Full Length Research Paper

ABSTRACT
This study assessed the capabilities of the dung beetle, Euoniticellus intermedius (Coleoptera: Scarabaeida), larva gut consortia in degrading cellulose that can serve as glucose source for biofuels production. A total of 144 live dung beetles were randomly collected from a dairy farm and bred in a temperature controlled insect rearing room. On reaching the late second to third instar stage, dung beetle larvae were harvested, dissected and the gut micro-flora were cultured in medium containing cellulose as sole carbon source. Microbial growth (total protein concentration) and cellulose degradation activity (reducing sugars concentration) in the cellulose cultures were monitored successively for 15 days. Statistical analysis showed that there was significant microbial growth, but no significant increase in reducing sugar levels. Despite the lack of increase in reducing sugar levels, it was concluded that the dung beetle larva gut has micro-flora with cellulose degrading capabilities that allowed it to grow and survive in the cellulose minimal medium.
Key words: Dung beetle, Euoniticellus intermedius, cellulose, biofuels.
INTRODUCTION
The global energy demand has increased more than twenty fold in the last century (Jegannathan et al., 2009). Previously, abundant and cheaply accessible, fossil fuels have undoubtedly played a big role in the industrialization of most of the developed states seen today and they are still an integral part of today’s energy mix. However, due to their non-renewable nature, fossil fuel reserves have been dwindling at an alarming rate, at a crucial time when the world’ energy needs have also been on a constant rise (Adsul et al., 2011; Jegannathan et al., 2009). In addition, the damaging environmental effects such as global warming and climate change, associated with fossil fuel use have also compounded the current challenges attributed to the world’s heavy reliance on fossil fuels (Adsul et al., 2011; Jegannathan et al., 2009). To counter the consequences associated with rampant fossil fuel use, massive investments in renewable and sustainable energy resources have been recently noted with sustainable bio-fuels research also given top priority.
Historically, transportation bio-fuels like bio-ethanol have primarily been derived from food sources such as sugar cane and corn (Balat and Balat, 2009; Yuan et al., 2008), a situation that has sparked a lot of controversy on food versus fuel debate. This has consequently led to more research being focused towards the production of second generation bio-fuels, such as cellulose-derived bio-ethanol. Despite huge financial investment and concerted research efforts in this area, and the abundance of cellulose raw materials, production of second generation bio-ethanol at an industrial scale is still hampered by unreasonably high operating costs due to the recalcitrant nature of cellulose, which ultimately makes its production economically unfeasible (Margeot et al., 2009; Ruane et al., 2010).
Many organisms (cattle, rabbits, buffalo, elephants and insects) have been reported to have the capability to degrade cellulose matter through the use of symbiotic micro-organisms and their own cellulose degrading enzymes (Inoue et al., 2005; Feng et al., 2007; Duan et al., 2009; Wang et al., 2009). Scarab beetles have been reported as one of such organisms since they can effectively utilize various cellulose plant matter and animal waste as energy sources (Huang et al., 2010). Dung beetles belong to the family Scarabaeidae and order Coleoptera, which is regarded as the largest order in the insect kingdom. Scarab beetles are commonly found in various environmental niches where decaying plant and/or animal wastes form a high proportion of the available biomass (Huang et al., 2010). This is because scarabaeids are reportedly herbivorous or saprophagous and many of their species feed on plant roots, decaying organic matter and animal waste of very low nutritional value (Zhang and Jackson, 2008). As such, scarab beetles play an important role in the ecological control of dung, decaying wood and plant matter (Koyama et al., 2003).
Even today, a lot of research is still being undertaken in a bid to find the elusive solution and consequently break the financial barriers which are currently hindering any progress in the industrial production of the second generation bio-fuels. In this study, the biology of E. intermedius, a dung beetle that spends its life cycle in cow dung was studied. The dung beetle larvae were selected in this study because they eat the actual dung, whereas adult dung beetles only survived on the dung juices (Khanyile et al., 2008). Consequently, it is expected that the larval stage of this organism possesses biological capability for efficient processing of dung. This research seeks to contribute to the growing list of possible biological routes of cellulose degradation by investigating the cellulose degrading capabilities of the symbiotic micro-flora naturally residing in the gut of the dung beetle, Euoniticellus intermedius (Coleoptera: Scarabaeida), larvae, and their potential application and contribution to the sustainable production of cellulose-derived bio-fuels.
MATERIALS AND METHODS
Dung beetle collection
In this study, a total of 144 dung beetles that were randomly collected from a dairy farm about 10 km South-West of Johannesburg, South Africa were used. The insects were collected between August and September, 2010 and bred at a temperature (28°C) controlled dung beetle rearing room. According to Edwards (1991), this is regarded as the best time to collect the dung beetles in Johannesburg, due to the high rainfall patterns, which favour high reproduction and survival rates. The adult dung beetles were collected from deep inside or the surface of the soil beneath the dung pats.
Dung beetle rearing
Initially, 144 beetles were sorted into 17 males and 127 females. Males were identified by the presence of a horn on the head, a feature that is absent in the females. In population numbers, the females were found to be more than 6 times numerous than the males. It is also known that a single male can mate with more than one female. Therefore, instead of using 3 pairs of dung beetles in equal male and female proportion, two males were mixed with four females in a breeding container. Breeding was done in plastic containers measuring 160 × 130 × 130 mm. The containers were half-filled with thoroughly sieved and compacted slightly moist soil. Fresh cattle dung collected from the same farm as the dung beetles was used as the dung beetle feed and was stored in small 1 kg plastic packets at -20°C. On every third day (72 h), one packet of dung was added to each container containing the breeding pairs. The soil component provided the beetles with tunnel burrowing space to make the brood balls.
Once a week, the breeding containers were sieved to remove any brood balls present. An average of 20 to 30 brood balls per week was harvested from each plastic container with dung beetle breeding pairs. The brood balls from different containers were then pooled together and placed in a separate plastic container, half-filled with moist sand, measuring 400 × 300 × 200 mm and then labeled according to the date of collection. A wet sponge was placed on top of the sand to maintain the desired moisture levels.
Larvae developed inside the brood ball and their nourishment came from the dung contained therein. Unlike the adult dung beetles which feed on the fluid component of the dung, the larvae feed by ingesting whole dung particles (Edwards, 1991). To propagate the dung beetle culture, larvae were initially allowed to grow for 4 to 5 weeks developing into adults. Newly, emerged beetles were collected and kept in single-sex plastic containers for 2 weeks to allow them to develop into sexual maturity (Lailvaux et al., 2005; Pomfret and Knell, 2006).
Dung beetle larvae dissection
Larvae were allowed to develop for approximately 2 to 3 weeks before harvesting them for dissection. This allowed them to grow into late second or third instars.
A modified dung beetle dissection protocol similar to the one described by Lemke et al. (2003) was used to perform the dissection. The steel/metal dissection equipment was autoclaved at 121°C for 15 min. The preparation dish was immersed in 70% ethanol and left overnight (approximately 12 hours) to minimize microbial contamination. Insect ringer solution was prepared according to the protocol described by Hayashi and Kamimura (2001) and autoclaved for 15 min at 121°C. Larvae dissection was performed in a sterile preparation dish under a dissecting microscope, in a laminar flow cabinet. The larvae were first anesthetized by exposing them to a nitrogen, hydrogen and carbon dioxide (71/7/22 vol/vol respectively) (Afrox grade) gas mixture for 15 min. They were then fixed onto the preparation dish with steel pins, with the larvae laid on its sides, and sterile insect ringer solution added. The cuticle was cut along the side lines and the ventral integument, circular muscles and trachea were then carefully removed. After decapitating a larva, a circular cut was made on the anus to allow careful retrieval of the intestinal tract. The guts were then stored in 1.5 ml tubes at -70°C.
Culturing of dung beetle larvae gut micro-flora
Media preparation
A basal cellulose medium for gut micro-flora cultivation was prepared using cellulose (Whatman filter paper) as the sole carbon source, according to a modified method described by Lemke et al. (2003). To the basal media preparation, 1 ml of trace element solution (SL11) and 1 ml of selenium tungstate solution were added and then adjusted to pH 8 using a 0.1 M sodium phosphate buffer before autoclaving.
Media inoculation
Larval guts weighing 1 g were homogenized in 9 ml of phosphate buffered saline (pH 7.4). The homogenate (1 ml) was then inoculated into 9 ml of cellulose media (in the place of Whatman filter paper, 0.405 g of cellulose microcrystalline was used during pre-culturing experiments). The inoculations were done in triplicate. The pre-cultures were incubated aerobically at 30°C for 48 h in the dark, shaking at a speed of 80 revolutions/min on a rotary shaker. Microbial growth was visually observed by the degree of turbidity of cultures after 24 and 48 h of incubation period.
Inoculation of pre-culture samples (10 ml) into the main cultures (90 ml) was done in triplicate after 48 h of pre-culturing. The main cultures were incubated aerobically at 30°C in the dark, shaking at 80 rpm for 15 days. Samples for microbial growth determination and cellulose degradation activity were aseptically taken on a daily basis, 3 ml per culture sample and kept at -70°C until analysis.
Microbial growth determination
In order to monitor microbial growth, the time (number of days) required to achieve maximal microbial growth was determined as a function of total protein concentration, using the Bradford protein assay procedure (Bradford, 1976). This assay is based on the observation that the absorbance maximum for an acidic solution of Coomassie Brilliant Blue G-250 shifts from 465 to 595 nm when binding to protein occurs. Both hydrophobic and ionic interactions play a crucial role in stabilizing the anionic form of the dye, resulting in a visible colour change (Bradford, 1976; Stoscheck, 1990).
Samples (Day 0 to Day 15) for microbial growth determination were taken daily in triplicate (one sample from each main culture) and pooled together to make a composite sample. Due to the poor microbial growth in the culture samples, the Bradford micro-assay method was used to plot the standard curve.
Total protein concentration determination in the test-samples was done by adding test-samples to the Bradford working reagent in a glass spectrophotometer cuvette and incubating the mixture for 5 min at room temperature. Volume adjustments were done on the samples to get absorbance values of between 0.1 and 0.9. The absorbance values of the test-samples were then measured using a spectrophotometer at 595 nm wavelength, as mentioned earlier. The total protein concentrations of the samples were determined by comparing the samples’ absorbance values against the plotted Bradford micro-assay standard curve. Any volume adjustments performed on the samples were catered for during calculations of total protein concentration. Measurements were done in quartet for every composite daily culture sample. Total protein concentration values were regarded as directly proportional to microbial growth.
Cellulose degradation
Cellulose degradation activity was determined by measuring the concentration of released reducing sugars in the culture samples using a modified dinitrosalicyclic acid (DNS) assay described by Adney and Baker (1996).
To measure the reducing sugar concentration in the cellulose culture samples, 0.5 ml samples were added to test tubes containing 1 ml citrate buffer and 3 ml of DNS solution added, as described earlier. The mixture was boiled for 5 to 10 min, allowed to cool. Absorbance of the test-samples was then measured at 540 nm without diluting them. Reducing sugar concentration was determined by comparing the absorbance values of test-samples against the plotted glucose standard curve. The volume adjustments made for the standard curve were taken into consideration during the test-sample glucose/reducing sugar concentration calculations. Measurements were done in quartet for every composite culture sample.
RESULTS
Microbial growth
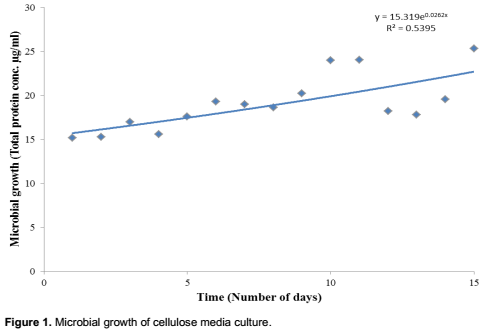
Microbial growth was observed during the course of 15 days increasing from a total protein concentration of 15.1955 µg/ml on Day 1 to 25.34475 µg/ml on Day 15. This increase was shown to be significant at 5% level using the statistical analysis equation y = 15.319e0.0262x and R2 value of 0.5395 as shown in Figure 1. Statistical analyses were done using Microsoft Excel (2010).
Cellulose degradation
The reducing sugar concentrations in culture samples remained relatively constant at 2.9 mg/ml from Day 0 to Day 15, with an exception of Day 5 where a concentration of 3.1 mg/ml was observed (Figure 2).

DISCUSSION
The observed increase in microbial growth is attributed to the ability of the dung beetle larva gut consortia to successfully degrade cellulose and use the by-products for its metabolic activities essential for growth and survival. This is in agreement with previous research that has shown that different insects have gut microbial consortia capable of digesting cellulose material into many different metabolites (Lemke et al., 2003; Dillion and Dillion, 2004; Egert et al., 2005). Instead of sharp increase in growth normally associated with microbial cultures, the observed steady growth pattern can be explained by the disruptive effect of cellulose medium culturing (by only selecting cellulose degrading micro-flora and excluding non-cellulose degrading organisms) to the inherent community structure and relationships which naturally exists in the dung beetle larva gut. According to Streit and Schmitz (2004), many culture-dependent approaches currently used to explore the diversity and potential of microbial communities are biased, because of the limitations of cultivation methods. In addition to this, in the dung beetle larva digestive tract, different types of micro-organisms work synergistically, by performing different roles which collectively contribute to the survival and well-being of the whole microbial community (Streit and Schmitz, 2004).
Previous research work has shown that the gut of scarab beetle larvae comprises a rich population of xylan and pectin degrading micro-organisms (Egert et al., 2005; Zhang and Jackson, 2008). The lack of increase in reducing sugar concentration in the cultures is attributed to the ability of the larva gut micro-flora to immediately utilize the produced reducing sugars as a carbon source, in addition to cellulose matter. As mentioned previously, in the digestive tract of the dung beetle larva, different types of micro-organisms work synergistically by performing different roles (these may include cellulose degradation and other hydrolytic roles), which collectively contribute to the survival and well-being of the whole microbial community (Huang et al., 2010). This can mean that any reducing sugars produced during the cellulose degradation process were immediately utilized by another sub-section of the cultured gut micro-flora before they could be accounted for using the DNS assay.
CONCLUSION
The cellulose degradation capability of the cultured dung beetle larva gut micro-flora was successfully demon-strated by the ability of the gut micro-flora to grow in exclusive cellulose medium. Despite the observed insignificant increase in reducing sugars, it can be concluded that the mere ability of the gut micro-flora to significantly grow and survive on cellulose media containing only filter paper as the sole carbon source should confirm that there is indeed a sub-set of gut micro-flora from E. intermedius that can degrade cellulose matter.
ACKNOWLEDGEMENTS
The authors would like to thank Mr. Lazarus Chapungu for his help in editing this manuscript. Special thanks go to Mr. Phanankosi Moyo for laying the foundation upon which this study was established and for taking time from his busy schedule to review the manuscript. This research was funded by the University of the Witwatersrand Postgraduate Merit Award (PMA), the Bradlow Scholarship and the African Union Commission (Mwalimu Nyerere African Union Postgraduate Scholarship).
REFERENCES
|
Adney B, Baker J (1996). Measurement of cellulase activities. Laboratory Analytical Procedures (LAP). Technical Report NREL/TP-510-42628. January 2008. |
|
|
Adsul MG, Singhvi MS, Gaikaiwari SA, Gokhale DV (2011). Development of biocatalysts for production of commodity chemicals from lignocellulosic biomass. Bioresour. Technol. 102:43042-4312. |
|
|
Balat M, Balat H (2009). Recent trends in global production and utilization of bio-ethanol fuel. Appl. Energy. 86:2273-2282. |
|
|
Bradford MM (1976). A rapid and sensitive method for the quantitation of microgram quantitites of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72:248-254. |
|
|
Dillion RJ, Dillion VM (2004). The gut bacteria of insects: Non-pathogenic interactions. Annu. Rev. Entomol. 49:71-92. |
|
|
Duan C, Xian L, Zhao G, Feng Y, Pang H, Bai X, Tang J, Ma Q, Feng J (2009). Isolation and partial characterization of novel genes encoding acidic cellulases from metagenomes of buffalo rumens. J. Appl. Microbiol. 107:245-256. |
|
|
Edwards PB (1991). Seasonal variation in the dung of African grazing mammals, and its consequences for coprophagous insects. Funct. Ecol. 5:617-628. |
|
|
Egert M, Stingl U, Bruun LD, Pommerenke B, Brune A, Friedrich MW (2005). Structure and topology of microbial communities in the major gut compartments of Melolontha melolontha larvae (Coleoptera: Scarabaeidae). Appl. Environ. Microbiol. 71:4556-4566. |
|
|
Feng Y, Duan C, Pang H, Mo X, Wu C, Yu Y, Hu Y, Wei J, Tang J, Feng J (2007). Cloning and identification of novel cellulase genes from uncultured microorganisms in rabbit cecum and characterization of the expressed cellulases. Appl. Microbiol. Biotechnol. 75:319-328. |
|
|
Hayashi F, Kamimura Y (2001). The potential for incorporation of male derived proteins into developing eggs in the leafhopper Bothrogonia ferruginea. J. Insect Physiol. 48:153-159. |
|
|
Huang S, Zhang H, Marshall S, Jackson TA (2010). The scarab gut: A potential bioreactor for bio-fuel production. Insect Sci. 17:175-183. |
|
|
Inoue T, Moriya S, Ohkuma M, Kudo T (2005). Molecular cloning and characterization of a cellulase gene from a symbiotic protist of the lower termite, Coptotermes formosanus. Gene 349:67-75. |
|
|
Jegannathan KR, Chan E, Ravindra P (2009). Harnessing biofuels: A global Renaissance in energy production? Renew. Sustain. Energy Rev. 13: 2163-2168. |
|
|
Khanyile L M, Hull R, Ntwasa M (2008). Dung beetle database: Comparison with other invertebrate transcriptomes. Bioinformation 3 (4):159-161. |
|
|
Koyama M, Iwata R, Yamane A, Katase T, Ueda S (2003). Nutrient intake in the third instar larvae of Anomala cuprea and Protaetia orientalis submarmorea (Coleoptera: Scarabaeidae) from a mixture of cow dung and wood chips: Results from stable isotope analyses of nitrogen and carbon. Appl. Entomol. Zool. 38:305-311. |
|
|
Lailvaux SP, Hathway J, Pomfret J, Knell RJ (2005). Horn size predicts physical performance in the beetle Euoniticellus intermedius (Coleoptera: Scarabaeidae). Funct. Ecol. 19:632-639. |
|
|
Lemke T, Stingl U, Egert M, Friedrich MW, Brune A (2003). Physicochemical conditions and microbial activities in the highly alkaline gut of the humus feeding larva of Pachnoda ephippiata (Coleoptera: Scarabaeidae). Appl. Environ. Microbiol. 69:6650-6658 |
|
|
Margeot A, Hahn-Hagerdal B, Edlund M, Slade R, Monot F (2009). New improvements for lignocellulosic ethanol. Curr. Opin. Biotechnol. 20:372-380. |
|
|
Pomfret JC, Knell RJ (2006). Sexual selection and horn allometry in the dung beetle Euoniticellus intermedius. Anim. Behav. 71:567-576. |
|
|
Ruane J, Sonnino A, Agostini A (2010). Bioenergy and the potential contribution of agricultural biotechnologies in developing countries. Biomass Bioenergy 34:1427-1439. |
|
|
Stoscheck CM (1990). Quantitation of Protein. Methods Enzymol. 182:50-69. |
|
|
Streit WR, Schmitz RA (2004). Metagenomics – the key to the uncultured microbes. Curr. Opin. Microbiol. 7:492-498. |
|
|
Wang F, Li F, Chen G, Liu W (2009). Isolation and characterization of novel cellulase genes from uncultured microorganisms in different environmental niches. Microbiol. Res. 164:650-657. |
|
|
Yuan JS, Tiller KH, Al-Ahmad H, Stewart NR, Stewart Jr. CN (2008). Plants to power: bioenergy to fuel the future. Trends Plant Sci. 13:421-429. |
|
|
Zhang HY, Jackson TA (2008). Autochthonous bacterial flora indicated by PCR-DGGE of 16S rRNA gene fragments from the alimentary tract of Costelytra zealandica (Coleoptera: Scarabaeidae). J. Appl. Microbiol. 105:1277-1285. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


