Full Length Research Paper

ABSTRACT
Approximately 80-90% of cultivated grassland areas in Brazil are covered by Urochloa brizantha [syn. Brachiaria brizantha (Hochst. ex A. Rich.) Stapf.]. Some genotypes of Urochloa have being widely used with a wrong nomenclature, like species and cultivars. In this way, the Urochloa cultivar identification is primordial for breeding programs and seed production. Considering the importance of genetic purity in commercialized seed lots, the present work aimed to evaluate the use of inter-simple sequence repeat (ISSR) markers in six cultivars of U. brizantha (Xaraés; Piatã; Basilisk; MG4; MG5 and Marandu) to discriminate and determine the contamination in seed batches. Results showed that it is possible to discriminate all cultivar with only two primers in pure samples. Basilisk was confirmed as a U. brizantha cultivar. ISSR markers showed a low polymorphism level. It was not possible to separate samples intentionally contaminated even at 5%.
Key words: Brachiaria, molecular markers, varietal purity.
INTRODUCTION
MATERIALS AND METHODS

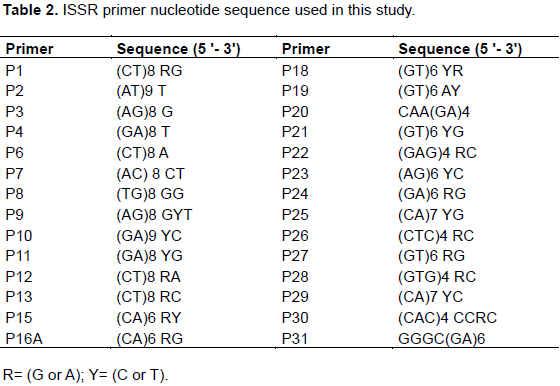
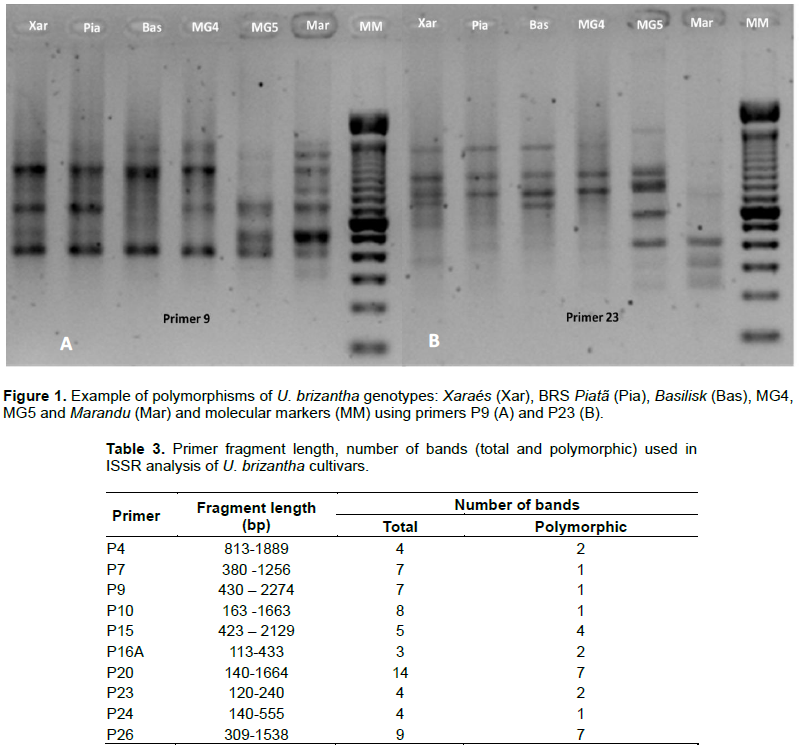
RESULTS AND DISCUSSION
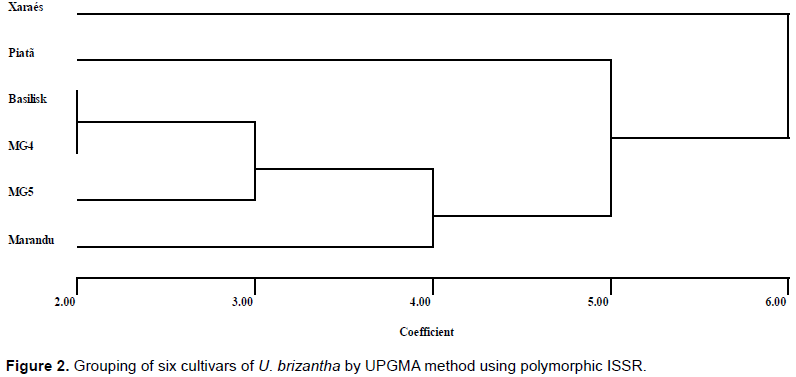

CONCLUSIONS
CONFLICT OF INTERESTS
REFERENCES
|
Almeida CMA, Lima SN, Lima GSA, Brito JZ, Donato VMTS, Silva IMV (2009). Caracterização molecular de cultivares de cana-de-açúcar utilizando marcadores ISSR. Ciênc. Agrotec. 33:1771-1776. |
|
|
Almeida MCC, Chiari L, Jank L, Valle CB (2011). Diversidade genética molecular entre cultivares e híbridos de Brachiaria spp. e Panicum maximum. Ciênc. Rural 41:1998-2003. |
|
|
Ambiel AC, Machado Neto NB, Guaberto LM, Vanderlei TM (2008). Agrupamento de acessos e cultivares de três espécies de Brachiaria por RAPD. Acta Scient. Agron. 30:457-464. |
|
|
Ambiel AC, Machado Neto NB, Guaberto LM, Vanderlei TM (2010). Brachiaria germplasm dissimilarity shown by the RAPD markers. Crop Breed. Appl. Biotechnol. 10:55-64. |
|
|
Anand KK, Srivastava RK, Chaudhary LB, Singh AK (2010). Delimitation of species of the Astragalus rhizanthus complex (Fabaceae) using molecular markers RAPD, ISSR and DAMD. Taiwania 55:197-207. |
|
|
Assis GML, Euclydes RF, Cruz CM, Valle CB (2003). Discrimination species of Urochloa based on different groups of morphological characters. Rev. Bras. Zootec. 32:576-584. |
|
|
Azevedo ALS, Costa PP, Machado MA, De Paula CMP, Sobrinho FS (2011). High degree of genetic diversity among genotypes of the forage grass Brachiaria ruziziensis (Poaceae) detected with ISSR markers. Genet. Mol. Res. 10:3530-3538. |
|
|
Bianchi VJ, Fachinello JC, Schuch MW, Sansavini S (2004). Molecular characterization of cultivars of peach and nectarine with microsatellites. Rev. Bras. Frutic. 26:490-493. |
|
|
Brasil (2008). Ministério da Agricultura e Reforma Agrária. Instrução Normativa nº 30, de 21 de maio de 2008. |
|
|
Brasil (2009). Ministério da Agricultura e Reforma Agrária. Regras para análise de sementes. Brasília, DF, CLAV, 2009. 365p. |
|
|
Brasil (2011). Ministério da Agricultura e Reforma Agrária. Instrução |
|
|
Normativa nº 30, de 9 de Junho de 2011. |
|
|
Brasil (2016). Ministério da Agricultura e Pecuária. Registro Nacional de Cultivares. Available at: |
|
|
Brody JR, Kern SE (2004). Sodium boric acid: a Tris-free, cooler conductive medium for DNA electrophoresis. Biotechniques 36(2):214-217. |
|
|
Crusciol CAC, Soratto RP, Borghi E, Mateus GP (2009). Integração lavoura-pecuária: benefícios das gramíneas perenes nos sistemas de produção. Inf. Agron. 125:1-15. |
|
|
De Paula LA, Bianchi VJ, Fachinello JC (2012). Molecular characterization and genetic variability among port peach grafts based on codominant markers. Pesq. Agropecu. Bras. 47:193-199. |
|
|
Dogan B, Duran A, Hakki EE (2007). Phylogenetic analysis of Jurinea (Asteraceae) species from Turkey based on ISSR amplification. Ann. Bot. Fenn. 44:353-358. |
|
|
Ferreira AME, Grattapaglia D (2012). Introduction to using RAPD and RFLP markers in genetic analysis. 3.ed. Brasilia: EMBRAPA-CENARGEN, Brasil. P.220 |
|
|
Gupta M, Chyi YS, Romero-Severson J, Owen JL (1994). Amplification of DNA markers from evolutionarily diverse genomes using single primers of simple-sequence repeats. Theor. Appl. Genet. 89:998-1006. |
|
|
Haq QMR, Singh U, Arif M, Zaidi N (2011). Genetic variability within Fusarium solani as revealed by PCR-fingerprinting based on ISSR markers. Indian Phytopathol. 61:305-310. |
|
|
Jorgensen T, Hauser TP, Jorgensen RB (2007). Adventitious presence of other varieties in oilseed rape (Brassica napus) from seed banks and certified seed. Seed Sci. Res. 17:115-126. |
|
|
Karia CT, Duarte JB, Araújo ACG (2006). Cultivar development of the genus Brachiaria (Trin.) Griseb in Brazil. Planaltina: Embrapa Cerrado. 57. (Embrapa Cerrado Documents, 163). |
|
|
Keller-Grein G, Maass BL, Hanson T (1996) Natural variation in Brachiaria and existing germplasm collections. In: Miles JW, Maass BL and Valle CB (Eds.). Brachiaria: biology, agronomy, and improvement. 45 Cali, CIAT. pp. 16-42. |
|
|
Lin XC, Lou YF, Liu J, Peng JS, Liao GL, Fang W (2010). Crossbreeding of Phyllostachys species (Poaceae) and identification of their hybrids Liao using ISSR markers. Genet. Mol. Res. 9:1398-1404. |
|
|
Macedo MCM (2006). Respostas morfofisiológicas de Brachiaria spp. ao alagamento do solo e a síndrome da morte do capim-marandu. Embrapa Amazônia Oriental. |
|
|
Marotti I, Bonetti A, Minelli M, Catizone P, Dinelli G (2007). Characterization of some italian common bean (Phaseolus vulgaris L.) landraces by RAPD, semi-random and molecular markers ISSR. Genet. Resour. Crop Evol. 54:175-188. |
|
|
Matsuda (2016). Espécies forrageiras. Available at |
|
|
Pharmawati M, Yan G, Mcfarlane IJ (2004). Application of RAPD and ISSR markers to analyze molecular relationships in Grevillea (Proteaceae). Aust. Syst Bot. 17:49-61. |
|
|
Ramos NP, Brunelli KR, Camargo LDA, Marcos Filho J (2006). Sensibilidade dos Microssatélites para determinar a pureza varietal em sementes de milho. Rev. Bras. Semen. 28:99-105. |
|
|
Reddy MP, Sarla N, Siddiq EA (2002). Inter simple sequence repeat (ISSR) polymorphism and its application in plant breeding. Euphytica 128:9-17. |
|
|
Renvoize SA, Clayton WD, Kabuye CHS (1996). Morphology, taxonomy, and natural distribution of Brachiaria (Trin.) Griseb. In. Miles JW et al. (Ed.). Brachiaria: biology, agronomy, and improvement. Cali: CIAT, 1996. pp. 1-17. |
|
|
Rodrigues JF, van den Berg C, Abreu AG, Novello M, Veasey EA, Oliveira GC, Koehler S. (2015). Species delimitation of Cattleya coccinea and C. mantiqueirae (Orchidaceae): insights from phylogenetic and population genetics analyses. Plant Syst. Evol. 301(5):1345-1359. |
|
|
Rohlf FJ (2004). NTSYS-Pc: numerical taxonomy and multivariate analysis system, version 2.1, user's guide New York: Exeter Software. |
|
|
Saini N, Jain N, Jain S, Jain RK (2004). Assessment of genetic diversity within and among Basmati and Basmati rice varieties non-using AFLP, ISSR and SSR markers. Euphytica 140:133-146. |
|
|
Satya P, Karan M, Sarkar D, Sinha MK (2012). Genome synteny and evolution of AABB allotetraploids in Hibiscus section Furcaria revealed by interspecific hybridization, ISSR and SSR markers. Plant Syst. Evol. 298:1257-1270. |
|
|
Silva KVP, Alves AAC, Martins MIS, Melo CAF, Carvalho R (2011). Variabilidade genética entre acessos do gênero Manihot por meio de marcadores moleculares ISSR. Pesq. Agropecu. Bras. 46:1082-1088. |
|
|
Valle CB, Simioni C (2008). Melhoramento genético da Braquiária. In. Resende, RMS; Valle CB; Jank, L. Melhoramento de forrageiras tropicais. Campo Grande: Embrapa Gado de Corte. |
|
|
Vigna BBZ, Jugmann L, Francisco PM, Zucchi MI, Valle CB, Souza AP (2011). Genetic diversity and population structure of the Brachiaria brizantha germplasm. Trop. Plant Biol. 4:157-169. |
|
|
Vilela H (2005). Pastagem: seleção de plantas forrageiras, implantação e adubação. Aprender Fácil. Viçosa. |
|
|
Wickert E, Lemos EDM, Pereira FM, Mayer M (2012). Marcadores fAFLP na caracterização de três genótipos de umezeiro selecionados como porta-enxertos para pessegueiro. Pesq. Agropecu. Bras. 42:1741-1746. |
|
|
Wolfe AD, Liston A (1998). Contributions of PCR-based methods to plant systematics and evolutionary biology. In. Molecular systematics of plants II. Springer US. pp. 43-86. |
|
|
Wood TE, Nakazato T (2009). Investigating species boundaries in the Giliopsis group of Ipomopsis (Polemoniaceae): Strong discordance among molecular and morphological markers. Am. J. Bot. 96:853-861. |
|
|
Zanine AM, Dias PF, Souta M, Ferreira DJ, Santos EM, Pinto FB (2007). Uso de funções discriminantes para comparação de cultivares dos gêneros Cynodon e Digitaria quanto à produção de matéria seca e teores de macronutrientes. Arch. Latinoam. Prod. Anim. 15(4):159-164. |
|
|
Zietkiewicz E, Rafalski A, Labuda D (1994). Genome fingerprinting by simple sequence repeat (SSR) anchored polymerase chain-reaction amplification. Genomics 20:176-183. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


