Full Length Research Paper

ABSTRACT
Chlamydomonas reinhardtii CC125 (wild type) and CW15 (cell wall mutants) were feed up on solid and liquid Tris phosphate (TP) media with various concentrations of acetate, glycerol(10-100 mM) or methanol (0.01-718 mM) and cultivated under phototrophic, mixotrophic and heterotrophic conditions. Use of 10 and 35 mM acetate and 10 and 50 mM glycerol under constant 38 µE m−2 s−1 light illumination (mixotrophically) was the optimum condition for both strains to have a higher biomass and growth compared to other carbon sources and concentrations. Both strains had a quicker growth rate in just 35 mM of acetate and 10 mM glycerol although feed of algal cells on 35 mM acetate produced more and quicker biomass. In use of 10 mM acetate in micro plate and tissue culture flasks, CW15 had a maximum growth rate of 5.3×104 and 1.3×104 cells/hour; while on use of 35 mM acetate, the growth rate was 8.8×104 (micro plate) and 4.0×104 cells/hour (tissue culture flasks). Wild type had a maximum 2.7×104 (micro plate) and 4×103 (tissue culture) cells/hour in use of 10 mM acetate. In feed of CC125 with 35 mM acetate, growth rate correspondingly for micro plate and tissue culture flasks was 2.5×104 and 2.6×104 cells/ hour. Among the two strains, CW15 with specific growth rate of 8.8×104 cells/hour (in micro plates) and 4.0×104 cells/hour (in tissue culture flasks) on 35 mM acetate also grew quicker than CC125. Susceptibility to bacterial contamination was checked on both strains and we also found that, just as the absence of a cell wall in CW15 accelerated the growth, it also appeared to increase the chance of contamination by about twofold compared to the wild type but this can be minimized by the use of antibiotics in the growth media.
Key words: Heterotrophic growth, mixotrophic growth, acetate, glycerol methanol Chlamydomonas, CC125, CW15.
Abbreviation: TAP, Tris-acetate phosphate; TP, Tris-phosphate; TPA, TP+2% agar; DMR, digital module R; TGA, triglyceride.INTRODUCTION
Reducing the cost of bio-products processes from its earlier stage to the final one is a vital factor that has been carried out throughout the world. Although discovery of some bacteria such as Escherichia coli
reduces bio-process applications for many decades, it is time to substitute an alternative organism in which to mount the yield of biomass with minimal inputs and costs. Microalgae are a kind of organism that seems to have this functionality in addition to other advantages ranging from (1) their accessibility from fresh water and soil, (2) photosynthetic ability, and, (3) well-annotated genomes such as Chlamydomonas reinhardtii, Chlorella vulgaris, Volvox carteri, Chlorokybus atmophyticus, and Dunaliella salina to just name few.
The green algae C. reinhardtii among other micro algae specious has been utilized as a model species in many bio-applications (Roacha et al., 2013; Fischer et al. 2006; Ledford et al., 2007; Peers et al., 2009). Generally, it can grow phototrophically and this means it uses incident light and CO2 and converts it to sugar and O2.Some strains of C. reinhardtii have the capability to grow mixotrophically and/ or heterotrophically (Brennan and Owende, 2010; Dragone et al., 2010). Despite heterotrophic feed of microalgae that let algal cell consume solely the external organic carbon source, microalgae which can grow under mixotrophic growth condition can form synthesized carbon source and has both photosynthetic and heterotrophic characteristic. However, in mixotrophic growth regime microalgae consume simultaneously light and external organic carbon source for its growth (Dragone et al., 2010).
Production of microalgae in large-scale is somehow considered as a key parameter for industrialization and commercialization of microalgae and has been studied for decades (Becker, 1994). The ease of its, well annotated genomes and cells behavioral make it as a remarkable laboratory tool to be used in wide range variety of either practical or potential products ranging from lipid, enzymes, isoprenoids, polysaccharides cultivation as well as the production of biofuels as a green and substitute energy source to traditional fossil fuels (Brennan and Owende, 2010).
The achievement of above mentioned product (enzymes, biofuel, lipid and polysaccharides cultivation), mostly was occurred by cultivating the microalgae on varied mineral media, organic substrates, and synthetic or real wastewaters (Pulz, 2001; De-Bashan et al., 2002; De-Bashan et al., 2004; Pulz, 2004; Lebeau et al., 2006; Harun et al., 2010).
Today, growth of micro algae under phototrophic condition is the most common method compared to mixotrophic and heterotrophic conditions. However, the most common one does not necessarily mean the best one, as literatures has shown that the use of both light and external carbon source in mixotrophic condition not only causes a higher yield but also reduces the cost of cultivation compared to phototrophically growth condition (Bhatnagar et al., 2011; Cerón García et al., 2005) and this is commercially quite vital (Abad and Turon, 2012; Liang et al., 2009).
Also, Yu et al. (2009) (Shi et al., 1999) has shown that growing algae via heterotrophic and mixotrophic conditions can overcome such disadvantages of phototrophic growth (low cell densities and longtime cultivation) and the culture conditions for optimum growth and yield rate have not been suggested therefore a think of suitable organic carbon among several carbon sources can be challenging and crucial. To our knowledge, the most common used organic carbon sources for hetero-trophic and mixotrophic algal growth are glucose, acetate, sucrose, lactate, lactose, ethanol and glycerol (Shi et al., 1999; Chen and Johns, 1996; Octavio et al., 2010; Zhang et al., 1999).
Despite the importance of right carbon sources and right growth conditions, several factors such as algal strains with various characteristic and behavior, pH, oxygen level, association with or without bacteria or fungi, temperature and introduction of adequate light intensity (Ras et al., 2013; Spreitzer et al., 1998; Falk et al., 2006) to permit a faster algae growth are key objectives that may alter the biomass yielding (Mandalam and Palsson, 1998; Yang et al., 2000; Suh and Lee, 2003). Here our aim is to examine the wild type (CC125) and the cell wall mutants (CW15) of C. reinhardtii algae grown on different types of carbon sources in particular (acetate, glycerol and methanol) at various concentration in both dark and light conditions to suggest an optimal culture condition for achieving a higher growth yield and biomass.
Acetate (or acetic acid) is one of well-known carbon sources that its use has been generally accepted for variety of both microbial species and microalgae (Droop, 1974; Moon et al., 2013). Generally, acetate can be metabolized through two routes when it has been carried by coenzyme A. The first pathway will occur from glyoxylate cycle in order to produce malat and the second pathway that can provide the energy (ATP and NADH) and carbon skeletons through the tricarboxylic acid (TCA) to citrate cycle. Solely, those micro algae species that has the first metabolic pathway can grow as the two crucial enzymes isocitrate lyase and malate synthetase (which themselves are necessary for operating the glyoxylate cycle) later will be stimulated in algal cells when they are in acetate media (Neilson and Lewin, 1974; Boyle and Morgan, 2009).
Albeit solely some types of micro algae such as Phaeodactylum tricornutum, Nannochloropsis sp., Rhodomonas reticulate, and Cyclotella cryptica can consume glycerol as a carbon source, optimizing the use of glycerol for a faster growth and higher yield in mixotrophic condition has been done for decades (Choi et al., 2011; Cero´n Garcı´a et al., 2000). It is a by-product of biofuel production and a good source for preservation of microorganism including microalgae at low temperatures (Moon et al., 2013). Despite acetate that in high concentration will caused cells degradation due to its acidity and toxicity characteristic, glycerol has not have any toxicity effect on cells even at higher concentration (Kaplan et al., 1986) and so far several efforts have been done to utilize this non-toxic organic carbon source in microalgae growth.
Methanol is the most basic form of alcohol and in its use in biofuel processing it plays a key role to convert triglycerides (TGA) into biodiesel through a mechanism called transesterification. Introducing the methanol into algal system for instance (0.01-718 mM) compared to other carbon sources (glucose, acetate glycerol) has an advantage as it can sterilize the media and therefore minimize bacteria and fungi contamination in open ponds (Choi et al., 2011). Methanol frequently has been used in commercial purposes due to its low cost and the fact that is a biofuel by-product. In a study by Choi et al. (2011), it was shown that growth of Chlorella in 1% (v/v) methanol can increase the lipid yield and growth rate (Moon et al., 2013). However, the use of methanol either mixotrophically or heterotrophically has not been reported as an organic carbon source has been reported a level of flocculation in C. reinhardtii algal cells (cite methanol algae article). By consideration of merit and demerit points of each mentioned organic carbon source in this study we will conclude the most efficient carbon source for faster growth and yield.
MATERIALS AND METHODS
Solid cultures
In a 500 ml bottle, Tris-Phosphate in absence of acetic acid medium (TP) (Gorman and Levine, 1965) was mixed with 2% Difco agar and autoclaved for 15 minute at 121°C. Then acetate and glycerol with various concentrations (10, 20, 35, 50 and 100 mM) were individually added in a separate 500 ml TP + 2 % agar media and they also autoclaved for 15 min at 121°C.
Methanol (0.01, 25, 123, 245 and 718 mM) was added to the sterilized TP media. Each of the above prepared media (TP, Acetate, Glycerol and Methanol) was poured in 8 Petri dishes (two for each strain one for light and one for dark condition). Each axenic algal strain which was initially grown on TAP media + 2% agar and was taken from Professor Colin Robinson’s lab, inoculated and streaked into the above solid prepared TP, TP-Acetate, TP-Glycerol, TP-Methanol media and was grown for 7 days in light with intensity of 38 µE m−2 s−1 and dark condition.
Micro-plate
Into two 24 wells micro plates (Cellstar company) 1.5 ml of TP (as phototrophic control), Acetate or Glycerol (10, 20, 35, 50 and 100mM) and (0.01, 25, 123, 245 and 718 mM) of methanol independently was pipetted twice. Algal cells were inoculated and mixed in a separate 1.5 ml Eppendorf tube containing sterilized TP media and then 50 µL of each strain individually was pipetted into each well of the micro- plate with specific mentioned acetate, glycerol and methanol concentration. For mixotrotrophic growth one of the two microplates was put under light intensity of with 38 µE m−2 s−1 and the other one covered with aluminum foil for heterotrophic purposes. Both micro-plates then were put in the algae room on a separate Panasonic double orbital shaker with speed of 170 rpm for 7 days. Once cells were grown for one week, growth rate were visualized and only acetate and glycerol mixotrophically seemed to be utilized by algal cells with certain concentrations seemingly be used more efficiently, that is, 10 and 35mM of acetate and 10 and 50mM of glycerol. Consequently, each strain once more was grown at that efficient concentration of carbon sources in triplicate in a light intensity of 38 µE m−2 s−1 in both 24 well micro plate. Daily biomass rate was checked by a Panasonic micro plate reader in 600 nm wave length. Sample also was taken from each micro plate well and cells counted by Neubaur hemocytometer (Figure 5, part C and D, illustrate the callibration of cell numbers with absorption in 600nm).
Tissue culture flasks
To find out a possible role of different shaking speed in algal respiration system, the experiment was repeated at half the speed of 24 wells micro plate (70 rpm) but in tissue culture flasks. A set of three of Sigma-Aldrich 25 cm3 tissue culture flasks independently was used for each concentration of chosen carbon source. Each set then separately filled with 25 ml of prepared TP with (10 and 35 mM) acetate and (10 and 50 mM) glycerol. In each flask, 100 µl algae cells from each strain was pipetted. Flasks were thereafter put on a 70 rpm Panasonic double orbital shaker for 7 days. Cell counting, optical density at 600 nm (Figure 5, part A and B, display the callibration of cell numbers with absorption in 600nm) and chlorophyll measurement was measured daily.
Chlorophyll extraction from tissue culture flasks
One milliliter from each tissue culture flasks containing algal cells was pipetted into individual 1.5 ml centrifuge Eppendorfs and centrifuged for 5 min at 6000 rpm. Supernatant was removed and algal cells vortexed with 1 ml 95% ethanol (v/v) for about 45 s. Eppendorfs were again centrifuged for 4 min at 13,000 rpm. The remaining supernatant containing extracted algal chlorophyll was transferred in 1 ml plastic cuvette. The spectrophotometer was blanked against 95% ethanol before each absorption at 649, 665 and 750 nm.
Chlorophyll a counting
The rate of produced chlorophyll a calculated via the below equation (Bergmann and Peters, 1980):
.png)
Where, V is the volume of extract in ml; V is the volume of sample filtered in; l is the length of cuvette in cm.
Algae strains from solid TAP media were inoculated and mixed with 1.5 ml fresh liquid TP media and vortexed for 2 min. Algae cells were diluted 100 times and cell density was measured by counting the cells in a hemocytometer (CW15 and CC125 respectively had 1.25×106 cells/ml and 5×105 cells/ml). Our aim was to observe 3000 bacterial colonies in use of 1ml algal culture meaning in 0.1 ml of algae culture we would expect to see 300 bacteria colonies. Samples were diluted appropriately to give 300 colonies per plate. Two percent agar from Diefco Company was poured individually in a 100 ml of TP, TAP and TSB (Tryptone-Soya Broth) and autoclaved as before.
The autoclaved media independently poured in 4 individual sterilized petri dishes. 100 µl diluted sample were pipetted into 4 individual solid TP, TAP and TSB petri dishes and distributed perfectly in all petri dishes. While TSB petri dishes were kept in a 30°C incubator, TP and TAP Petri dishes were kept in 27°C algae growth room with 38 µE m−2 s−1 light intensity.
Physical morphology
Both algal strain which grown without additional of any carbon source in solid and liquid TP, TAP were visualized under the Leica DMR (Digital Module R) microscope with 1000 magnification and compared to the grown algae cells in various concentration of acetate, glycerol (10, 20, 35, 50 and 100 mM).
RESULTS AND DISCUSSION
Physical morphology
Visualization of both strains under light microscope with 1000X magnification showed the existence of cell wall and flagella respectively in CC125 and its absence in CW15. Despite having an approximate same size (50 µm) and circular shape of both strains, cells in CC125 mostly joined together as a colony in a capsulated environment by their cell wall while the CW15 cells remained separate from each other.
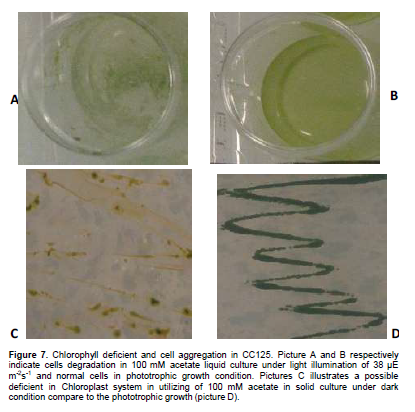
Moreover, further microscopy of the grown CC125 algal cells either mixotrophically or heterotrophically with different concentrations of acetate (10, 20, 35, 50 and 100 mM) indicated some valuable results. For instance, in dark growth conditions using 50 and 100 mM acetate the color of existed chlorophyll in CC125’s cells mostly was bright yellow rather than green (Figure 7, part C and D clearly show the differentiate color of chlorophyll).
The reason that chlorophyll looks green is that it absorbs other colors existed in light (red and blue) except green. Also, in low light intensity more rate of chlorophyll will expect to produce. And the more chlorophyll means more green color (Lange et al., 1981). Therefore, this may suggest an existence of a deficient in CC125 chloroplast pigments (chlorophyll a, b or xanthophyll) in which the absence of light and high rate of concentration (50 and 100 mM) inhibits algal cells to either absorb the light or consume high concentrated acetate for growth. Another possibility which also was seen in 24 wells micro plate can be cell degradation and death due to the high toxicity of 50 and 100 mM acetate. Figure 7A shows cell degradation and death while part B display steady cell aggregation). We also found that in heterotrophic use of 50 and 100 mM acetate most of CW15 algal cells lost their circular shapes and instead formed some unstructured shapes. In Figure 1, a noticeable difference in shape of CW15 in use of low acetate concentration (B) and high acetate concentration (D) was observed.
.png)
The optimum condition and concentration for acetate in solid and media culture
From the data gathered after a period of seven days we found that mixotrophic growth of both strains was much higher than growing under either phototrophic or heterotrophic conditions. Moreover, mixotrophic growth of algal cells on acetate, glycerol and methanol revealed that acetate was the carbon source that was consumed better compared to the others. Our experiment also showed that the optimum concentration of acetate for improving the yield was 35 Mm. Figure 2A indicates a higher mixotrophic growth compared with other carbon sources (C&E)) and higher concentration of acetate (100 mM) caused cell death and degradation specifically in CC125 which might be due to the toxic effect of 100 mM acetate on algal cells. A rate of flocculation also was seen on CW15 when a range of methanol concentration (0.01, 25, 123, 245 and 718 mM) was used mixotrophically (Scholz et al., 2011).
.png)
.png)
.png)
.png)
.png)
The maximum growth rate of CW15 grown on micro plates in feed of 10 and 35 mM acetate correspondingly was 5.3 × 104 and 8.8 × 104 cells/hour. The wild type (CC125) showed less growth rate, being 2.7 × 104 (10 mM) and 2.5 × 104 (35 mM) cells/hour. Figure 3A and B show algal cell growth in 10 and 35 mM acetate.
.png)
.png)
.png)
.png)
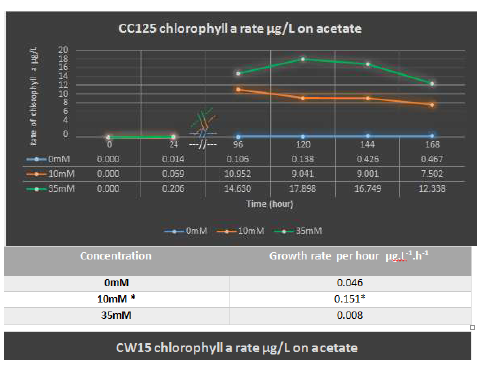
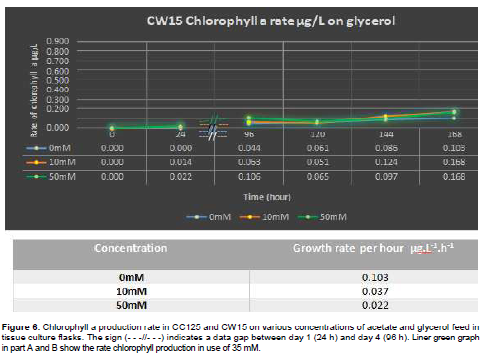
Results given from tissue culture flask also has shown that regardless of shaking speed and its possible effect on algal respiration system, CW15 grew quicker on 35 mM acetate with maximum rate of 4.0 × 104 cells/hour than CC125 with maximum rate of 2.6 × 104 cells/hour. Figure 4C and D reflect the faster growth of CW15 on 35 mM acetate feed. The CW15 also had a faster chlorophyll production rate of 0.046 µg.L-1.h-1 compared to CC125 with a rate of 0.008 µg.L-1.h-1 (Figure 6).
.png)
.png)
.png)
.png)
.png)
.png)
.png)
.png)
.png)
We also found that in mixotrophical use of 35 mM acetate both strains produced a higher biomass than other concentrations and conditions. The maximum rate of extracted chlorophyll a, for instance, in 35 mM acetate was 17.89 µg.L-1.h-1 (CC125) and 12.516 µg.L-1.h-1 (CW15) which was noticeably higher than the phototrophic yield with rates of 0.467 µg.L-1.h-1 (CC125) and 0.103 µg.L-1.h-1 (CW15). Utilization of 10 mM acetate however produced lower chlorophyll a than 35 mM acetate but higher than phototrophic growth as CC125 had 10.95 µg.L-1.h-1 and CW15 had solely 4.758 µg.L-1.h-1.
The optimum condition and concentration for glycerol
In mixotrophic utilization of 10 and 50 mM glycerol however the outcomes from micro plates and tissue culture flasks were considerably different. In micro plates the maximum growth rate of CW15 respectively for 10 and 50 mM glycerol was 5.2 × 104 and 7.5 × 104 cells/hour while in tissue culture flasks cells were grown better on 10mM than 50mM as 2.4 × 104 and 1.3 × 104 cells/hour respectively were generated
The CC125 we had a higher growth in 10 mM glycerol in both micro plates and tissue culture flasks. The maximum growth rate in micro plate and tissue culture flasks was 2.0 × 104 and 7.8 × 104 cells/hour respectively. In 50 mM glycerol media results were 1.2 × 104 cells/hour in micro plate and 2.8 × 104 cells/hour in tissue culture flasks. This is how the maximum phototrophic growth rate in CC125 and CW15 in two times replicate respectively was 3.4 × 104 and 5 × 103 cells/hour and 4.9 × 104 and 6.5 × 104 and 2.3 × 104 and 1.1 × 104 cells/hour (Figures 3 and 4.).
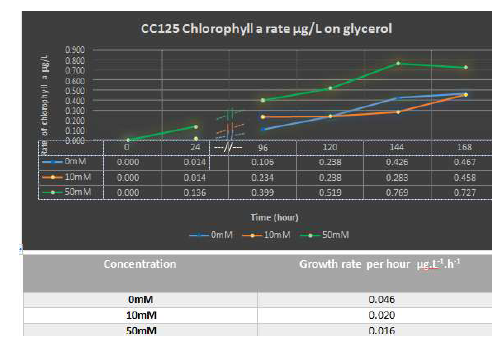
Extraction of chlorophyll from algal cells were grown on tissue culture flask also showed that CW15 and CC125 both had a faster chlorophyll production growth rate in 10 mM glycerol with corresponding rate of 0.037 (CW15) and 0.020 (CC125) µg.L.1 h1 than feed on 50 mM glycerol with maximum production growth rate of 0.022(CW15) and 0.016 µg.L.1 h1 (CC125) (Figure 6C and D). However, the rate of produced biomass (chlorophyll a amount) in use of 50 mM glycerol was higher in CC125 than 10 mM (0.769 and 0.458 µg.L.1 h1, respectively). The cell wall mutants produced a same amount of chlorophyll a in use of 10 and 50mM glycerol (0.168 µg.L.1 h1). Likely to use of acetate, feed of algal cells on glycerol culture either in 10 or 50 mM concentration, showed a higher yield than phototrophic condition with rate of 0.467 µg.L.1 h1 (CC125) and 0.103 µg.L.1 h1 (CW15).
The data suggests that growing CC125 on small sized culture such as micro plate may not be industrial while the growth rate of phototrophic was higher than 10 and 50 mM glycerol however in bigger culture (tissue culture flasks) the use of just 10 mM glycerol can be quite interesting.
Economics matter in use of 10 or 50 mM glycerol
An important point in yield and growth of algae is economic matters in which to reduce the cost of experiment as much as possible and introduce the most optimum industrialized system. Although CW15 had a higher growth rate in micro plate on feed of 50 mM glycerol, as commercial fermenters are much larger than micro plate and as we got a higher growth rate in use of 10 mM glycerol in tissue culture flasks as an example of bigger culture, so utilizing the 10 mM glycerol commercially and economically is more beneficial as its cost is 1/5 of 50 mM glycerol. The cost of at least minimum volume of 1000 L 10 and 50 mM glycerol solution will be, $360 and $1802, respectively.
Susceptibility to bacterial contamination
The rates of algal mixotrophical growth on solid (TPA) and liquid TP cultures were different. On solid TP plates, the growth ratio of CC125 was higher compared to CW15 in both acetate and glycerol (Table 1). In the susceptibility to bacterial contamination experiment that was done on both strains, we found that on TP media CW15 had 28 out of 300 bacterial colonies (10 %) and CC125 had 20 out of 300 colonies (6.6%) whereas on TAP media CW15 and CC125 got 23 out of 300 (7.6%) and 9 out of 300 (3%) bacteria colonies, respectively. On TSB plates, however, solely 2 out of 300 (0.6%) and 1 out of 300 (0.3%) of bacteria colonies were seen on CW15 and CC125 in orderly fashion. This can suggest that the lack of cell wall would increase the chance of bacterial contamination almost 2 times more than the wild type with its cell wall as a barrier.
Moreover, the rate of fungal contamination of CW15 on TSB plates was much higher than CC125 compared to other TP and TAP media cultures.
Thus, one possibility could be that it was as a result of the negative effect of bacteria on growth of CW15 on solid culture. Hence, when cells are introduced in a soluble medium, researchers should not always expect to see the same behavior as to when such cells were in solid media. In shaking liquid culture, there is a possibility that micro gradients are introduced in a watery atmosphere and therefore a lower potential for neighbor effects and bacteria interactions will occur compared to solid media.
.png)
Further discussion and future applications
The potential of micro algae as laboratory tool in cosmetic industries (Potvin and Zhang, 2010), drug designing (Bumbak et al., 2011; Milledge, 2010) and recombinant proteins or as a renewable energy source specifically in biofuel production make it an interesting microorganism to invest more attention and effort on its growth and yield (Chisti, 2007; Stephens et al., 2010; Wijffels and Barbosa, 2010; Hempel et al., 2010). So far we showed that mixotrophically growth of CC125 and CW15 on 35mM acetate was the optimum way for faster growth and biomass rate. By knowing that each algal strain behave differently in different media and conditions and there was not any fixed pathway so far therefore a need for further researches and experiments is vital to confidently propose a system with highest and fastest biomass production.
In theme of biofuel one possible application for future purposes can be use of organic (acetate) and inorganic carbon (bicarbonate) in TP media to see its effect on both growth and lipid production.
As it is in general acceptance that for commercial purposes inorganic carbons are needed and as (Gardner et al., 2012; Gardner et al., 2013) have shown that bicarbonate as a lipid trigger can increase the yield of lipid production such as triacylglycerol (TAG) in some microalgae specious(Gardner et al., 2012; White et al., 2012) thus it can worth to determine the outcomes of our experiment (use of 35 mM acetate for faster growth) to other literatures results as a new application to find out the possible rate of lipid production specifically in CW15 that lacks cell wall. The absent of cell wall in CW15 not only has shown a faster growth but also can speed up the cultivation process. What also we really interested in to examine in our future experiment is correlating the effect of various light intensity and temperature (Ras et al., 2013; Spreitzer et al., 1998; Falk et al., 2006) on growth and lipid (TAG) production of our used algal cells (CW15 and CC125).
Moreover, as the rate of bacterial contamination was higher in CW15 than CC125 due to the absence of its cell wall, one interesting application could be the use of bacterial 16srRNA probes to investigate a possible interaction between CW15 and bacteria and the role of bacteria in rate of biomass. The use of this small ribosomal ribonucleic acids subunit (16 srRNA) still is one of the precise method for identification of bacteria as they are conserve molecules and present in all bacteria thus for identification of different bacteria still the recognition system will remain fixed (Ludwig et al., 1998).
CONCLUSION AND RECOMMENDATION
The potentiality of microalgae to substitute as renewable energy source to some conventional ways made it interesting. So far micro algae has been cultivated in various conditions (phototrophic, mixotrophic and heterotrophic), and nutritious feed, but each has some merit and demerit points. Introducing algal cells in phototrophic condition minimizes the growth rate, but its advantage is to also decrease the possibility of bacterial contamination. Mixotrophic condition can boost the algal growth and yield due to use of addition of external carbon source and light simultaneously, but also can encourage bacteria into the system due to the existed carbon source. On the other hand, in heterotrophic conditions although the cost of experiment clearly is less than both mixotrophic and phototrophic conditions, because the created dark atmosphere in addition of external carbon is strongly a suitable condition for bacterial and fungus growth the chance of algal yield and growth also will decrease. Moreover, solely some specious of algae have capability to grow on dark condition. Despite the advantage and disadvantage of each growth condition our results suggested that acetate was the most effective organic carbon source among here tested carbon sources and the optimal concentration for C. reinhardtii CC125 (wild type) and CW15 (cell wall mutants) to grow quicker was use of 35 mM acetate mixotrophically.
Also, another recent study done by Moon et al. in 2013 showed that among acetate, glucose, glycerol or sucrose, mixotrophically growth of C. reinhardtii on acetate was the optimum way in terms of growth and lipid yield. Also, use of 35 mM acetate in 1000 L fermenter is economically even cheaper than use of the alternative carbon source (10 mM glycerol), being $327 and 363, respectively.
Even though in our experiment a minimum growth rate was seen in feed of both C. reinhardtii strains (CC125 and CW15) on glycerol culture (Cerón García et al., 2000) has shown that the use of 0.1M glycerol with light intensity of 165 µE m-2s-1 let Phaeodactylum tricornutum grow 74% faster than phototrophic condition.
Comparatively to the wild type (CC125), our experiment suggests that the lack of cell wall in CW15 possibly decrease the digesting process time and this can be the reason for a faster growth rate (cells or chlorophyll a) per hour in liquid culture. However, in absence of antibiotics in media the chance of bacterial contamination is approximately 2 times higher in CW15 compare to CC125 that has cell wall as a barrier.
Though the cells morphology data shows that a single algae cell in both CC125 and CW15 has a ring like shape with an approximate same size of 50 µ, biotechnologically growing and cultivating CW15 can be preferred since cells in CW15 cells remain separately from each other while in CC125 mostly join together as a colony in a capsulated environment by their cell wall and this means more small sized cells are better and offer a possible higher cultivation product than a massive cell in less numbers.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENT
The author appreciates all lab technicians and specifically Professor Mark Smales and Dr. Dan Liyod who patiently guided and leaded him to increase his knowledge.
REFERENCES
|
Abad S, Turon X (2012). Valorization of biodiesel derived glycerol as a carbon source to obtain added-value metabolites focus on polyunsaturated fatty acids. Biotechnol. Adv. 30(3):733-741. |
|
|
Becker EW (1994). Microalgae:Biotechnology and Microbiology. Cambridge University Press. 293p. |
|
|
Bergmann M, Peters R H (1980). A simple reflectance method for the measurement of particular pigment in lake water and its application to Phosphorus-Chlorophyll-Seston Relationship. Can. J. Fish. Aquat. Sci. 37:111-114. |
|
|
Bhatnagar A, Chinnasamy S, Singh M, Das K (2011). Renewable biomass production by mixotrophic algae in the presence of various carbon sources and wastewaters. Appl. Energy. 88 (10): 3425-3431. |
|
|
Boyle N, Morgan J (2009). Flux balance analysis of primary metabolism in Chlamydomonas reinhardtii. BMC Syst. Biol. 3:4. |
|
|
Brennan L, Owende P (2010). Biofuels from microalgae - a review of technologies for production, processing, and extractions of biofuels and co-products. Renew. Sustain. Energy Rev. 14:557-577. |
|
|
Bumbak F, Cook S, Zachleder V, Hauser S, Kova K (2011). Best practices in heterotrophic high-cell-density microalgal processes. Appl. Microbiol. Biotechnol. 91(1):31-46. |
|
|
Cerón García M, Sánchez Mirón A, Fernández Sevilla J, Molina Grima E, García Camacho F (2005). Mixotrophic growth of the microalga Phaeodactylum tricornutum. Influence of different nitrogen and organic carbon sources on productivity and biomass composition. Process Biochem. 40(1):297-305. |
|
|
Cerón García M, Sánchez Mirón A, Fernández Sevilla J, Molina Grima E, García Camacho F (2000). Mixotrophic growth of the microalga Phaeodactylum tricornutum on glycerol, growth rate and fatty acid. J. Appl. Phycol. 12:239-248. |
|
|
Chen F, Johns MR (1996). Heterotrophic growth of Chlamydomonas reinhardtii on acetate in chemostat culture. Process Biochem. 31:601-604. |
|
|
Chisti Y (2007). Biodiesel from microalgae. Biotechnol. Adv. 25:294-306. |
|
|
Choi WY, Oh SH, Seo YC, Kim GB, Kang DH, Lee SY, Jung KH, Cho JS, Ahn JH, Choi GP, Lee HY (2011). Effects of methanol on cell growth and lipid production from mixotrophic cultivation of Chlorella sp. Biotechnol. Bioprocess Eng. 16(5):946-955. |
|
|
De-Bashan LE, Bashan Y, Moreno M, Lebsky V K, Bustillos, JJ (2002). Increased pigment and lipid content, lipid variety and cell and population size of the microalgae Chlorella spp. When co-immobilized in alginate beads with the microalgae-growth promoting bacterium Azospirillum brasilense. Can. J. Microbiol. 48:514-521. |
|
|
De-Bashan LE, Hernandez J P, Morey T, Bashan Y (2004). Microalgae growth, promoting bacteria as "helpers" for microalgae, a novel approach for removing ammonium and phosphorus from municipal wastewater. Water Res. 38:466-474. |
|
|
Dragone G, Fernandes B, Vicente A, Teixeira JA (2010). Third generation biofuels from microalgae. In. Vilas, A.M. (Ed.), Current Research, Technology and Education Topics in Applied Microbiology and Microbial Biotechnology. Formatex Research Center, Badajoz. 2:1355-1366. |
|
|
Droop MR(1974). Heterotrophy of carbon. Algal physiology and biochemistry. Edited by Stewart WDP. Oxford. Blackwell. pp. 530-559. |
|
|
Falk S, Samuelsson G, Oquist G (2006). Temperature-dependent photo inhibition and recovery of photosynthesis in the green alga Chlamydomonas reinhardtii acclimated to 12 and 27°C. Wiley Online Library 78(2):173-180. |
|
|
Fischer B, Eggen R, Trebst A, Krieger-Liszkay A (2006). The glutathione peroxidase homologous gene Gpxh in Chlamydomonas reinhardtii is upregulated by singlet oxygen produced in photosystem II. Planta 223:583-590 |
|
|
Gardner R, Cooksey K, Mus F, Macur R, Moll K, Eustance E, Carlson R, Gerlach R, Fields M, Peyton B (2012). Use of sodium bicarbonate to stimulate triacylglycerol accumulation in the Chlorophyte Scenedesmus. sp. and the diatom Phaeodactylum tricornutum. J. Appl. Phycol. 24(5):1311-1320. |
|
|
Gardner R, Lohman E, Gerlach R, Cooksey K, Peyton B M (2013). Comparison of CO2 and Bicarbonate as Inorganic Carbon Sources for Triacylglycerol and Starch Accumulation in Chlamydomonas reinhardtii. Biotechnol. Bioeng. 110:87-96. |
|
|
Gorman DS, Levine RP (1965). Cytochrome f and plastocyanin. their sequence in the photosynthetic electron transport chain of Chlamydomonas reinhardii. U.S.A: Proc. Natl. Acad. Sci. 54:1665-1669. |
|
|
Harun R, Singh M, Forde GM, Danquah MK (2010). Bioprocess engineering of microalgae to produce a variety of consumer products. Renew. Sustain. Energy Rev. 14:1037-1047. |
|
|
Hempel F, Lau J, Klingl A, Maier U G (2011). Algae as Protein Factories. Expression of a Human Antibody and the Respective Antigen in the Diatom Phaeodactylum tricornutum. PLoS ONE 6(12):e28424. |
|
|
Kaplan D, Richmond A, Dubinsky Z, Aaronson S (1986). Algal nutrition. In. Handbook for Microalgal Mass Culture. Edited by Richmond, A. Boca Raton FL, USA. CRC Press. pp. 147-198. |
|
|
Lange L, Nobel P, Osmond C, Ziegler H (1981). Physiological Plant Ecology I Responses to the Physical Environment 12A. Springer-Verlag. 67:259. |
|
|
Lebeau T, Robert JM, Subba Rao DV (2006). Biotechnology of immobilized micro algae: a culture technique for the future. In. Algal Cultures, Analogues of Blooms and Applications. Science Publishers 9(1):801-837. |
|
|
Ledford H, Chin B, Niyogi K (2007). Acclimation to singlet oxygen stress in Chlamydomonas reinhardtii. Eukaryot. Cell. 6:919-930. |
|
|
Liang Y, Sarkany N, Cui Y (2009). Biomass and lipid productivities of Chlorella vulgaris under autotrophic, heterotrophic and mixotrophic growth conditions. Biotechnol. Lett. 31 (7): 1043–1049. |
|
|
Ludwig W, Strunk O, Klugbauer S, Klugbauer N, Weizenegger M, Neumaier J, Bachleitner M, Schleifer KH (1998). Bacterial phylogeny base on comparative sequence analysis. Electrophoresis 19(4):554-68. |
|
|
Mandalam RK, Palsson BØ (1998). Elemental balancing of biomass and medium composition enhances growth capacity in high-density Chlorella vulgaris cultures. Biotechnol. Bioeng. 59:605-611. |
|
|
Milledge J (2010). Commercial application of microalgae other than as biofuels. A brief review. Rev. Environ. Sci. Biotechnol. 10(1):31-41. |
|
|
Moon M, Kim C, Park W, Yoo G, Choi Y, Yang J (2013). Mixotrophic growth with acetate or volatile fatty acids maximizes growth and lipid production in Chlamydomonas reinhardtii. Algal Res. 2:352-357. |
|
|
Neilson A, Lewin R (1974). The uptake and utilization of organic carbon by algae. an essay in comparative biochemistry. Phycologia 13:227-264. |
|
|
Octavio PG, Froylan MEE, Luz E de-B, Yoav B (2010). A review of Heterotrophic cultures of microalgae: Metabolism and potential products. Water Res. 45:11-36. |
|
|
Peers G, Truong T, Ostendorf E, Busch A, Elrad D, Grossman A, Hippler M, Niyogi K (2009). An ancient light-harvesting protein is critical for the regulation of algal photosynthesis. Nature 462:518-521. |
|
|
Potvin G, Zhang Z (2010). Strategies for high-level recombinant protein expression in transgenic microalgae. Biotechnol. Adv. 28:910- 918. |
|
|
Pulz O (2001). Photo bioreactors: production systems for phototrophic microorganisms. Appl. Microbiol. Biotechnol. 57:287-293. |
|
|
Pulz O, Gross W (2004). Valuable products from biotechnology microalgae. Appl. Microbiol. Biotechnol. 65:635-648. |
|
|
Ras M, Steyer J, Bernard O (2013). Temperature effect on micro algae. A crucial factor for outdoor production. Rev. Environ. Sci. Biotechnol. 12(2):153-164. |
|
|
Roacha T, Sedoud A, Krieger-Liszkay A (2013). Acetate in mixotrophic growth medium affects photosystem II in Chlamydomonas reinhardtii and protects against photoinhibition. Biochim. Biophys. Acta 1827(10):1141-1256. |
|
|
Scholz M, Hoshino T, Johnson D, Riley M R, Cuello J (2011). Flocculation of wall-deficient cells of Chlamydomonas reinhardtii mutant cw15 by calcium and methanol. Biomass Bioenergy 35(12):4835-4840. |
|
|
Shi X M, Liu H J, Zhang X W, Chen F (1999). Production of biomass and lutein by Chlorella protothecoides at various glucose concentrations in heterotrophic cultures. Process Biochem. 34(4):341-347. |
|
|
Spreitzer RJ, Al-Abed S, Huether M (1998). Temperature-sensitive photosynthesis deficient mutants of Chlamydomonas reinhardtii. Plant Physiol. 86(3):773-777. |
|
|
Stephens E, Ross IL, King Z, Mussgnug JH, Kruse O, Posten C, Borowitzka MA, Hankamer B (2010). An economic and technical evaluation of microalgal biofuels. Nat. Biotechnol. 28:126-128. |
|
|
Suh IS, Lee CG (2003). Photobioreactor engineering: design and performance. Biotechnol. Bioprocess Eng. 8:313-321. |
|
|
White D, Pagarette A, Rooks P, Ali S (2012). The effect of sodium bicarbonate supplementation on growth and biochemical composition of marine microalgae cultures. J. Appl. Phycol. 25(1):153-165. |
|
|
Wijffels RH, Barbosa MJ (2010). An outlook on microalgal biofuels. Science 329:796-799. |
|
|
Yang C, Hua Q, Shimizu K (2000). Energetics and carbon metabolism during growth of microalgal cells under photoautotrophic, mixotrophic and cyclic light-autotrophic/dark-heterotrophic conditions. Biochem. Eng. J. 6: 87-102. |
|
|
Yu E Zendejas F, Lane P, Gaucher S, Simmons B, Lane T (2009). Triacylglycerol accumulation and profiling in the model diatoms Thalassiosira pseudonana and Phaeodactylum tricornutum (Baccilariophyceae) during starvation. J. Appl. Phycol. 21:669-681. |
|
|
Zhang X W, Chen F, Johns M R (1999). Kinetic models for heterotrophic growth of Chlamydomonas reinhardtii in batch and fed-batch cultures. Process Biochem. 35:385-389. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


