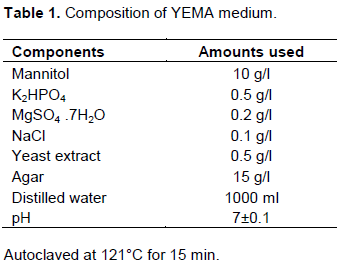
Entrapped Mesorhizobia were isolated from the collected nodules after brief surface sterilization using 70% ethanol for 10 s and 5% local bleach for 3 min (Vincent, 1970). Then, treated nodules were rinsed five times by using sterilized water. Sterilized nodules were then crushed and loop full of sap was transferred onto yeast extract mannitol agar (YEMA) containing plates (Table 1). Inoculated plates were incubated in a bacteriological incubator adjusted at a temperature of 28°C for 3-5 days (Vincent, 1970). After growth, a single colony was picked up and purified periodically by re-streaking method on the fresh YEMA medium. Then, pure isolates were preserved on YEMA slants containing 0.3% (w/v) CaCO3 and stored in refrigerator adjusted at a temperature of 4°C (Vincent, 1970).
Authentication of the isolates
Isolates infectivity of Desi types of chickpea was confirmed by inoculating them onto plant seedlings. Activated isolates were inoculated onto chickpea seedlings planted on sand filed plastic pots and allowed to grow for 45 days in the greenhouse. After 45 days of the growth, plant were uprooted and the existence of nodule were checked.
Characterization of the Isolates
All isolates were checked on YEMA medium containing 25 µg ml-1 Congo red to evaluate their ability to absorb the dye. In addition, isolates were inoculated on medium containing 25 µg ml-1 bromothymol blue (BTB) to determine their ability to produce acid or base and color change of medium were observed (Lupwayi and Haque, 1994). Furthermore, appearance, color and size of the grown colonies were examined on YEMA plates.
Physiological characterization of isolates
All tests were carried out three times on YEMA plates and compared with control. The isolates growth was qualitativlly determined and recorded as (+) for growth, (±) for limited growth and (-) for no growth.
Salt tolerance
Isolates were tested for their salinity tolerance using YEMA medium supplemented with NaCl at concentrations of 0.1, 0.3, 0.5, 0.8, 1, 2, 3, 4, 5 and 6% (w/v) (Belay and Assfa, 2011).
Temperature tolerance
The ability of isolates growth at high and low temperatures were monitored using YEMA medium incubated at 5, 10, 15, 35, 40 and 45°C (Jida and Assefa, 2012).
pH tolerance
Isolates acid and alkaline tolerance were evaluated by growing them on the medium where pH was adjusted to 4, 4.5, 5, 5.5, 8 and 9 using sterile HCl and NaOH (Belay and Assefa, 2011).
Carbohydrates utilization by the isolates
Carbohydrate utilization by isolates was determined using the methods described by Somasegaran and Hoben (1994) on six carbohydrates. These carbohydrates were prepared as 10% (w/v) solution in water. Carbohydrate free medium, which is essentially similar to YEMA medium were modified by reducing yeast extract to 0.05 g/L. Heat-labile carbohydrate solutions were sterilized by membrane filtration method using Millipore with a pore size of 0.22 µm and added to the autoclaved basal medium. The heat-stable carbohydrates were autoclaved together with the medium. YEMA medium without carbon source and with mannitol was used as negative and positive controls, respectively.
Intrinsic antibiotic resistance (IAR)
The intrinsic antibiotic resistance of isolates was determined using some selected antibiotics. The tested antibiotics were ampicillin, streptomycin, kanamycin, erythromycin, azithromycin and chloramphenicol. These antibiotics were incorporated into YEMA medium after membrane filter sterilization using 0.22 µm size at the concentration of 2.5, 5 and 10 µg/ml (Beynon and Josey, 1980). Then, the isolates growth and failure were recorded.
Evaluation of isolates N2-fixation effectiveness
The effectiveness of isolates was tested in a pot experiment conducted in greenhouse condition. 3 kg of carefully washed, sieved and HCl acid sterilized river sand were filled with alcohol-sterilized (70%) plastic pots. Chickpea seeds of uniform size and color were surface sterilized as described before and transferred to 0.75% (w/v) of water agar plates and allowed to germinate at 25°C for 3 days. Four chickpea seedlings were transferred into each pot, which were later thinned down to three. Each isolates grown in YEMA broth medium to logarithmic phase were adjusted to 109 cells ml-1. Activated 1 ml of isolates were inoculated onto each seedling (1 ml/seedling) of the sand culture. The experiment set up was a complete randomized design (CRD) with three replicates. A plus –N with no inoculation and a non-inoculated with no N were used as the controls. The plus control contains 70 mg/L of N applied as a 0.05% KNO3 (w/v) solution every week (Somasegaran and Hoben, 1994). Plants were supplied with tap water every two days and fertilized once a week with the quarter strength of N-free nutrient solution (Belay and Assefa, 2011). Plants growth were carried out in a greenhouse with a 12/12 h light/dark cycle. Finally, after 45 days of growth, all plants were harvested and the roots were scored for nodulation. The top plants and nodules were oven dried at 70°C for 48 h to determine the dry weight.
The percentage of isolates symbiotic effectiveness were calculated using equation proposed by Date et al. (1993) and indicated in Belay and Assefa (2011) with N2-fixing effectiveness classified as ineffective <35%; lowly-effective, 35 to 50%; effective, 50 to 80%; and highly effective, >80%.

SE = Symbiotic effectiveness; SDW = shoot dry weight.
Data analysis
Data analysis was done using one way analysis of variance (ANOVA) using version 20 SPSS statistical program. Mean separation was calculated using Tukey’s HSD test when the value was significant at p = 0.05.
RESULTS AND DISCUSSION
A total of 17 chickpea bacteria were recovered from the rhizospheric soil collected from chickpea farms of two purposely selected Districts (Dejen and Aneded) by using soil-host plant trap method. All the isolates were authenticated as chickpea rhizobia by re-inoculation test using sterilized sand-filled pot experiment.
Colony characteristics and dyes absorption ability of the isolates are summarized in Table 2. After 72 h of growth on YEMA medium, colonies were found to be large in size (3.0 to 5.0 mm), diameter, and showed large mucoid, watery, flattened and raised appearance similar to the findings obtained by Singh and Bamania (2012). Most of them were colorless and transparent, while some became yellowish after 3 days of the growth. The staining experiment also confirmed that all bacterial cell wall were stained as pink as the color of safranin and grouped under Grahams’ negative category (Agrawal et al., 2012). Furthermore, isolates showed considerable diversity on bromothymol blue (BTB) color conversion after 48 h of the growth. The isolates changed YEMA-BTB medium to yellow and deep yellow were categorized as fast growing and others which changed to moderate yellow and did not show any color change were considered as a slow growing rhizobia. Chickpea rhizobia was reported to have both fast and slow growing strains (Nour et al., 1994). Moreover, isolates obtained from chickpea nodules failed to grow on BTB-medium (Wei et al., 2003). Most of the isolates were Congo red dyes absorbent, except the three DMU-1, DMU-2 and DMU-3.
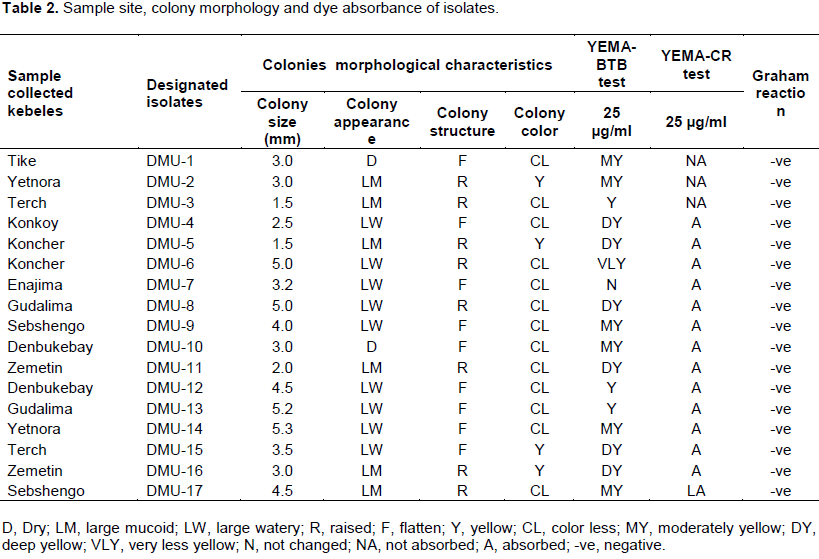
Edaphic condition of the soil is the most determinant factor for successful symbiotic association of Rhizobium with their host plants. Temperature, pH, salinity, antibiotic tolerance and carbohydrates utilization are important parameters to characterize rhizobia by consideration as a phenotypic identification marker (Maatallah et al., 2002). Temperature and pH tolerance of isolates is presented in Table 3. Chickpea rhizobia in this study showed a variation in these parameters. Almost all the isolates were grown at a temperature range of 5 to 40°C. Temperature tolerance of chickpea rhizobia ranging from 10 to 42°C has been already reported in India (Rai et al., 2012). Only five (DMU-1, DMU-2, DMU-10, DMU-14 and DMU-15) isolates were tolerant to temperature at 45°C. These isolates were expected to have high temperature resistancy and considered as an important candidate to develop inoculants as a bio-fertilizer. Furthermore, all the isolates grew well at pH range of 4.5 to 8.5 and this report is in line with findings of Kucuk et al. (2006), Baoling et al. (2007) and Singh and Bamania (2012).
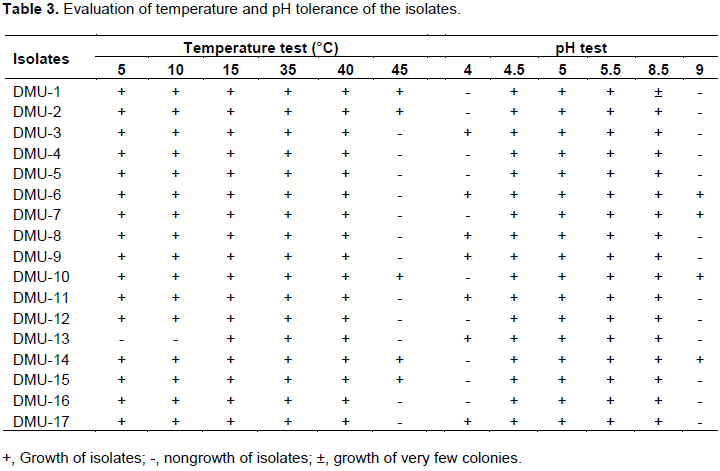
Most importantly, eight isolates (DMU-3, DMU-6, DMU-8, DMU-9, DMU-11, DMU-13 and DMU-17) showed their acidity tolerance by growing at pH 4. These isolates were considered as fast-growing strains as recognized from BTB-medium dyes conversion test and important candidate for acidic soil. This report is in agreement with findings of Gao et al. (1994) that showed that the rhizobia grown at pH as low as 4 were grouped under fast-growing strains whereas, four isolates such as DMU-6, DMU-7, DMU- 10 and DMU-14 were grown at pH 9. Some chickpea rhizobial isolates grew very well at pH 10 and tolerance to alkalinity increased at pH 11 (Singh et al., 2015). These alkaline condition preferring rhizobia were reported as slow-growing strains (Anand and Dogra, 1991). However, a number of reports indicated complete growth failures of chickpea rhizobia at pH of 9 (Kucuk et al., 2006; Baoling et al., 2007; Singh and Bamania, 2012). In this study, isolates showed edaphic factor tolerance diversity and similar with findings of Rai et al. (2012). Salinity test result also showed bacterial diversity towards different concentrations of the salt (Table 4). All the isolates were grown on the medium containing NaCl salt concentration ranging from 0.1 to 2%. Some isolates tolerated salt concentration upto 4%, while only a few isolates were grown at 5 and 6% of salt concentration. Most isolates are reported not to grow from 5% NaCl concentration and salt tolerance ability reduced with increase in salt concentration (Saraf and Dhandhukia, 2005; Singh and Bamania, 2012).
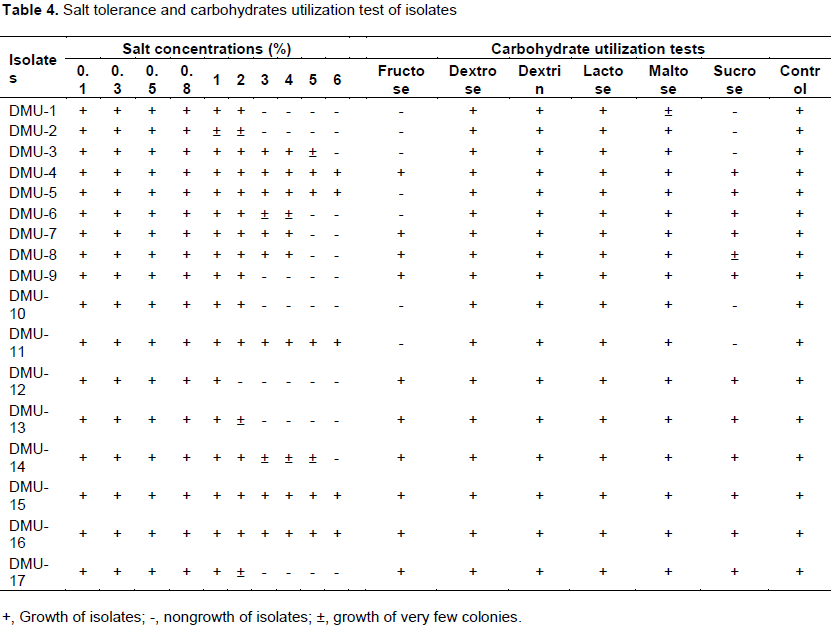
The isolates grown at 5 and 6% NaCl concentration were considered as salt tolerate and expected to have better adaptability to salty soil conditions. Therefore, isolates designated as DMU-4, DMU-5, DMU-11, DMU-15 and DMU-16 were grouped as salt tolerant rhizobial groups in this study.
Similarly, the salt tolerant isolates were better utilized among all tested carbohydrates. Only isolates DMU-5 and DMU-11 failed to grow on the disaccharide sugar, fructose, although tolerant to high salt concentration (Table 4). All the isolates were grown on dextrose, dextrin, lactose and maltose; however, isolates DMU-1, DMU-2, DMU-3, DMU-5, DMU-6, DMU-10 and DMU-11 failed to grow on the fructose and sucrose. With regards to this, there is well-established fact on Rhizobium utilization of various carbon sources for their growth and this is used as important tool to characterize the isolates (Maatallah et al., 2002). Rhizobial strains isolated from chickpea nodules were reported to utilize mannitol, lactose, sucrose, sorbitol, arabinose, galactose, mannose, maltose and raffinose as carbon sources (Singh and Bamania, 2012). Especially, fast-growing rhizobia were broadly recognized to grow on several types of carbon substrates, whereas slow growing rhizobia were grown only on very limited types of carbon sources. However, in this study, almost all isolates were grown on the tested carbohydrates and a broad range of carbohydrates was used as sources of carbon for growth. Hence in this regard, the result of this study is in line with the results of other studies (L’taief et al., 2007; Jida and Assefa, 2012). It is very interesting to note that chickpea Mesorhizobium can utilize a broad spectrum of carbohydrates for their cell growth and development. Such characteristics are usually used as diagnostic features for root nodule bacteria to test different carbon sources for their survival (Kucuk and Kıvanc, 2008).
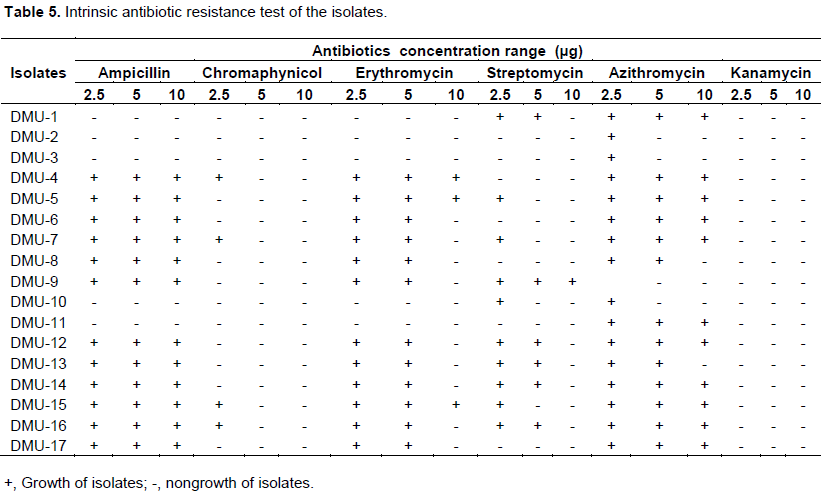
On the other hand, as summarized in the Table 5, majority of the isolates failed to tolerant several types of antibiotics in different concentrations sspecially isolates that were completely susceptible to kanamycin and only a few isolates were tolerant to chloramphenicol at 2.5 μgl-1 concentration. However, most of the isolates were grown on different concentrations of Ampicillin, Erythromycin and Azithromycin antibiotics. Isolates grown on the Streptomycin showed very less tolerance with respect to concentration. Several studies reported the existence of broad variation among chickpea rhizobia with respect to the fate of their intrinsic antibiotics resistance (Maâtallah et al., 2002; Kucuk and Kıvanc, 2008). The isolates’ sensitivity to antibiotics may be due to inability to resist exposed toxicity with less adaptation in natural environments (Singh and Bamania, 2012).
The legume food crop production was expected to be boosted by use of indigenous rhizobia as biofertilizer to supplement nitrogen requirement for cultivation of plants. Symbiotic effectiveness test was carried out to select the best N2-fixing strains among the obtained isolates (Table 6). In this greenhouse experiment, all the 17 isolates showed variation in host plant nodulation, with scores lesser (9 N/p) up to higher (22 N/p) nodules per plant. Although, plants showed a variation on nodulation, N2-fixation efficiency were found within effective up to highly effective ranges. Especially, shoot dry weight value was proved and considered as a direct indicator of isolates’ N2-fixation efficiency. Furthermore, this study showed the maximum (1.84 g/p) and minimum (1.18 g/p) shoot dry mass. For instance, isolate DMU-5, that scored high SDW of 1.84 g/p was highly effective (100%) and isolate, DMU-7 that scored SDW of 1.74 g/p was very effective (70%) on N2-fixation performance. Nitrogen fixation performance was positively associated with plant SDW (Qureshi et al., 2013). Nine isolates, namely DMU-2, DMU-3, DMU-4, DMU-5, DMU-6, DMU-7, DMU-8, DMU-9, and DMU-12 were potential N2-fixing isolates with highly effective (85 - 100%) fixation performance. The other isolates showed moderate percentage of N2-fixation performance variation. Such variation in each evaluated parameters were expected to depend on the chickpea bacterial diversity in the soil (Sahgal and Johri, 2003).
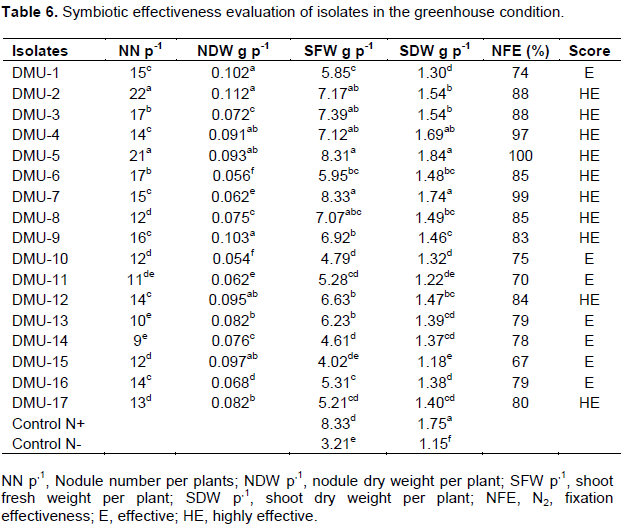
Some of the isolates that showed two or more environmental stress tolerance has attracted the interest of investigators in this study. Especially, four isolates such as DMU-6, DMU-10, DMU-14, and DMU-15 showed multiple abiotic stress tolerance, namely high temperature, pH and salinity, with scoped effective (67 to 78%) to highly effective (85%) performance of N2-fixation (Table 6).
Similarly, there are chickpea rhizobia which were reported from the alkaline condition of Indian soil (Singh et al., 2015). Therefore, these isolates were expected to be used as bio-fertilizer inocula in future, particularly in stressed farmlands to boost chickpea production and improve soil fertility. Inoculation with bacterial bio-fertilizer to farmland to improve crop production is not sole target rather it can have greate role to reduce the application of synthetic nitrogenous fertilizer on the farmland there by reducing pollution (Kennedy et al., 2004; Mia and Shamsuddin, 2010). However, biological N2-fixation (BNF) use has an incredible role in substituting commercially synthetic N-fertilizer in cereal production thereby, reducing the environmental problem (Agrawal et al., 2012). The dependency and high amounts of synthetic fertilizers application are both costly for farmers and set the hazardous problem on nature and biodiversity. Therefore, BNF provides a better alternative to chemical fertilizers as the process, besides supplying nitrogen to crop, enriches soil nitrogen content and maintains soil health and productivity (Reddy and Reddy, 2004).
CONCLUSION
As shown in the study, chickpea rhizobia isolated from rhizosphere soil showed variation in agro-ecological stresses tolerance. These isolates which were tolerant to edaphic stresses could be the potential asset for an alternative source of environmentally friendly bio-fertilizer and potential resources for varied agro-ecology. Isolates from this study showed sounding tolerance to temperature, pH and salinity and could have potential to tolerate environmental toxicity and hence increase N2-fixation effectiveness to enhance soil fertility in chickpea farming, thus increasing chickpea production. On the other hand, sensitive strains are least in tolerance to environmental toxicity and hence may not improve chickpea production. From this study, it could be deduced that nodulation performance of the rhizobia strain is positively correlated to N2-fixing effectiveness as well as higher shoots dry weight which confirms high assimilation of nitrogen of the chickpea seedlings. The isolates achievement on the N2-fixing process is very high and had better nodulation, and effective to highly effective fixation performances. Thus, this confirms the presence of potentially efficient chickpea rhizobia candidates in the rhizospheric soil of the study area although further work on filed condintion is needed.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
The authors gratefully acknowledge the financial support received from Research and Technology transfer Directorate Office of Debre Markos University in implementing this research. They also thank Dejene and Aneded District Agricultural and Rural Development Offices for providing various secondary basic data in support of the study and presenting chickpea farms of different kebele by considering different agroecological zones of the districts. Furthermore, they thank Adet Agricultural Research Center for providing good chickpea variety for this study.
REFERENCES
|
Agrawal PK, Agrawal S, Singh U, Katiyar N, Verma SK (2012). Phenotypic characterization of rhizobia from legumes and its application as a bioinoculant. J. Agric. Technol. 8(2):681-692. |
|
|
Anand RC, Dogra RC (1991). Physiological and biochemical characteristics of fast and slow growing Rhizobium spp. Pigeonpea (Cajanus cajan). J. Appl. Bacteriol. 70:197-205. |
|
|
Baoling H, Cheng QL, Bo W, Liqin F (2007). Rhizobia strain isolated from root nodule of gymnosperm Podocarpus macrophyllus. Sci. Chin. Ser. C-Life Sci. 50:1-6. |
|
|
Belay Z, Assefa F. (2011). The symbiotic and phenotypic diversity of Rhizobium leguminosarum by Viciae from Northern Gondar, Ethiopia. Afr. J. Biotechnol. 10(21):4372-4379. |
|
|
Beyene S, Dessalegn F, Worku W (2013). Improving Food Security in the Highlands of Ethiopia through Improved and Sustainable Agricultural Productivity an Human Nutrition. Final Technical Report. Hawassa University and University of Saskatchewan. |
|
|
Beynon JL, Josey D P (1980). Demonstration of heterogeneity in a natural population of Rhizobium phaseoli using variation in intrinsic antibiotic resistance. J. Gen. Microbiol. 118:437-442. |
|
|
Christodoulou V, Bampidis VA, Hucko B, Ploumi K, Iliadis C, Robinson PH, Mudrik Z (2005). Nutritional value of chickpeas in relations of lactating ewes and growing lambs. Anim. Feed Sci. Technol. 118:229-241. |
|
|
CSA (Central Statistical Authority) (2011). Agricultural samples survey 2010/2011: Report on area and production of major crops. Vol. 1. Statistical Bulletin, Addis Ababa, Ethiopia. |
|
|
Funga A, Ojiewo OC, Turoop L, Mwangi SG (2016). Symbiotic effectiveness of elite rhizobia strains nodulating desi type chickpea (Cicer arietinum L.) varieties. J. Plant Sci. 4(4):88-94. |
|
|
Gao JI, Sun JG, Li Y, Wang ET, Chen WX (1994). Numerical taxonomy and DNA relatedness of tropical rhizobia isolated from Hainan Province, China. Int. J. Syst. Bactriol. 44:151-162. |
|
|
IFPRI (International Food Policy Research Institute) (2010). Pulses Value Chain Potential in Ethiopia: Constraints and opportunities for enhancing exports, P 12. |
|
|
Imran A, Mirza SM, Shah MT, Kauser A, Malik AK, Hafee YF (2015). Differential response of kabuli and desi chickpea genotypes toward inoculation with PGPR in different soils. Front. Microbiol. 6:859-873. |
|
|
Jida M, Assefa F (2012). Phenotypic diversity and plant growth promoting characteristics of Mesorhizobium species isolated from chickpea (Cicer arietinum L.) growing areas of Ethiopia. Afr. J. Biotechnol. 11(29):7483-7493. |
|
|
Keneni G, Bekele E, Getu E, Imtiaz M, Dagne K, Assefa F (2011). Characterization of Ethiopian chickpea (Cicer arietinum L.) germplasm accessions for response to infestation by Adzuki Bean Beetle (Callosobruchus chinensis L.) performance evaluation. Ethiopia. J. Agric. Sci. 21: 65-83. |
|
|
Kennedy IR., Choudhury, ATMA, Kecskés ML (2004). Non-symbiotic bacterial diazotrophs in crop-farming systems: can their potential for plant growth promotion be better exploited? Soil Biol. Biochem. 36:1229-1244. |
|
|
Kucuk C, Kivanc M (2008). Preliminary characterization of Rhizobium Strains. isolated from chickpea nodules. Afr. J. Biotechnol. 7(6):772-775. |
|
|
Kucuk C, Kıvanc M, Kınacı E (2006). Characterization of Rhizobium sp. isolated from bean. Turk. J. Biol. 30:127-132. |
|
|
L'taief B, Sifi B, Gtari M, Zaman-Allah M, Lachaal M (2007). Phenotypic and molecular characterization of chickpea rhizobia isolated from different areas of Tunisia. Can. J. Microbiol. 53:427-434. |
|
|
Lupwayi NZ, Haque I (1994). Legume-Rhizobium Technology Manual. Environmental Science Division, International Livestock Center for Africa, Addis Ababa, Ethiopia, p. 97. |
|
|
Maâtallah J, Berraho EB, Sanjuan J, Lluch C (2002). Phenotypic characterization of rhizobia isolated from chickpea (Cicer arietinum) growing in Moroccan soils. Agronomie 22:321-329. |
|
|
Mia, MAB, Shamsuddin, ZH (2010). Rhizobium as a crop enhancer and biofertilizer for increased cereal production. Afr. J. Biotechnol. 9(37):6001-6009. |
|
|
Miller P, McKay K, Jenks B, Riesselman J, Neill K, Buschena D, Bussan JA (2002). Growing Chickpea in the northern Great Plains. Montana State University, pp. 1-8. |
|
|
Mohammed I, Ahmed AR, Senge B (2011). Dynamic rheological properties of chickpea and wheat flour Doughs. J. Appl. Sci. 11:3405-3412. |
|
|
Naser MS, Mohammad C, Ali AS, Ali MA, Abolfazl AG (2008). Nutritional evaluation of kabuli and desi type chickpeas (Cicer arietinum L.) for ruminants using in vitro gas production technique. Afr. J. Biotechnol. 7(16):2946-2951. |
|
|
Nour SM, Cleyet-Marel JC, Beck D, Effosse A, Fernandez MP (1994). Genotypic and phenotypic diversity of Rhizobium isolated from chickpea (Cicer arietinum L.). Can. J. Microbiol. 40(5):345-354. |
|
|
Qureshi TS, Bux H, Khan RM (2013). Symbiotic characterization of rhizobia nodulating Cicer arietinum L. isolated from Pakistan. Int. J. Agro. Plant Prod. 4(11):2912-2918. |
|
|
Rai R, Dash KP, Mohapatra T, Singh A (2012). Phenotypic and molecular characterization of indigenous rhizobia nodulating chickpea in India. Int. J. Exp. Biol. 50:340-450. |
|
|
Reddy BG, Reddy MS (2004). Soil health and crop productivity in alfisols with integrated plant nutrient supply system. Proceedings of the Australian Agronomy Conference, Australian Society of Agronomy, Perth, W.A., pp. 102-104. |
|
|
Sahgal M, Johri BN (2003). The changing the face of rhizobial systematics. Curr. Sci. 84:43-48. |
|
|
Saraf M, Dhandhukia P, (2005). Response of Sinorhizobium meliloti to high salt concentration and effect of added osmotica. J. Microbial World 7:250-257. |
|
|
Shiferaw B, Teklewold H (2007). Structure and functioning of chickpea markets in Ethiopia: Evidence-based on analyses of value chains linking smallholders and markets. Improving Productivity and Market Success (IPMS) of Ethiopian Farmers Project Working Paper 6. ILRI (International Livestock Research Institute), Nairobi, Kenya. P 63. |
|
|
Simon T, Mtei K, Amare GA, Ndakidemi AP (2014). Isolation and characterization of nitrogen-fixing rhizobia from cultivated and uncultivated soils of Northern Tanzania. Am. J. Plant Sci. 5:4050-4067. |
|
|
Singh KAG, Bamania M (2012). Characterization of Mesorhizobium sp. isolated from root nodules of Cicer arietinum. Int. J. Agric. Sci. Res. 2(3):142-154. |
|
|
Singh PR, Manchanda G, Singh NR, Srivastava KA, Dubey CR (2015). Selection of alkalotolerant and symbiotically efficient chickpea nodulating rhizobia from North-West Indo Gangetic Plains. J. Basic Microbiol. 56:14-25. |
|
|
Singh S, Singh I, Kapoor K, Gaur MP, Chaturvedi KS, Singh PN, Sandhu SJ (2014). Chickpea: broadening the genetic base of grain legumes. Springer. pp. 51-75. |
|
|
Somasegaran P, Hoben HJ (1994). Handbook for Rhizobia. Springer- Verlag. |
|
|
Tena W, Wolde-Meskel E, Walley F (2017). Response to chickpea (Cicer arietinum L.) to inoculation with native and exotic Mesorhizobium strains in Southern Ethiopia. Afr. J. Biotechnol. 15(35):1920-1929. |
|
|
van der Maesen LJG (1987). Origin, history, and taxonomy of chickpea. In: Saxena MC, Singh KB (eds) The chickpea. C.A. B., Wallingford, pp. 11-34 |
|
|
Vincent JM (1970). A Manual for the Practical Study of Root-Nodule Bacteria. Black well Scientific, Oxford. |
|
|
Wei GH, Tan ET, Zhu ME, Wang ET, Han SZ, Chen WX (2003). Characterization of rhizobia isolated from legume species within the genera Astragalus and Lespedeza grown on the Loess Plateau of China and description of Rhizobium loessense sp. Int. J. Evol. Microbiol. 53:1575-1587. |
|
|
Werner D (2005). Production and biological nitrogen fixation of tropical legumes. In: Werner D and Newton W E. (eds) Nitrogen Fixation in Agriculture, Forestry, Ecology, and the Environment, Springer, Netherlands, pp. 1-13. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


